
The Quest for Neurodegenerative Disease Treatment—Focusing on Alzheimer’s Disease Personalised Diets


Abstract
:1. Introduction
2. Alzheimer’s Disease and Food—What Is the Correlation between Pathophysiology and Nutrition
3. Vitamin E Odyssey and AD

{kind=link}
{kind=link}
| Study | Study Type | Study Details | Intervention | Results |
|---|---|---|---|---|
| Animal models | ||||
| [52] | Review | - | - |
|
| [56] | RCS | Rat hippocampal cell culture | 50 µM α-tocopherol |
|
| [57] | RCS | Mouse astrocytes | 100 µM Trolox |
|
| [53] | RCS | Rats | 333 IU/kg α-tocopherol |
|
| Human patients | ||||
| [59] | Meta-analysis | 135,967 participants | ≥400 IU/day vs. <400 IU/day Vit. E |
|
| [60] | RCT | 613 participants | 2000 IU/day α-tocopherol |
|
| [60] | RCT | 847 participants | 2000 IU/day α-tocopherol |
|
| [62] | RCT | 6377 women | 600 IU q.a.d. α-tocopherol |
|
| [61] | Review | - | - |
|
4. Why Will Omega-3 Fatty Acids Not Be Enough?
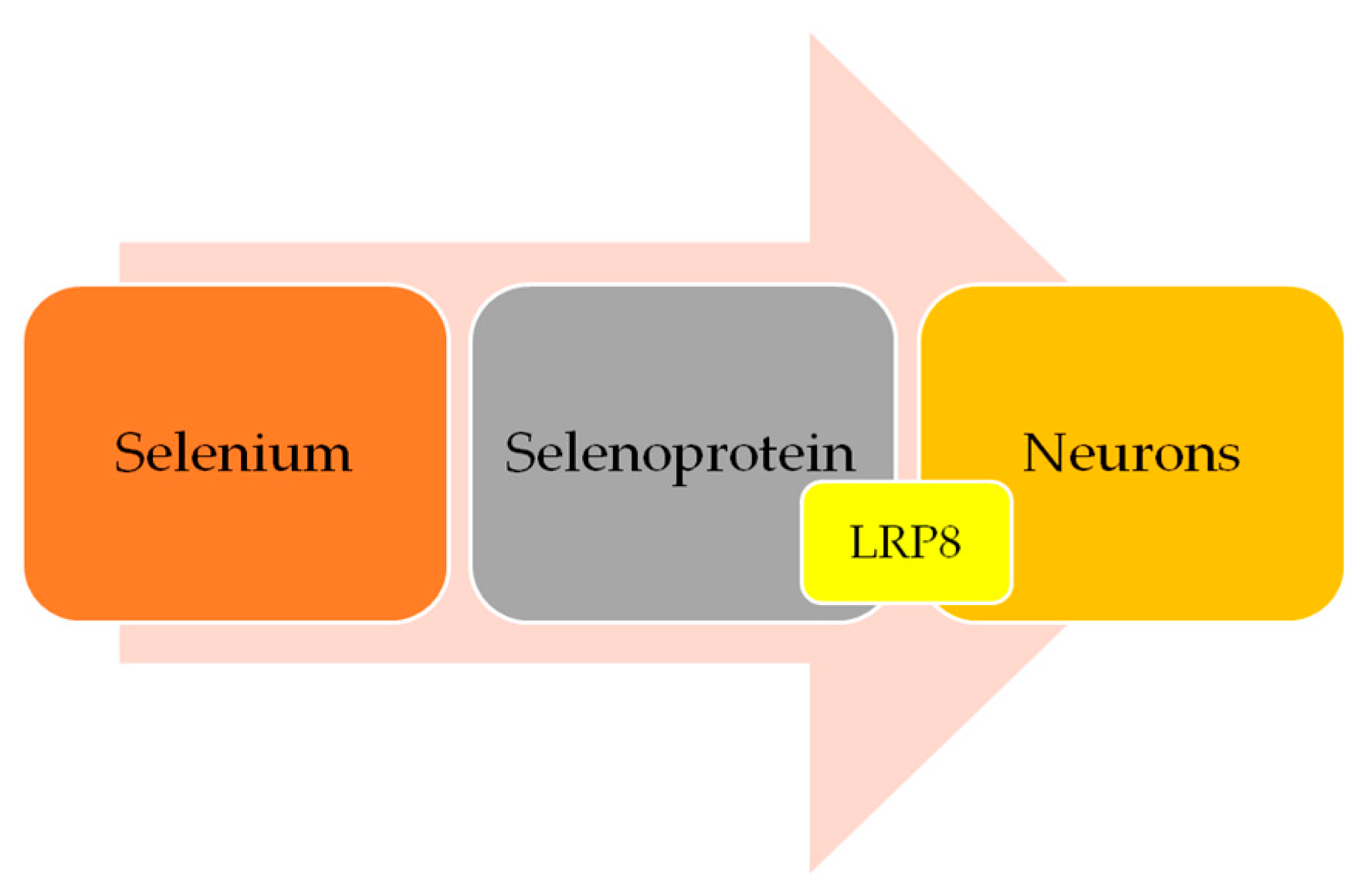
5. Selenium and AD: Is There a Link?
6. Traditional Medicine—How Would It Help?
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Checkoway, H.; Lundin, J.I.; Kelada, S.N. Neurodegenerative diseases. IARC Sci. Publ. 2011, 163, 407–419. [Google Scholar]
- Goodman, R.A.; Lochner, K.A.; Thambisetty, M.; Wingo, T.S.; Posner, S.F.; Ling, S.M. Prevalence of dementia subtypes in United States Medicare fee-for-service beneficiaries, 2011–2013. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2017, 13, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Society, UK. Alzheimer’s Society’s View on Demography. Available online: https://www.alzheimers.org.uk/about-us/policy-and-influencing/what-we-think/demography (accessed on 30 June 2020).
