

Ozoile Reduces the LPS-Induced Inflammatory Response in Colonic Epithelial Cells and THP-1 Monocytes
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Ozoile Preparation
2.3. Cell Culture and Treatment
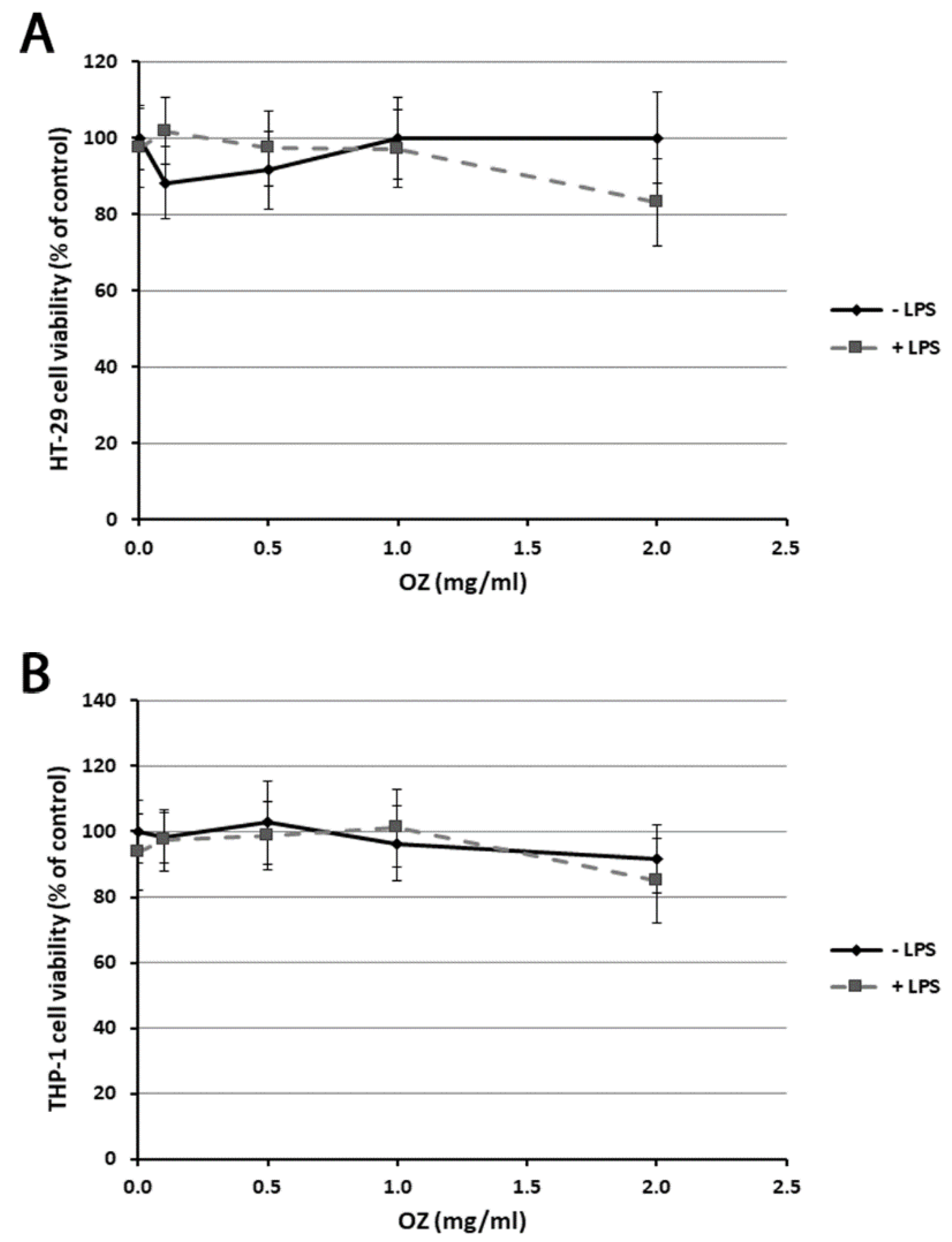
2.4. Cell Viability Assay
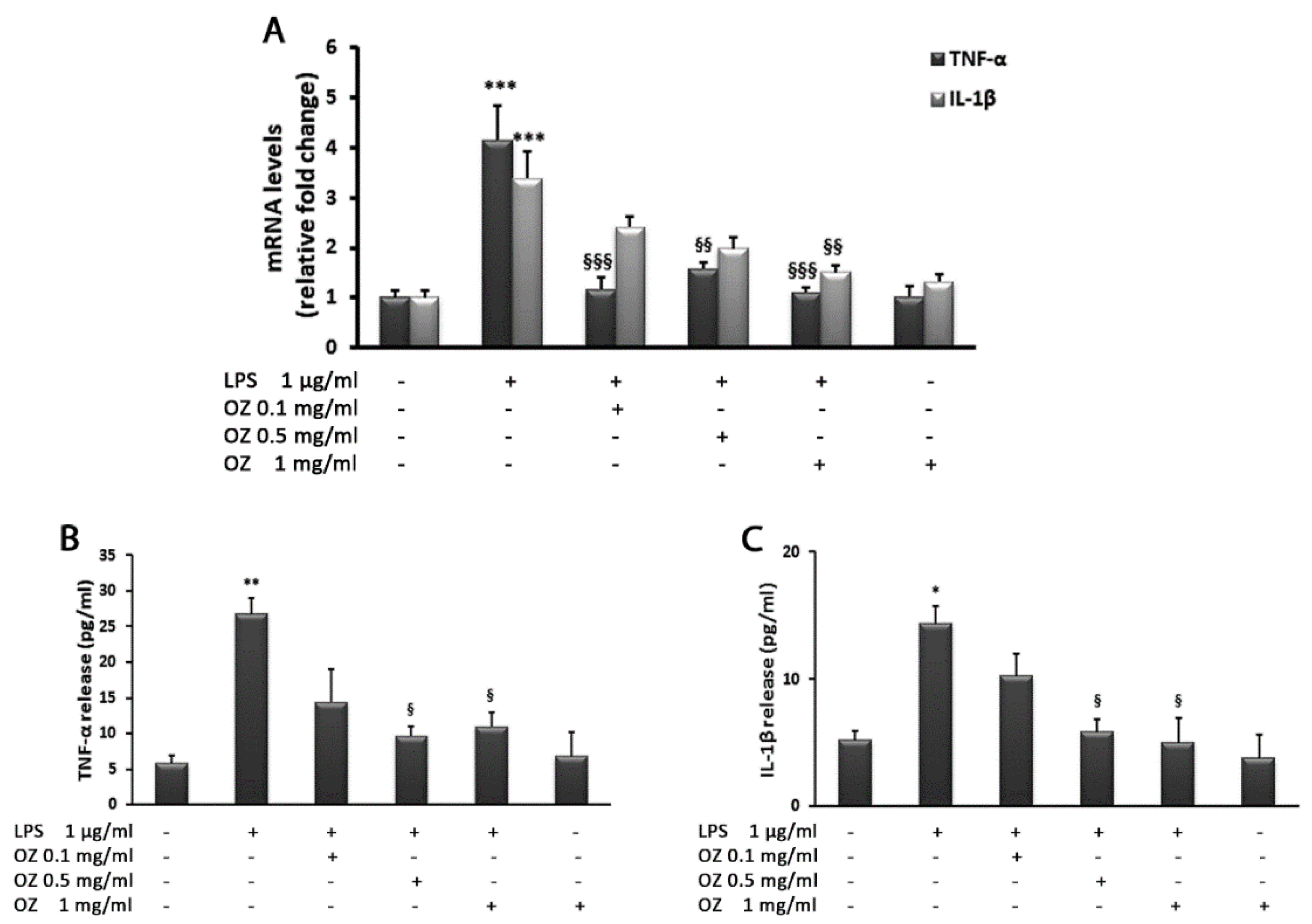
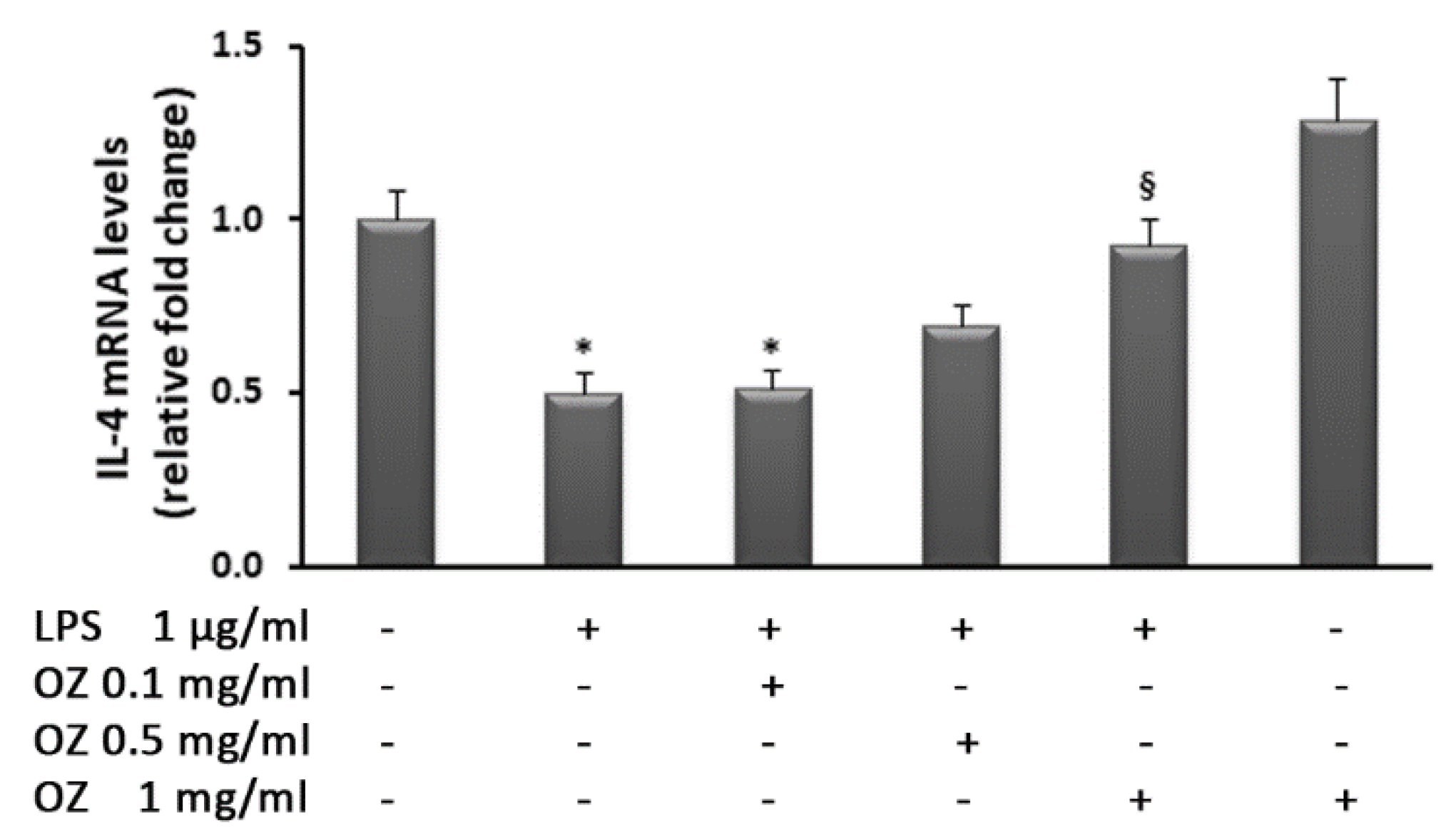
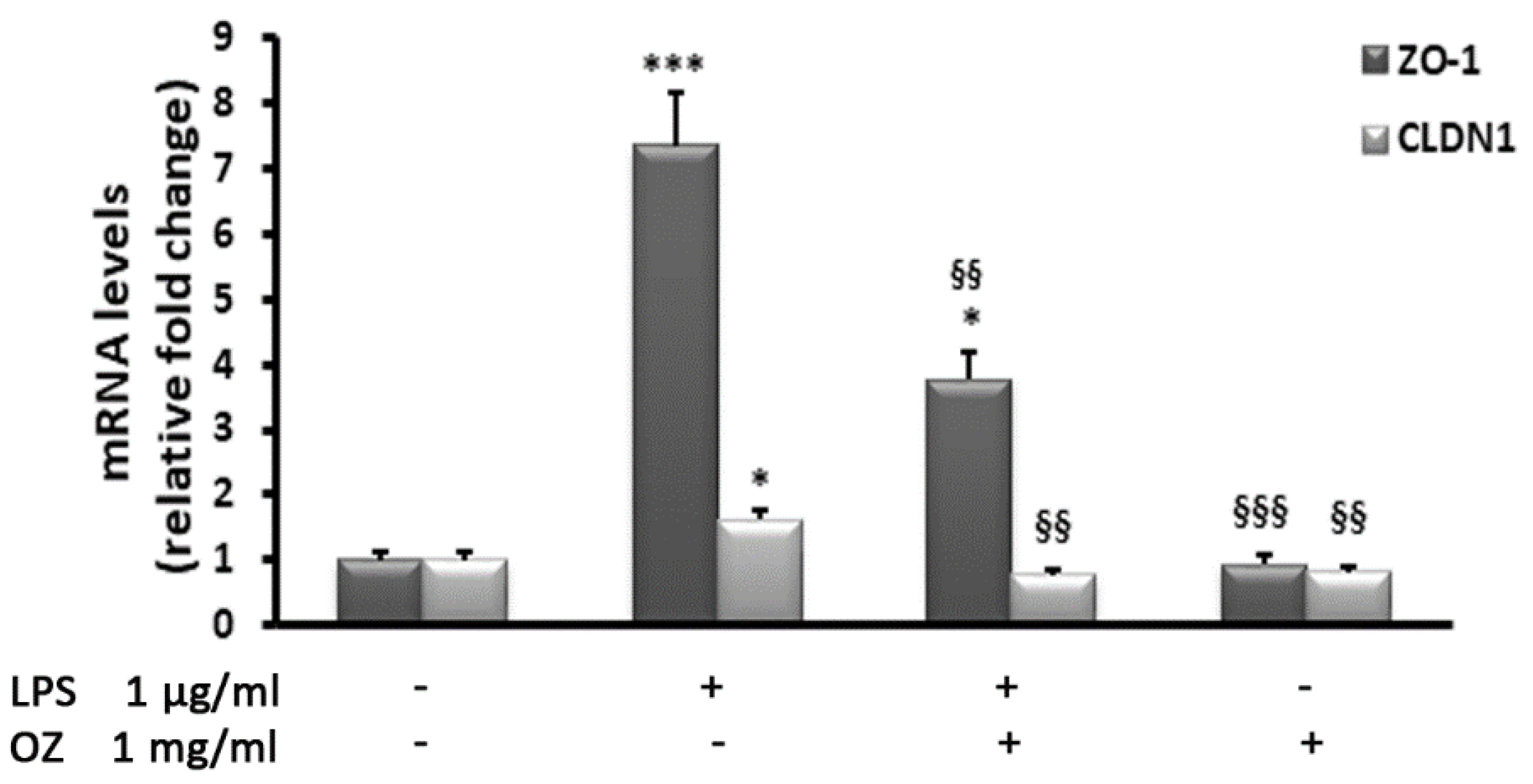
2.5. Gene Expression Analysis by Real-Time PCR
2.6. Western Blotting
2.7. Evaluation of Cytokine Secretion by ELISA
2.8. Statistical Analysis
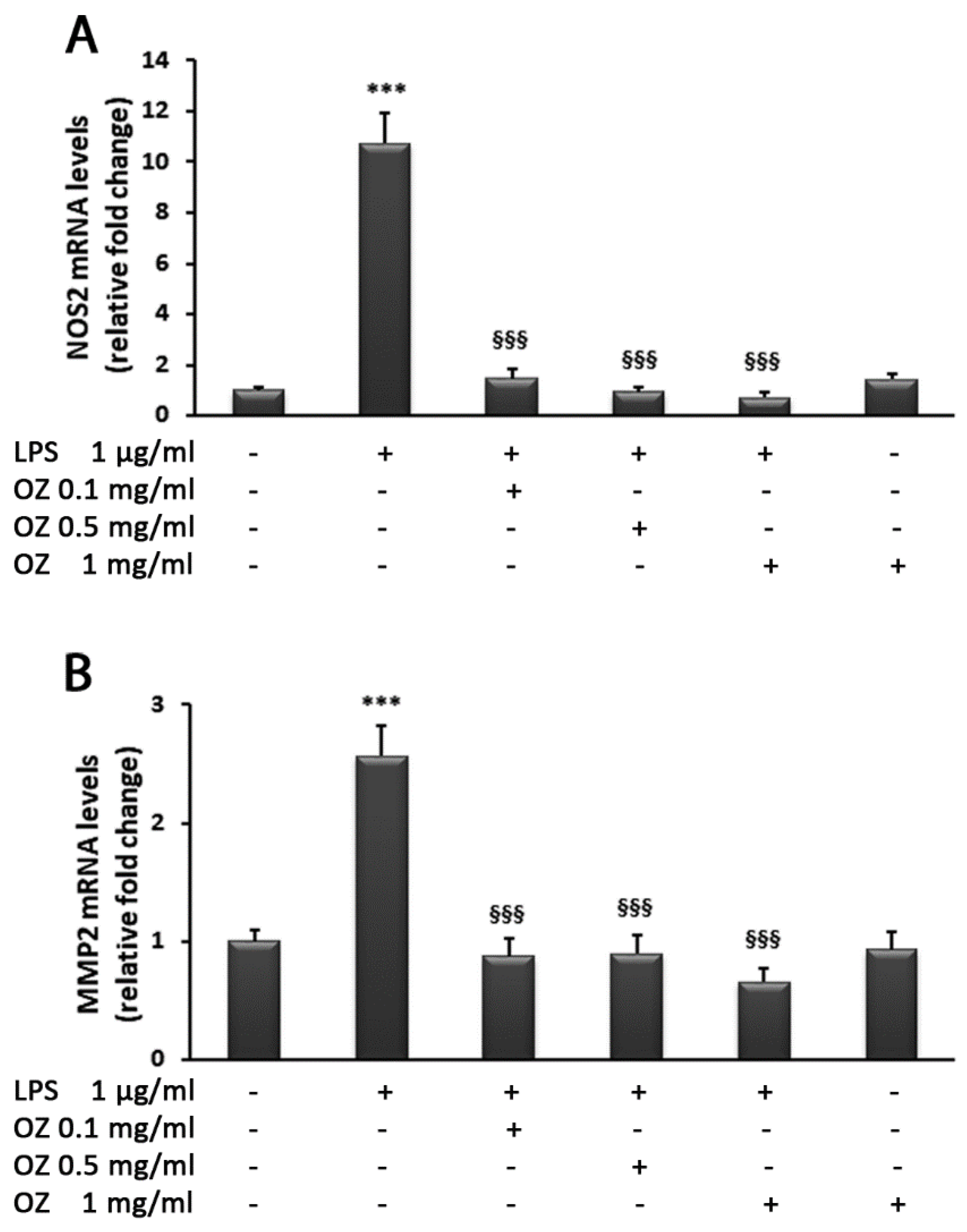
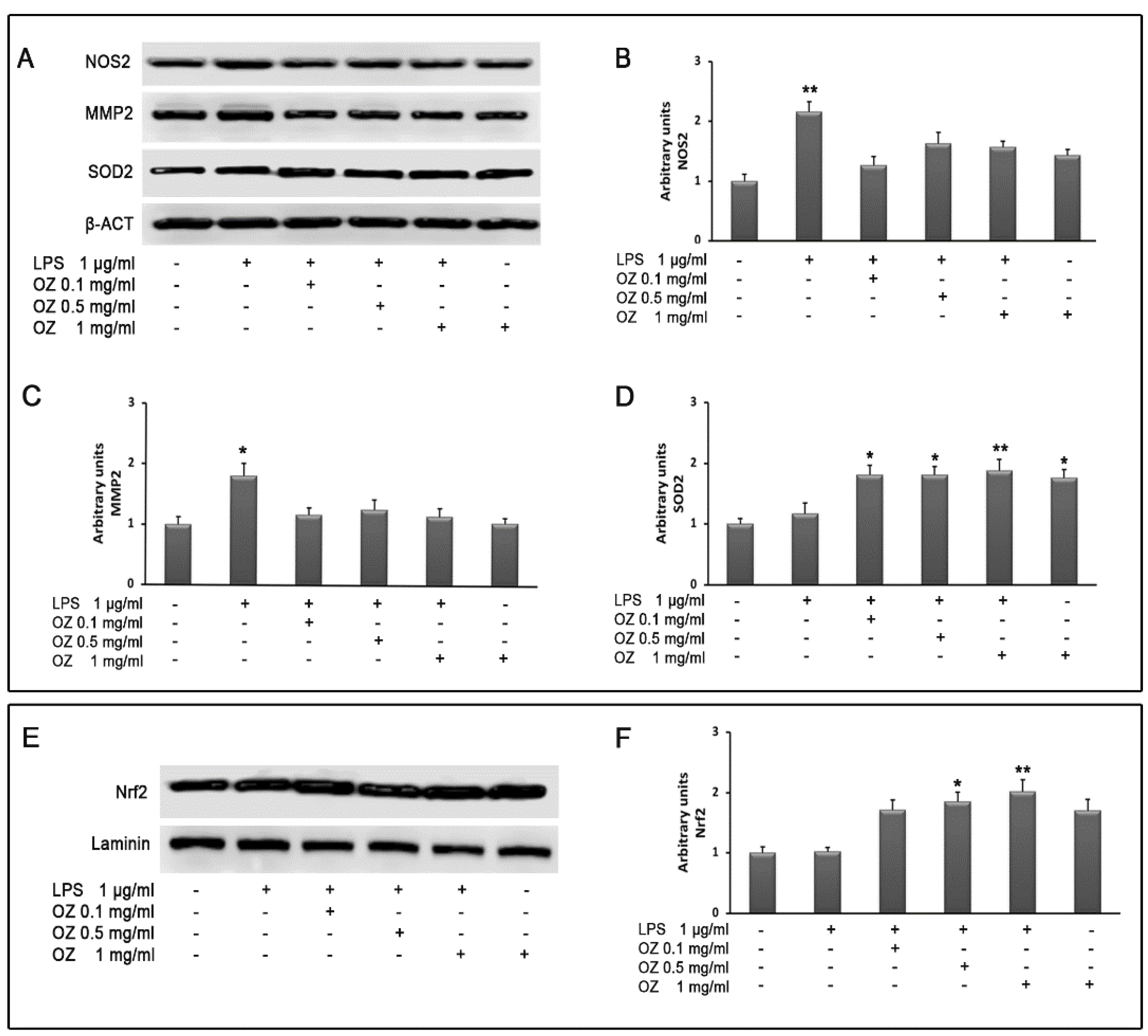
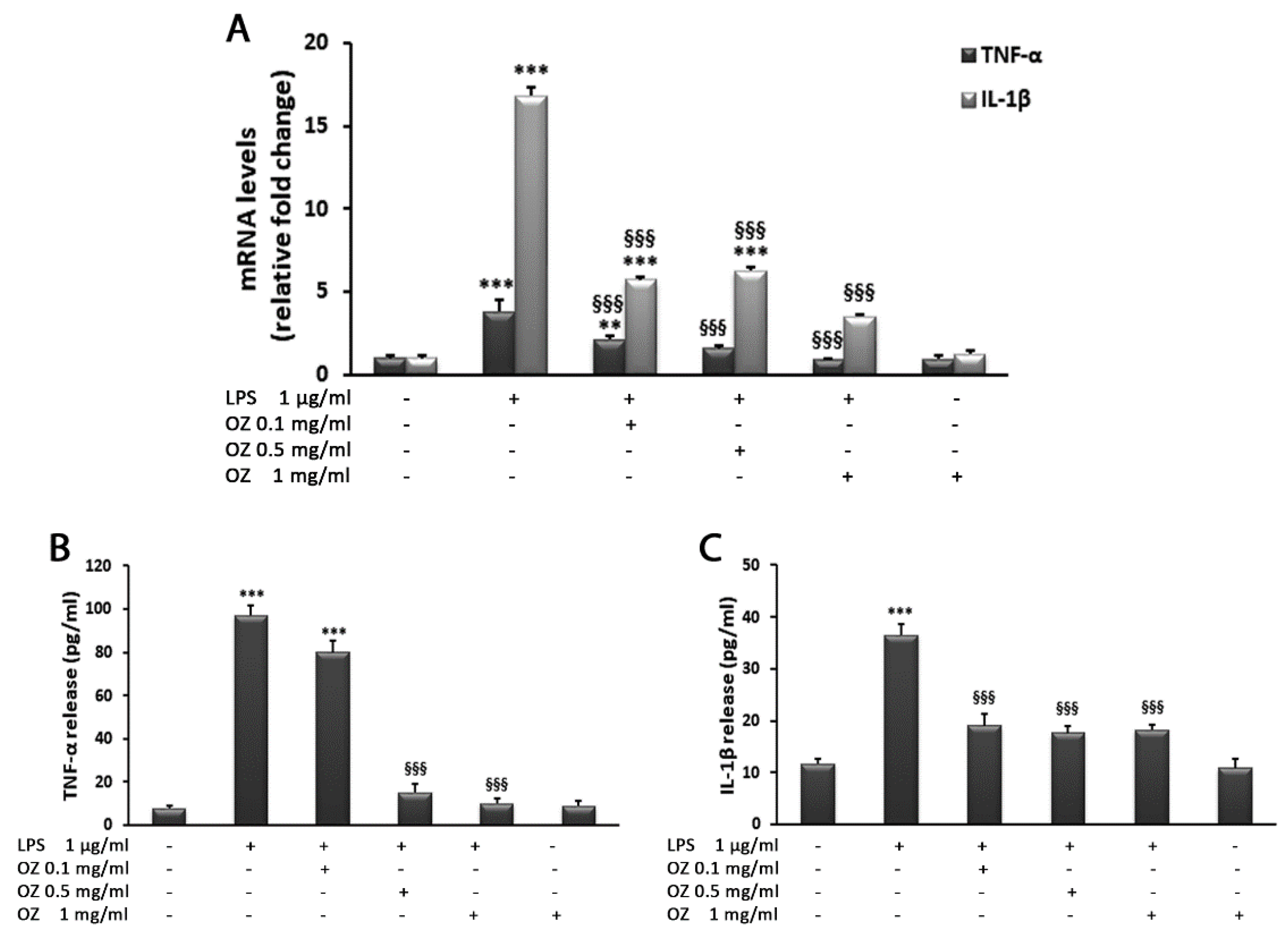
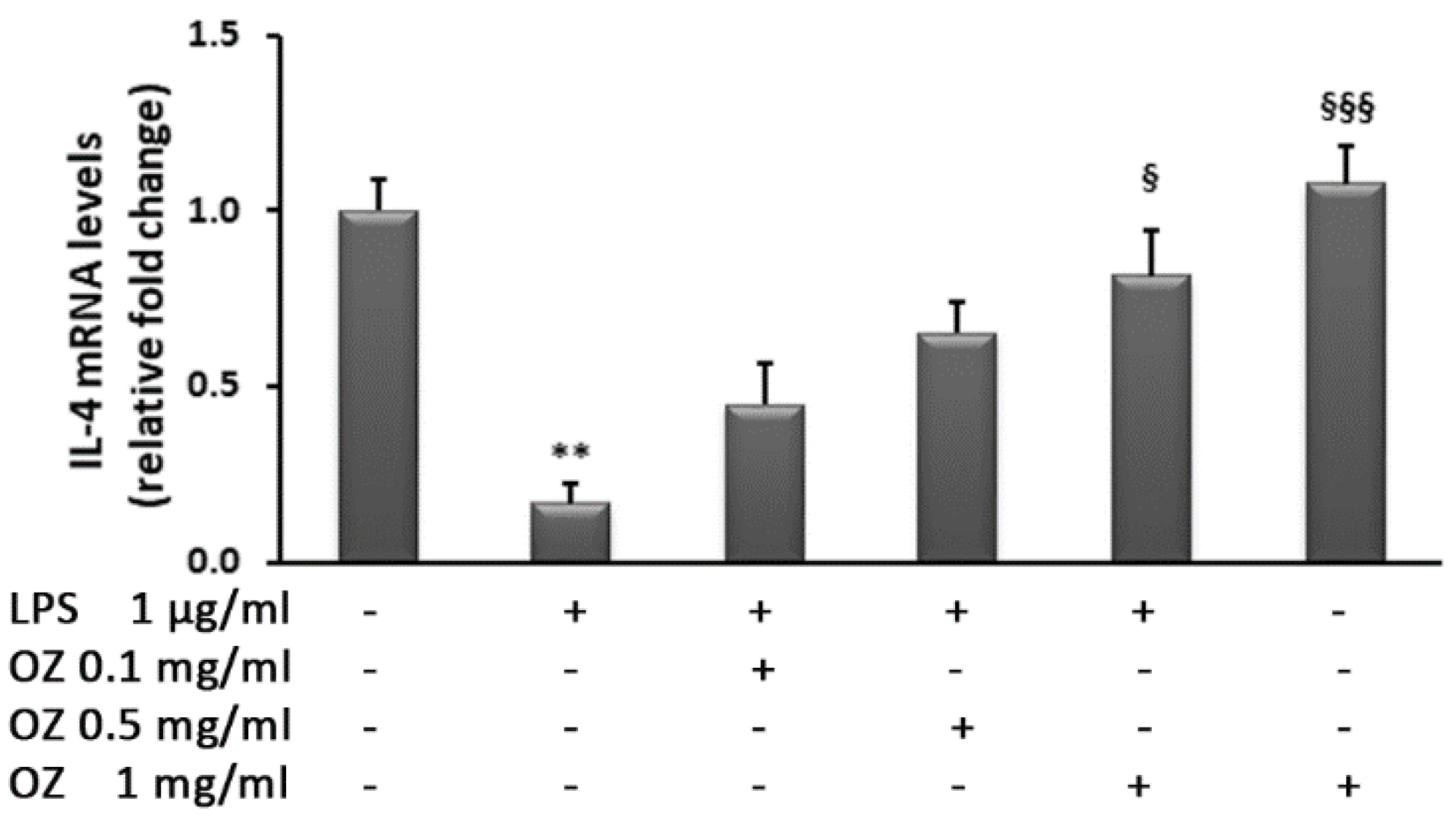
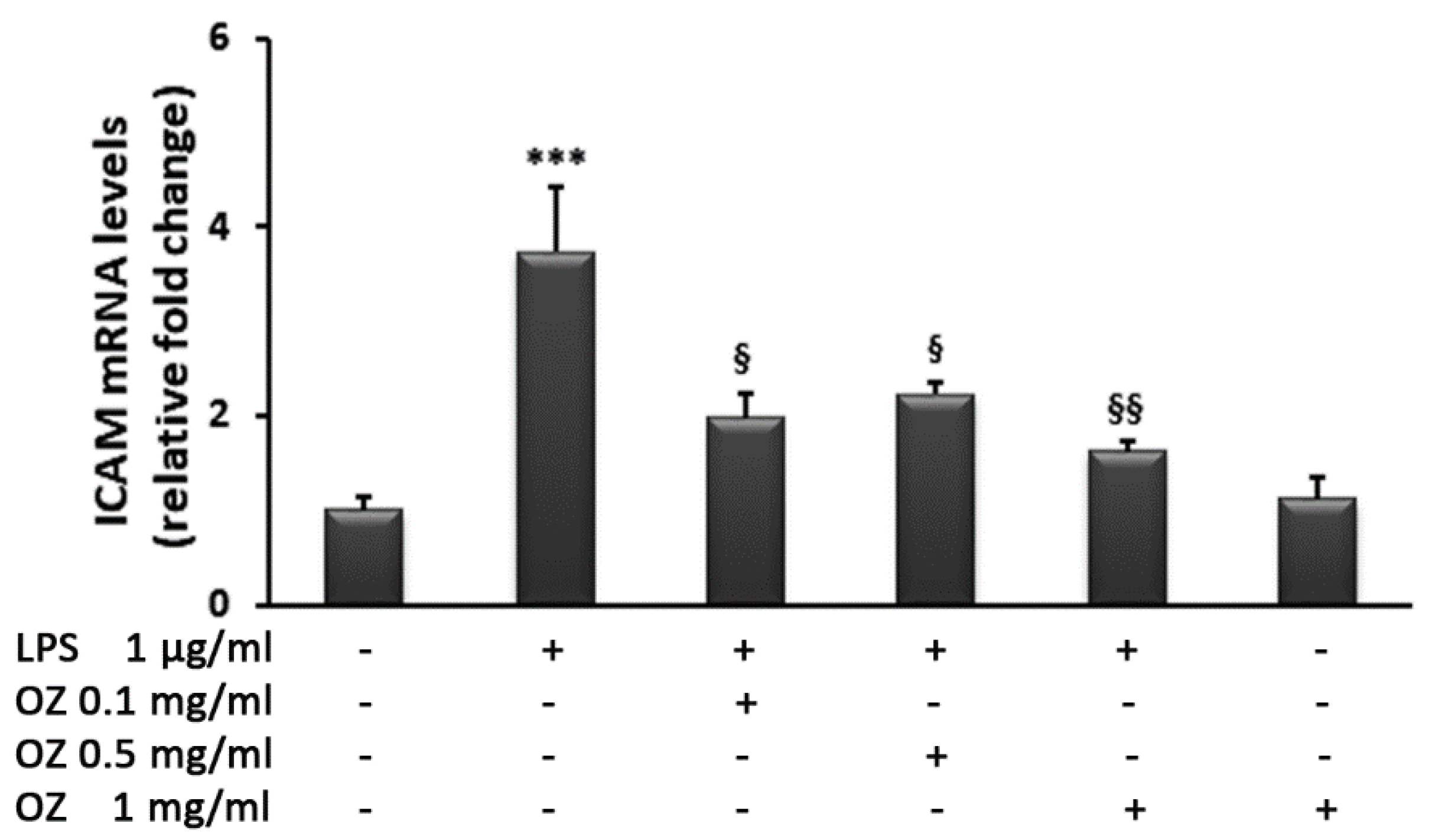