- GBD 2019 Dementia Forecasting Collaborators. Estimation of the global prevalence of dementia in 2019 and forecasted prevalence in 2050: An analysis for the Global Burden of Disease Study 2019. Lancet. Public Health 2022, 7, e105–e125. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.L.; Tong, G.; Ballard, C. Treatment Combinations for Alzheimer’s Disease: Current and Future Pharmacotherapy Options. J Alzheimers Dis. 2019, 67, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Lyketsos, C.G. Treatment Development for Alzheimer’s Disease: How Are We Doing? Adv. Exp. Med. Biol. 2020, 1195, 19. [Google Scholar] [CrossRef]
- Briggs, R.; Kennelly, S.P.; O’Neill, D. Drug treatments in Alzheimer’s disease. Clin Med. 2016, 16, 247–253. [Google Scholar] [CrossRef]
- Duce, J.A.; Zhu, X.; Jacobson, L.H.; Beart, P.M. Therapeutics for dementia and Alzheimer’s disease: New directions for precision medicine. Br. J. Pharmacol. 2019, 176, 3409–3412. [Google Scholar] [CrossRef]
- Mossello, E.; Ballini, E. Management of patients with Alzheimer’s disease: Pharmacological treatment and quality of life. Ther. Adv. Chronic Dis. 2012, 3, 183–193. [Google Scholar] [CrossRef]
- Ströhle, A.; Schmidt, D.K.; Schultz, F.; Fricke, N.; Staden, T.; Hellweg, R.; Priller, J.; Rapp, M.A.; Rieckmann, N. Drug and Exercise Treatment of Alzheimer Disease and Mild Cognitive Impairment: A Systematic Review and Meta-Analysis of Effects on Cognition in Randomized Controlled Trials. Am. J. Geriatr. Psychiatry Off. J. Am. Assoc. Geriatr. Psychiatry 2015, 23, 1234–1249. [Google Scholar] [CrossRef]
- Xu, H.; Garcia-Ptacek, S.; Jönsson, L.; Wimo, A.; Nordström, P.; Eriksdotter, M. Long-term Effects of Cholinesterase Inhibitors on Cognitive Decline and Mortality. Neurology 2021, 96, e2220–e2230. [Google Scholar] [CrossRef]
- Murakawa-Hirachi, T.; Mizoguchi, Y.; Ohgidani, M.; Haraguchi, Y.; Monji, A. Effect of memantine, an anti-Alzheimer’s drug, on rodent microglial cells in vitro. Sci. Rep. 2021, 11, 6151. [Google Scholar] [CrossRef]
- Padda, I.S.; Parmar, M. Aducanumab. Available online: https://www.ncbi.nlm.nih.gov/books/NBK573062/ (accessed on 31 August 2022).
- Ebell, M.H. Aducanumab Is Not Effective for Mild Cognitive Impairment or Mild Alzheimer Dementia and Has Major Safety Issues. Am. Fam. Physician 2022, 106, 462–463. [Google Scholar]
- Panza, F.; Lozupone, M.; Solfrizzi, V.; Custodero, C.; Valiani, V.; D’Introno, A.; Stella, E.; Stallone, R.; Piccininni, M.; Bellomo, A.; et al. Chapter 9-Contribution of Mediterranean Diet in the Prevention of Alzheimer’s Disease; Academic Press: Cambridge, MA, USA, 2018; pp. 139–155. ISBN 9780128119594. [Google Scholar] [CrossRef]
- Sofi, F.; Abbate, R.; Gensini, G.F.; Casini, A. Accruing evidence on benefits of adherence to the Mediterranean diet on health: An updated systematic review and meta-analysis. Am. J. Clin. Nutr. 2010, 92, 1189–1196. [Google Scholar] [CrossRef]
- García-Casares, N.; Gallego Fuentes, P.; Barbancho, M.Á.; López-Gigosos, R.; García-Rodríguez, A.; Gutiérrez-Bedmar, M. Alzheimer’s Disease, Mild Cognitive Impairment and Mediterranean Diet. A Systematic Review and Dose-Response Meta-Analysis. J. Clin. Med. 2021, 10, 4642. [Google Scholar] [CrossRef]
- McCleery, J.; Abraham, R.P.; Denton, D.A.; Rutjes, A.W.; Chong, L.Y.; Al-Assaf, A.S.; Griffith, D.J.; Rafeeq, S.; Yaman, H.; Malik, M.A.; et al. Vitamin and mineral supplementation for preventing dementia or delaying cognitive decline in people with mild cognitive impairment. Cochrane Database Syst. Rev. 2018, 11, CD011905. [Google Scholar] [CrossRef]
- Śliwińska, S.; Jeziorek, M. The role of nutrition in Alzheimer’s disease. Rocz. Panstw. Zakl. Hig. 2021, 72, 29–39. [Google Scholar] [CrossRef]
- Nishal, S.; Phaugat, P.; Bazaad, J.; Dhaka, R.; Khatkar, S.; Khatkar, A.; Khayatkashani, M.; Alizadeh, P.; Haghighi, S.M.; Mehri, M.; et al. A concise review of common plant-derived compounds as potential therapy for Alzheimer’s disease and Parkinson’s disease: Insight into Structure-Activity-Relationship. CNS Neurol. Disord. Drug Targets. 2023, 22. [Google Scholar] [CrossRef]
- Guo, P.; Zhang, B.; Zhao, J.; Wang, C.; Wang, Z.; Liu, A.; Du, G. Medicine-Food Herbs against Alzheimer’s Disease: A Review of Their Traditional Functional Features, Substance Basis, Clinical Practices and Mechanisms of Action. Molecules 2022, 27, 901. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Gibson, M.G.; Sharma, V. Diagnostic and Statistical Manual of Mental Disorders: DSM-5; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 0890425558. [Google Scholar]