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gianchecchi, E.; Fierabracci, A. Recent Advances on Microbiota Involvement in the Pathogenesis of Autoimmunity. Int. J. Mol. Sci. 2019, 20, 283. [Google Scholar] [CrossRef] [PubMed]
- Bramuzzo, M.; De Carlo, C.; Arrigo, S.; Pavanello, P.M.; Canaletti, C.; Giudici, F.; Agrusti, A.; Martelossi, S.; Di Leo, G.; Barbi, E. Parental Psychological Factors and Quality of Life of Children with Inflammatory Bowel Disease. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Siegel, C.A.; Bernstein, C.N. Identifying Patients with Inflammatory Bowel Diseases at High vs Low Risk of Complications. Clin. Gastroenterol. Hepatol. 2019, 18, 1261–1267. [Google Scholar] [CrossRef]
- Jang, H.J.; Kang, B.; Choe, B.H. The difference in extraintestinal manifestations of inflammatory bowel disease for children and adults. Transl. Pediatr. 2019, 8, 4–15. [Google Scholar] [CrossRef]
- Hruz, P.; Juillerat, P.; Kullak-Ublick, G.A.; Schoepfer, A.M.; Mantzaris, G.J.; Rogler, G. Management of the Elderly Inflammatory Bowel Disease Patient. Digestion 2020, 14, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Devos, S.A.; Van Den Bossche, N.; De Vos, M.; Naeyaert, J.M. Adverse skin reactions to anti-TNF-alpha monoclonal antibody therapy. Dermatology 2003, 206, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Salaga, M.; Zatorski, H.; Sobczak, M.; Chen, C.; Fichna, J. Chinese herbal medicines in the treatment of IBD and colorectal cancer: A review. Curr. Treat. Options Oncol. 2014, 15, 405–420. [Google Scholar] [CrossRef]