- Khan, S.; Barve, K.H.; Kumar, M.S. Recent Advancements in Pathogenesis, Diagnostics and Treatment of Alzheimer’s Disease. Curr. Neuropharmacol. 2020, 18, 1106–1125. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological alterations in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D. Imaging the progression of Alzheimer pathology through the brain. Proc. Natl. Acad. Sci. USA 2002, 99, 4135–4137. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.B.; Janelidze, S.; Ossenkoppele, R.; Kvartsberg, H.; Brinkmalm, A.; Mattsson-Carlgren, N.; Stomrud, E.; Smith, R.; Zetterberg, H.; Blennow, K.; et al. Untangling the association of amyloid-β and tau with synaptic and axonal loss in Alzheimer’s disease. Brain 2021, 144, 310–324. [Google Scholar] [CrossRef] [PubMed]
- Golde, T.E. Alzheimer’s disease–the journey of a healthy brain into organ failure. Mol. Neurodegener. 2022, 17, 18. [Google Scholar] [CrossRef]
- Kisler, K.; Nelson, A.R.; Montagne, A.; Zlokovic, B.V. Cerebral blood flow regulation and neurovascular dysfunction in Alzheimer disease. Nat. Rev. Neurosci. 2017, 18, 419–434. [Google Scholar] [CrossRef]
- Hussain, B.; Fang, C.; Chang, J. Blood-Brain Barrier Breakdown: An Emerging Biomarker of Cognitive Impairment in Normal Aging and Dementia. Front. Neurosci. 2021, 15, 688090. [Google Scholar] [CrossRef]
- Sharma, C.; Woo, H.; Kim, S.R. Addressing Blood–Brain Barrier Impairment in Alzheimer’s Disease. Biomedicines 2022, 10, 742. [Google Scholar] [CrossRef]
- Gąsiorowski, K.; Brokos, B.; Echeverria, V.; Barreto, G.E.; Leszek, J. RAGE-TLR Crosstalk Sustains Chronic Inflammation in Neurodegeneration. Mol. Neurobiol. 2018, 55, 1463–1476. [Google Scholar] [CrossRef]
- Wang, W.Y.; Tan, M.S.; Yu, J.T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann. Transl. Med. 2015, 3, 136. [Google Scholar] [CrossRef]
- DiSabato, D.J.; Quan, N.; Godbout, J.P. Neuroinflammation: The devil is in the details. J. Neurochem. 2016, 139 (Suppl. 2), 136–153. [Google Scholar] [CrossRef] [Green Version]
- Picca, A.; Calvani, R.; Coelho-Junior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial Dysfunction, Oxidative Stress, and Neuroinflammation: Intertwined Roads to Neurodegeneration. Antioxidants 2020, 9, 647. [Google Scholar] [CrossRef]
- Patergnani, S.; Bouhamida, E.; Leo, S.; Pinton, P.; Rimessi, A. Mitochondrial Oxidative Stress and "Mito-Inflammation": Actors in the Diseases. Biomedicines 2021, 9, 216. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, X. Antioxidant therapies for Alzheimer’s disease. Oxid. Med. Cell. Longev. 2012, 2012, 472932. [Google Scholar] [CrossRef]
- Jurcau, A. The Role of Natural Antioxidants in the Prevention of Dementia-Where Do We Stand and Future Perspectives. Nutrients 2021, 13, 282. [Google Scholar] [CrossRef]
- Norton, S.; Matthews, F.E.; Barnes, D.E.; Yaffe, K.; Brayne, C. Potential for primary prevention of Alzheimer’s disease: An analysis of population-based data. Lancet. Neurol. 2014, 13, 788–794. [Google Scholar] [CrossRef]
- Farooqui, A.A.; Ong, W.Y.; Horrocks, L.A.; Chen, P.; Farooqui, T. Comparison of biochemical effects of statins and fish oil in brain: The battle of the titans. Brain Res. Rev. 2007, 56, 443–471. [Google Scholar] [CrossRef]
- Lukiw, W.J.; Bazan, N.G. Docosahexaenoic acid and the aging brain. J. Nutr. 2008, 138, 2510–2514. [Google Scholar] [CrossRef]
- Jacobs, D.R., Jr.; Orlich, M.J. Diet pattern and longevity: Do simple rules suffice? A commentary. Am. J. Clin. Nutr. 2014, 100 (Suppl. 1), 313S–319S. [Google Scholar] [CrossRef]
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Doré, J.; Franceschi, C.; Lehtinen, M.J.; Recker, T.; Salvioli, S.; et al. Health relevance of the modification of low grade inflammation in ageing (inflammageing) and the role of nutrition. Ageing Res Rev. 2017, 40, 95–119. [Google Scholar] [CrossRef]
- Miquel, S.; Champ, C.; Day, J.; Aarts, E.; Bahr, B.A.; Bakker, M.; Bánáti, D.; Calabrese, V.; Cederholm, T.; Cryan, J.; et al. Poor cognitive ageing: Vulnerabilities, mechanisms and the impact of nutritional interventions. Ageing Res. Rev. 2018, 42, 40–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monacelli, F.; Acquarone, E.; Giannotti, C.; Borghi, R.; Nencioni, A. Vitamin C, Aging and Alzheimer’s Disease. Nutrients 2017, 9, 670. [Google Scholar] [CrossRef] [PubMed]
- Milanski, M.; Degasperi, G.; Coope, A.; Morari, J.; Denis, R.; Cintra, D.E.; Tsukumo, D.M.; Anhe, G.; Amaral, M.E.; Takahashi, H.K.; et al. Saturated fatty acids produce an inflammatory response predominantly through the activation of TLR4 signaling in hypothalamus: Implications for the pathogenesis of obesity. J. Neurosci. 2009, 29, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Solch, R.J.; Aigbogun, J.O.; Voyiadjis, A.G.; Talkington, G.M.; Darensbourg, R.M.; O’Connell, S.; Pickett, K.M.; Perez, S.R.; Maraganore, D.M. Mediterranean diet adherence, gut microbiota, and Alzheimer’s or Parkinson’s disease risk: A systematic review. J. Neurol. Sci. 2022, 434, 120166. [Google Scholar] [CrossRef]
- Hang, Z.; Lei, T.; Zeng, Z.; Cai, S.; Bi, W.; Du, H. Composition of intestinal flora affects the risk relationship between Alzheimer’s disease/Parkinson’s disease and cancer. Biomed. Pharmacother. 2022, 145, 112343. [Google Scholar] [CrossRef]
- Chen, L.; Xu, X.; Wu, X.; Cao, H.; Li, X.; Hou, Z.; Wang, B.; Liu, J.; Ji, X.; Zhang, P.; et al. A comparison of the composition and functions of the oral and gut microbiotas in Alzheimer’s patients. Front. Cell Infect. Microbiol. 2022, 12, 942460. [Google Scholar] [CrossRef]
- McGrattan, A.M.; McGuinness, B.; McKinley, M.C.; Kee, F.; Passmore, P.; Woodside, J.V.; McEvoy, C.T. Diet and Inflammation in Cognitive Ageing and Alzheimer’s Disease. Curr. Nutr. Rep. 2019, 8, 53–65. [Google Scholar] [CrossRef]
- Galli, F.; Azzi, A.; Birringer, M.; Cook-Mills, J.M.; Eggersdorfer, M.; Frank, J.; Cruciani, G.; Lorkowski, S.; Özer, N.K. Vitamin E: Emerging aspects and new directions. Free Radic. Biol. Med. 2017, 102, 16–36. [Google Scholar] [CrossRef]
- Lloret, A.; Esteve, D.; Monllor, P.; Cervera-Ferri, A.; Lloret, A. The Effectiveness of Vitamin E Treatment in Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 20, 879. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Bramanti, P.; Mazzon, E. Role of Vitamin E in the Treatment of Alzheimer’s Disease: Evidence from Animal Models. Int. J. Mol. Sci. 2017, 18, 2504. [Google Scholar] [CrossRef]
- Zhao, R.; Han, X.; Zhang, H.; Liu, J.; Zhang, M.; Zhao, W.; Jiang, S.; Li, R.; Cai, H.; You, H. Association of vitamin E intake in diet and supplements with risk of dementia: A meta-analysis. Front. Aging Neurosci. 2022, 14, 955878. [Google Scholar] [CrossRef]
- Yatin, S.M.; Varadarajan, S.; Butterfield, D.A. Vitamin E Prevents Alzheimer’s Amyloid beta-Peptide (1-42)-Induced Neuronal Protein Oxidation and Reactive Oxygen Species Production. J. Alzheimers Dis. 2000, 2, 123–131. [Google Scholar] [CrossRef]
- Scimemi, A.; Meabon, J.S.; Woltjer, R.L.; Sullivan, J.M.; Diamond, J.S.; Cook, D.G. Amyloid-β1-42 slows clearance of synaptically released glutamate by mislocalizing astrocytic GLT-1. J. Neurosci. 2013, 33, 5312–5318. [Google Scholar] [CrossRef]
- Yamada, K.; Tanaka, T.; Han, D.; Senzaki, K.; Kameyama, T.; Nabeshima, T. Protective effects of idebenone and alpha-tocopherol on beta-amyloid-(1-42)-induced learning and memory deficits in rats: Implication of oxidative stress in beta-amyloid-induced neurotoxicity in vivo. Eur. J. Neurosci. 1999, 11, 83–90. [Google Scholar] [CrossRef]
- Miller, E.R., 3rd; Pastor-Barriuso, R.; Dalal, D.; Riemersma, R.A.; Appel, L.J.; Guallar, E. Meta-analysis: High-dosage vitamin E supplementation may increase all-cause mortality. Ann. Intern. Med. 2005, 142, 37–46. [Google Scholar] [CrossRef]
- Dysken, M.W.; Sano, M.; Asthana, S.; Vertrees, J.E.; Pallaki, M.; Llorente, M.; Love, S.; Schellenberg, G.D.; McCarten, J.R.; Malphurs, J.; et al. Effect of vitamin E and memantine on functional decline in Alzheimer disease: The TEAM-AD VA cooperative randomized trial. JAMA-J. Am. Med. Assoc. 2014, 311, 33–44. [Google Scholar] [CrossRef]
- Hermann, D.M. Insufficient evidence for vitamin E in Alzheimer’s disease. Alzheimers Dement. 2016, 2, 199–201. [Google Scholar] [CrossRef]
- Kang, J.H.; Cook, N.; Manson, J.; Buring, J.E.; Grodstein, F. A randomized trial of vitamin E supplementation and cognitive function in women. Arch Intern. Med. 2006, 166, 2462–2468. [Google Scholar] [CrossRef]
- O’Brien, J.T.; Holmes, C.; Jones, M.; Jones, R.; Livingston, G.; McKeith, I.; Mittler, P.; Passmore, P.; Ritchie, C.; Robinson, L.; et al. Clinical practice with anti-dementia drugs: A revised (third) consensus statement from the British Association for Psychopharmacology. J. Psychopharmacol. 2017, 31, 147–168. [Google Scholar] [CrossRef]
- Lloret, A.; Badía, M.C.; Mora, N.J.; Pallardó, F.V.; Alonso, M.D.; Viña, J. Vitamin E paradox in Alzheimer’s disease: It does not prevent loss of cognition and may even be detrimental. J. Alzheimers Dis. 2009, 17, 143–149. [Google Scholar] [CrossRef]
- Petersen, R.C.; Thomas, R.G.; Grundman, M.; Bennett, D.; Doody, R.; Ferris, S.; Galasko, D.; Jin, S.; Kaye, J.; Levey, A.; et al. Alzheimer’s Disease Cooperative Study Group. Vitamin E and donepezil for the treatment of mild cognitive impairment. N. Engl. J. Med. 2005, 352, 2379–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, J. New approaches to symptomatic treatments for Alzheimer’s disease. Mol Neurodegener. 2021, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Dubois, B.; Hampel, H.; Feldman, H.H.; Scheltens, P.; Aisen, P.; Andrieu, S.; Bakardjian, H.; Benali, H.; Bertram, L.; Blennow, K.; et al. Proceedings of the Meeting of the International Working Group (IWG) and the American Alzheimer’s Association on “The Preclinical State of AD”; July 23, 2015; Washington DC, USA. Preclinical Alzheimer’s disease: Definition, natural history, and diagnostic criteria. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2016, 12, 292–323. [Google Scholar] [CrossRef]