- Maioli, T.U.; Trindade, L.M.; Souza, A.; Torres, L.; Andrade, M.E.R.; Cardoso, V.N.; Generoso, S.V. Non-pharmacologic strategies for the management of intestinal inflammation. Biomed. Pharmacother. 2022, 145, 112414. [Google Scholar] [CrossRef]
- Galie, M.; Covi, V.; Tabaracci, G.; Malatesta, M. The Role of Nrf2 in the Antioxidant Cellular Response to Medical Ozone Exposure. Int. J. Mol. Sci. 2019, 20, 4009. [Google Scholar] [CrossRef]
- Sadatullah, S.; Mohamed, N.H.; Razak, F.A. The antimicrobial effect of 0.1 ppm ozonated water on 24-hour plaque microorganisms in situ. Braz. Oral Res. 2012, 26, 126–131. [Google Scholar] [CrossRef]
- Sechi, L.A.; Lezcano, I.; Nunez, N.; Espim, M.; Dupre, I.; Pinna, A.; Molicotti, P.; Fadda, G.; Zanetti, S. Antibacterial activity of ozonized sunflower oil (Oleozon). J. Appl. Microbiol. 2001, 90, 279–284. [Google Scholar] [CrossRef]
- Uysal, B. Ozonated olive oils and the troubles. J. Intercult. Ethnopharmacol. 2014, 3, 49–50. [Google Scholar] [CrossRef]
- Elvis, A.M.; Ekta, J.S. Ozone therapy: A clinical review. J. Nat. Sci. Biol. Med. 2011, 2, 66–70. [Google Scholar] [CrossRef]
- Holz, O.; Tal-Singer, R.; Kanniess, F.; Simpson, K.J.; Gibson, A.; Vessey, R.S.; Janicki, S.; Magnussen, H.; Jorres, R.A.; Richter, K. Validation of the human ozone challenge model as a tool for assessing anti-inflammatory drugs in early development. J. Clin. Pharmacol. 2005, 45, 498–503. [Google Scholar] [CrossRef]
- Braidy, N.; Izadi, M.; Sureda, A.; Jonaidi-Jafari, N.; Banki, A.; Nabavi, S.F.; Nabavi, S.M. Therapeutic relevance of ozone therapy in degenerative diseases: Focus on diabetes and spinal pain. J. Cell. Physiol. 2018, 233, 2705–2714. [Google Scholar] [CrossRef]