- Pavlik, V.N.; Doody, R.S.; Rountree, S.D.; Darby, E.J. Vitamin E use is associated with improved survival in an Alzheimer’s disease cohort. Dement. Geriatr. Cogn. Disord. 2009, 28, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Dyall, S.C. Long-chain omega-3 fatty acids and the brain: A review of the independent and shared effects of EPA, DPA and DHA. Front. Aging Neurosci. 2015, 7, 52. [Google Scholar] [CrossRef]
- Avallone, R.; Vitale, G.; Bertolotti, M. Omega-3 Fatty Acids and Neurodegenerative Diseases: New Evidence in Clinical Trials. Int. J. Mol. Sci. 2019, 20, 4256. [Google Scholar] [CrossRef]
- Rosendorff, C.; Beeri, M.S.; Silverman, J.M. Cardiovascular risk factors for Alzheimer’s disease. Am. J. Geriatr. Cardiol. 2007, 16, 143–149. [Google Scholar] [CrossRef]
- Lee, J.H.; O’Keefe, J.H.; Lavie, C.J.; Marchioli, R.; Harris, W.S. Omega-3 fatty acids for cardioprotection. Mayo Clin. Proc. 2008, 83, 324–332. [Google Scholar] [CrossRef]
- Maclean, C.H.; Issa, A.M.; Newberry, S.J.; Mojica, W.A.; Morton, S.C.; Garland, R.H.; Hilton, L.G.; Traina, S.B.; Shekelle, P.G. Effects of omega-3 fatty acids on cognitive function with aging, dementia, and neurological diseases. Evid. Rep. Technol. Assess 2005, 114, 1–3. [Google Scholar]
- Wassall, S.R.; Brzustowicz, M.R.; Shaikh, S.R.; Cherezov, V.; Caffrey, M.; Stillwell, W. Order from disorder, corralling cholesterol with chaotic lipids. The role of polyunsaturated lipids in membrane raft formation. Chem. Phys. Lipids 2004, 132, 79–88. [Google Scholar] [CrossRef]
- Gamoh, S.; Hashimoto, M.; Sugioka, K.; Shahdat Hossain, M.; Hata, N.; Misawa, Y.; Masumura, S. Chronic administration of docosahexaenoic acid improves reference memory-related learning ability in young rats. Neuroscience 1999, 93, 237–241. [Google Scholar] [CrossRef]
- Conklin, S.M.; Gianaros, P.J.; Brown, S.M.; Yao, J.K.; Hariri, A.R.; Manuck, S.B.; Muldoon, M.F. Long-chain omega-3 fatty acid intake is associated positively with corticolimbic gray matter volume in healthy adults. Neurosci. Lett. 2007, 421, 209–212. [Google Scholar] [CrossRef]
- Conquer, J.A.; Tierney, M.C.; Zecevic, J.; Bettger, W.J.; Fisher, R.H. Fatty acid analysis of blood plasma of patients with Alzheimer’s disease, other types of dementia, and cognitive impairment. Lipids 2000, 35, 1305–1312. [Google Scholar] [CrossRef]
- Barberger-Gateau, P.; Raffaitin, C.; Letenneur, L.; Berr, C.; Tzourio, C.; Dartigues, J.F.; Alpérovitch, A. Dietary patterns and risk of dementia: The Three-City cohort study. Neurology 2007, 69, 1921–1930. [Google Scholar] [CrossRef]
- Tully, A.M.; Roche, H.M.; Doyle, R.; Fallon, C.; Bruce, I.; Lawlor, B.; Coakley, D.; Gibney, M.J. Low serum cholesteryl ester-docosahexaenoic acid levels in Alzheimer’s disease: A case-control study. Br. J. Nutr. 2003, 89, 483–489. [Google Scholar] [CrossRef]
- Burk, R.F.; Hill, K.E. Selenoprotein P-expression, functions, and roles in mammals. Biochim Biophys. Acta. 2009, 1790, 1441–1447. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Sies, H. Selenium homeostasis and antioxidant selenoproteins in brain: Implications for disorders in the central nervous system. Arch. Biochem. Biophys. 2013, 536, 152–157. [Google Scholar] [CrossRef]
- Ray, R.; Juranek, J.K.; Rai, V. RAGE axis in neuroinflammation, neurodegeneration and its emerging role in the pathogenesis of amyotrophic lateral sclerosis. Neurosci. Biobehav. Rev. 2016, 62, 48–55. [Google Scholar] [CrossRef]
- Shahar, A.; Patel, K.V.; Semba, R.D.; Bandinelli, S.; Shahar, D.R.; Ferrucci, L.; Guralnik, J.M. Plasma selenium is positively related to performance in neurological tasks assessing coordination and motor speed. Mov. Disord. 2010, 25, 1909–1915. [Google Scholar] [CrossRef]
- González-Domínguez, R.; García-Barrera, T.; Gómez-Ariza, J.L. Homeostasis of metals in the progression of Alzheimer’s disease. Biometals 2014, 27, 539–549. [Google Scholar] [CrossRef]
- Alehagen, U.; Aaseth, J.; Alexander, J.; Johansson, P.; Larsson, A. Supplemental selenium and coenzyme Q10 reduce glycation along with cardiovascular mortality in an elderly population with low selenium status-A four-year, prospective, randomised, double-blind placebo-controlled trial. J. Trace. Elem. Med. Biol. 2020, 61, 126541. [Google Scholar] [CrossRef] [PubMed]
- Aaseth, J.; Skalny, A.V.; Roos, P.M.; Alexander, J.; Aschner, M.; Tinkov, A.A. Copper, Iron, Selenium and Lipo-Glycemic Dysmetabolism in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 9461. [Google Scholar] [CrossRef] [PubMed]
- Tamtaji, O.R.; Heidari-Soureshjani, R.; Mirhosseini, N.; Kouchaki, E.; Bahmani, F.; Aghadavod, E.; Asemi, Z. Probiotic and selenium co-supplementation, and the effects on clinical, metabolic and genetic status in Alzheimer’s disease: A randomized, double-blind, controlled trial. Clin. Nutr. 2019, 38, 2569–2575. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhang, W.; Liu, W.; Zhu, W.; Guo, R.; Wang, Y.; Zhang, D.; Wang, J. The inhibitory effect of selenium nanoparticles on protein glycation in vitro. Nanotechnology 2015, 26, 145703. [Google Scholar] [CrossRef]
- Li, Y.B.; Han, J.Y.; Jiang, W.; Wang, J. Selenium inhibits high glucose-induced cyclooxygenase-2 and P-selectin expression in vascular endothelial cells. Mol. Biol. Rep. 2011, 38, 2301–2306. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Song, G.L. Roles of Selenoproteins in Brain Function and the Potential Mechanism of Selenium in Alzheimer’s Disease. Front. Neurosci. 2021, 15, 646518. [Google Scholar] [CrossRef]
- Leiter, O.; Zhuo, Z.; Rust, R.; Wasielewska, J.M.; Grönnert, L.; Kowal, S.; Walker, T.L. Selenium mediates exercise-induced adult neurogenesis and reverses learning deficits induced by hippocampal injury and aging. Cell Metab. 2022, 34, 408–423. [Google Scholar] [CrossRef]
- Cardoso, B.R.; Roberts, B.R.; Malpas, C.B.; Vivash, L.; Genc, S.; Saling, M.M.; Bush, A.I. Supranutritional sodium selenate supplementation delivers selenium to the central nervous system: Results from a randomized controlled pilot trial in Alzheimer’s disease. Neurotherapeutics 2019, 16, 192–202. [Google Scholar] [CrossRef]
- Singh, S.K.; Srivastav, S.; Castellani, R.J.; Plascencia-Villa, G.; Perry, G. Neuroprotective and Antioxidant Effect of Ginkgo biloba Extract Against AD and Other Neurological Disorders. Neurotherapeutics 2019, 16, 666–674. [Google Scholar] [CrossRef]
- Kandiah, N.; Ong, P.A.; Yuda, T.; Ng, L.L.; Mamun, K.; Merchant, R.A.; Chen, C.; Dominguez, J.; Marasigan, S.; Ampil, E.; et al. Treatment of dementia and mild cognitive impairment with or without cerebrovascular disease: Expert consensus on the use of Ginkgo biloba extract, EGb 761®. CNS Neurosci. Ther. 2019, 25, 288–298. [Google Scholar] [CrossRef]
- Chen, L.; Xu, X.; Cao, H.; Li, H. Comparison of effects of different dietary interventions on cognitive function in Alzheimer’s disease: Protocol for systematic review and network meta-analysis. BMJ Open. 2021, 11, e042997. [Google Scholar] [CrossRef]
- Yusufov, M.; Weyandt, L.L.; Piryatinsky, I. Alzheimer’s disease and diet: A systematic review. Int. J. Neurosci. 2017, 127, 161–175. [Google Scholar] [CrossRef]
- Gregory, J.; Vengalasetti, Y.V.; Bredesen, D.E.; Rao, R.V. Neuroprotective Herbs for the Management of Alzheimer’s Disease. Biomolecules 2021, 11, 543. [Google Scholar] [CrossRef]
- Alzobaidi, N.; Quasimi, H.; Emad, N.A.; Alhalmi, A.; Naqvi, M. Bioactive Compounds and Traditional Herbal Medicine: Promising Approaches for the Treatment of Dementia. Degener. Neurol. Neuromuscul. Dis. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Tsai, Y.-T.; Kao, S.-T.; Cheng, C.-Y. Medicinal Herbs and Their Derived Ingredients Protect against Cognitive Decline in In Vivo Models of Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 11311. [Google Scholar] [CrossRef]
- Balmus, I.M.; Ciobica, A. Main Plant Extracts’ Active Properties Effective on Scopolamine-Induced Memory Loss. Am. J. Alzheimers Dis. Other Demen. 2017, 32, 418–428. [Google Scholar] [CrossRef]
- Olivera-Pueyo, J.; Pelegrín-Valero, C. Dietary supplements for cognitive impairment. Actas. Esp. Psiquiatr. 2017, 45 (Suppl.), 37–47. [Google Scholar]
- Liu, H.; Ye, M.; Guo, H. An Updated Review of Randomized Clinical Trials Testing the Improvement of Cognitive Function of Ginkgo biloba Extract in Healthy People and Alzheimer’s Patients. Front. Pharmacol. 2020, 10, 1688. [Google Scholar] [CrossRef]
- Bonassi, S.; Prinzi, G.; Lamonaca, P.; Russo, P.; Paximadas, I.; Rasoni, G.; Rossi, R.; Ruggi, M.; Malandrino, S.; Sánchez-Flores, M. Clinical and genomic safety of treatment with Ginkgo biloba L. leaf extract (IDN 5933/Ginkgoselect®Plus) in elderly: A randomised placebo-controlled clinical trial [GiBiEx]. BMC Complement. Altern. Med. 2018, 18, 22. [Google Scholar] [CrossRef]
- Gauthier, S.; Schlaefke, S. Efficacy and tolerability of Ginkgo biloba extract EGb 761® in dementia: A systematic review and meta-analysis of randomized placebo-controlled trials. Clin. Interv. Aging. 2014, 9, 2065–2077. [Google Scholar] [CrossRef]
- Meng, M.; Ai, D.; Sun, L.; Xu, X.; Cao, X. EGb 761 inhibits Aβ1-42-induced neuroinflammatory response by suppressing P38 MAPK signaling pathway in BV-2 microglial cells. Neuroreport 2019, 30, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.; Rojas-Castaneda, J.; Rojas, P. Antioxidant properties of a Ginkgo biloba leaf extract (EGb 761) in animal models of Alzheimer’s and Parkinson’s diseases. Curr. Top. Nutraceutical. Res 2016, 14, 1. [Google Scholar]
- Cefali, L.C.; Ataide, J.A.; Fernandes, A.R.; Sanchez-Lopez, E.; Sousa, I.M.O.; Figueiredo, M.C.; Ruiz, A.L.T.G.; Foglio, M.A.; Mazzola, P.G.; Souto, E.B. Evaluation of In Vitro Solar Protection Factor (SPF), Antioxidant Activity, and Cell Viability of Mixed Vegetable Extracts from Dirmophandra mollis Benth, Ginkgo biloba L., Ruta graveolens L., and Vitis vinífera L. Plants 2019, 8, 453. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Ni, Y.; Hou, J.; Chen, C.; Zhao, B.; Xin, W. Hydrogen peroxide-induced oxidative damage and apoptosis in cerebellar granule cells: Protection by Ginkgo biloba extract. Pharmacol. Res. 2000, 41, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Wei, T.; Chen, C.; Ni, Y.; Zhao, B.; Hou, J. Mechanisms of apoptosis in rat cerebellar granule cells induced by hydroxyl radicals and the effects of EGb761 and its constituents. Toxicology 2000, 148, 103–110. [Google Scholar] [CrossRef]
- Shi, C.; Liu, J.; Wu, F.; Yew, D.T. Ginkgo biloba extract in Alzheimer’s disease: From action mechanisms to medical practice. Int. J. Mol. Sci. 2010, 11, 107–123. [Google Scholar] [CrossRef]
- Bodalski, T.; Karłowicz-Bodalska, K. Ginkgo biloba L.—Chemistry and activity. Postępy Fitoterapii (Adv. Phytother.) 2006, 4, 195–205. [Google Scholar]
- Abdel-Kader, R.; Hauptmann, S.; Keil, U.; Scherping, I.; Leuner, K.; Eckert, A.; Müller, W.E. Stabilization of mitochondrial function by Ginkgo biloba extract (EGb 761). Pharmacol. Res. 2007, 56, 493–502. [Google Scholar] [CrossRef]
- Wan, W.; Zhang, C.; Danielsen, M.; Li, Q.; Chen, W.; Chan, Y.; Li, Y. EGb761 improves cognitive function and regulates inflammatory responses in the APP/PS1 mouse. Exp. Gerontol. 2016, 81, 92–100. [Google Scholar] [CrossRef]
- Tchantchou, F.; Xu, Y.; Wu, Y.; Christen, Y.; Luo, Y. EGb 761 enhances adult hippocampal neurogenesis and phosphorylation of CREB in transgenic mouse model of Alzheimer’s disease. FASEB J. 2007, 21, 2400–2408. [Google Scholar] [CrossRef]
- Nowak, A.; Kojder, K.; Zielonka-Brzezicka, J.; Wróbel, J.; Bosiacki, M.; Fabiańska, M.; Wróbel, M.; Sołek-Pastuszka, J.; Klimowicz, A. The Use of Ginkgo Biloba L. as a Neuroprotective Agent in the Alzheimer’s Disease. Front. Pharmacol. 2021, 4, 775034. [Google Scholar] [CrossRef]
- Gantait, A.; Barman, T.; Mukherjee, P.K. Validated method for estimation of curcumin in turmeric powder. Ind. J. Trad. Knowledge. 2011, 10, 247–250. [Google Scholar]
- Zhang, L.; Fiala, M.; Cashman, J.; Sayre, J.; Espinosa, A.; Mahanian, M.; Rosenthal, M. Curcuminoids enhance amyloid -beta uptake by macrophages of Alzheimer’s disease patients. J. Alzheimers Dis. 2006, 10, 1–7. [Google Scholar] [CrossRef]
- Mishra, S.; Palanivelu, K. The effect of curcumin (turmeric) on Alzheimer’s disease: An overview. Ann. Indian Acad. Neurol. 2008, 11, 13–19. [Google Scholar] [CrossRef]
- Ambegaokar, S.S.; Wu, L.; Alamshahi, K.; Lau, J.; Jazayeri, L.; Chan, S.; Khanna, P.; Hsieh, E. Curcumin inhibits dose-dependently and time-dependently neuroglial proliferation and growth. Neuro Endocrinol. Lett. 2003, 24, 469–473. [Google Scholar]
- da Costa, I.M.; Freire, M.A.M.; de Paiva Cavalcanti, J.R.L.; de Araújo, D.P.; Norrara, B.; Moreira Rosa, I.M.M.; de Azevedo, E.P.; do Rego, A.C.M.; Filho, I.A.; Guzen, F.P. Supplementation with Curcuma longa Reverses Neurotoxic and Behavioral Damage in Models of Alzheimer’s Disease: A Systematic Review. Curr. Neuropharmacol. 2019, 17, 406–421. [Google Scholar] [CrossRef]
- Kim, H.J.; Jung, S.W.; Kim, S.Y.; Cho, I.H.; Kim, H.C.; Rhim, H.; Kim, M.; Nah, S.Y. Panax ginseng as an adjuvant treatment for Alzheimer’s disease. J Ginseng Res. 2018, 42, 401–411. [Google Scholar] [CrossRef]
- Kim, J.; Kim, S.H.; Lee, D.S.; Lee, D.J.; Kim, S.H.; Chung, S.; Yang, H.O. Effects of fermented ginseng on memory impairment and β-amyloid reduction in Alzheimer’s disease experimental models. J. Ginseng Res. 2013, 37, 100–107. [Google Scholar] [CrossRef]
- Lee, M.R.; Yun, B.S.; In, O.H.; Sung, C.K. Comparative study of korean white, red, and black ginseng extract on cholinesterase inhibitory activity and cholinergic function. J. Ginseng Res. 2011, 35, 421–428. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Yan, X.; Qin, K.; Shi, M.; Lin, T.; Zhu, Y.; Kang, T.; Zhao, G. Protective effects of ginsenoside Rd against okadaic acid-induced neurotoxicity in vivo and in vitro. J. Ethnopharmacol. 2011, 138, 135–141. [Google Scholar] [CrossRef]
- Lee, S.T.; Chu, K.; Sim, J.Y.; Heo, J.H.; Kim, M. Panax Ginseng Enhances Cognitive Performance in Alzheimer Disease. Alzheimer Dis. Assoc. Disord. 2008, 22, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.H.; Lee, S.T.; Chu, K.; Oh, M.J.; Park, H.J.; Shim, J.Y.; Kim, M. An open-label trial of Korean red ginseng as an adjuvant treatment for cognitive impairment in patients with Alzheimer’s disease. Eur. J. Neurol. 2008, 15, 865–868. [Google Scholar] [CrossRef] [PubMed]


| Study | Study Type | Study Details | Intervention | Results |
|---|---|---|---|---|
| [69] | Review | - | - |
|
| [72] | Review | - | - |
|
| [73] | Review | - | - |
|
| Animal models | ||||
| [74] | RCS | 16 (8) Rats (omega-3 depleted) | 300 mg/kg/day DHA |
|
| Human patients | ||||
| [75] | Correlational study | 55 participants | Self-reported consumption of omega-3 FA |
|
| [77] | Correlational study | 84 participants (19 AD; 10 OD; 36 CIND; 19 Control) | FA analysis in blood plasma |
|
| [78] | Longitudinal study | 8085 healthy adults | - |
|
| [79] | Correlational study | 148 participants (29 M) Control: 45 (9 M) | FA analysis in blood plasma |
|
| Study | Study Type | Study Details | Intervention | Results |
|---|---|---|---|---|
| [89] | Experimental study | Se nanoparticles on a bovine albumin and glucose medium |
| |
| [90] | Correlational study | Vascular endothelial cell cultures |
| |
| Animal models | ||||
| [91] | RCS | Mice | Normal cage vs. spin wheel cage |
|
| Mouse neural cell culture | Selenium-controlled growth medium |
| ||
| Mice | Sodium selenite infusion |
| ||
| Mice | Sepp1 knockout |
| ||
| Mice | Endhotelin-1-induced hippocampal lesion |
| ||
| Human patients | ||||
| [92] | RCT | 40 probable AD patients | 1 mg Na2SeO4/day vs. 10 mg Na2SeO4/day vs. placebo |
|
| [87] | RCT | 79 patients with AD | 200 μg /day Se vs. 200 μg /day Se + probiotics vs. placebo |
|
| Study | Study Type | Study Details | Intervention | Results |
|---|---|---|---|---|
| Animal models | ||||
| [108] | Experimental study | Rat cerebellar granule cell culture | 100 μg /mL GB extract |
|
| [109] | Experimental study | Rat cerebellar granule cell culture | 100 μg /mL GB extract |
|
| [112] | Experimental study | Rat and mice isolated mitochondria | 10, 100, and 500 μg /mL GB extract |
|
| [113] | Experimental study | APP/PS1 mice | 50 mg/kg GB extract ad libitum |
|
| [114] | Experimental study | TgAPP/PS1 mice | 100 mg/kg GB extract ad libitum |
|
| [122] | Experimental study | APP/PS1 double-TG mice | 100 and 500 mg/kg/day, p.o. for 12 days, white PG powder |
|
| [123] | Experimental study | ICR mice + 2 mg/kg, i.p. scopolamine | 200 mg/kg, p.o., red, white, and black PG extracts |
|
| [124] | Experimental study | Sprague Dawley rats tauopathy model | Pre-treatment: 10 mg/kg for 7 days red PG extract |
|
| [119] | Experimental study | In vitro rat cells | 4–20 mM curcumin |
|
| Human patients | ||||
| [90] | Meta-analysis | 1207 AD, VD | 120–240 mg GB extract |
|
| [120] | Meta-analysis | 32 studies—human cells, animal cells, and animal models | 0.1–200 μL extract/7.5–400 mg powder CL, 5 days–6 months |
|
| [106] | Experimental study | Microglial cell culture | GB extract 10–90 μg/mL |
|
| [107] | Experimental study | Human keratinocyte cell culture | 1 mg/mL GB extract |
|
| [125] | Experimental study | AD patients | PG powder (4.5 g/d), 12 weeks |
|
| [126] | Experimental study | AD patients | Red Korean PG powder, 9 g/day, 12 weeks |
|
| [119] | Experimental study | Human macrophages from AD patients | 36.8 mg/mL curcuminoids |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palimariciuc, M.; Balmus, I.-M.; Gireadă, B.; Ciobica, A.; Chiriță, R.; Iordache, A.-C.; Apostu, M.; Dobrin, R.P. The Quest for Neurodegenerative Disease Treatment—Focusing on Alzheimer’s Disease Personalised Diets. Curr. Issues Mol. Biol. 2023, 45, 1519-1535. https://doi.org/10.3390/cimb45020098
Palimariciuc M, Balmus I-M, Gireadă B, Ciobica A, Chiriță R, Iordache A-C, Apostu M, Dobrin RP. The Quest for Neurodegenerative Disease Treatment—Focusing on Alzheimer’s Disease Personalised Diets. Current Issues in Molecular Biology. 2023; 45(2):1519-1535. https://doi.org/10.3390/cimb45020098
Chicago/Turabian StylePalimariciuc, Matei, Ioana-Miruna Balmus, Bogdan Gireadă, Alin Ciobica, Roxana Chiriță, Alin-Constantin Iordache, Mihai Apostu, and Romeo Petru Dobrin. 2023. "The Quest for Neurodegenerative Disease Treatment—Focusing on Alzheimer’s Disease Personalised Diets" Current Issues in Molecular Biology 45, no. 2: 1519-1535. https://doi.org/10.3390/cimb45020098