- Currò, M.; Russo, T.; Ferlazzo, N.; Caccamo, D.; Antonuccio, P.; Arena, S.; Parisi, S.; Perrone, P.; Ientile, R.; Romeo, C.; et al. Anti-Inflammatory and Tissue Regenerative Effects of Topical Treatment with Ozonated Olive Oil/Vitamin E Acetate in Balanitis Xerotica Obliterans. Molecules 2018, 23, 645. [Google Scholar] [CrossRef]
- Russo, T.; Curro, M.; Ferlazzo, N.; Caccamo, D.; Perrone, P.; Arena, S.; Antonelli, E.; Antonuccio, P.; Ientile, R.; Romeo, C.; et al. Stable Ozonides with Vitamin E Acetate versus Corticosteroid in the Treatment of Lichen Sclerosus in Foreskin: Evaluation of Effects on Inflammation. Urol. Int. 2019, 103, 459–465. [Google Scholar] [CrossRef]
- Ugazio, E.; Tullio, V.; Binello, A.; Tagliapietra, S.; Dosio, F. Ozonated Oils as Antimicrobial Systems in Topical Applications. Their Characterization, Current Applications, and Advances in Improved Delivery Techniques. Molecules 2020, 25, 334. [Google Scholar] [CrossRef]
- Kato, Y.; Sakoh, M.; Nagai, T.; Yoshida, A.; Ishida, H.; Inoue, N.; Yanagita, T.; Nagao, K. Ozonated Olive Oil Alleviates Hepatic Steatosis in Obese Zucker (fa/fa) Rats. J. Oleo Sci. 2022, 71, 599–607. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.; Song, S.; Zhu, B.; Zhao, L.; Jiang, J.; Liu, N.; Wang, J.; Chen, X. Anti-inflammatory activity and structural identification of a sulfated polysaccharide CLGP4 from Caulerpa lentillifera. Int. J. Biol. Macromol. 2020, 146, 931–938. [Google Scholar] [CrossRef]
- Byun, E.B.; Kim, W.S.; Sung, N.Y.; Byun, E.H. Epigallocatechin-3-Gallate Regulates Anti-Inflammatory Action through 67-kDa Laminin Receptor-Mediated Tollip Signaling Induction in Lipopolysaccharide-Stimulated Human Intestinal Epithelial Cells. Cell. Physiol. Biochem. 2018, 46, 2072–2081. [Google Scholar] [CrossRef]
- Yurinskaya, M.M.; Krasnov, G.S.; Kulikova, D.A.; Zatsepina, O.G.; Vinokurov, M.G.; Chuvakova, L.N.; Rezvykh, A.P.; Funikov, S.Y.; Morozov, A.V.; Evgen’ev, M.B. H2S counteracts proinflammatory effects of LPS through modulation of multiple pathways in human cells. Inflamm. Res. 2020, 69, 481–495. [Google Scholar] [CrossRef]
- Mapa, M.S.T.; Araujo, M.; Zhao, Y.; Flynn, T.; Sprando, J.; Wiesenfeld, P.; Sprando, R.L.; Mossoba, M.E. A method to dissolve 3-MCPD mono- and di-esters in aqueous cell culture media. MethodsX 2020, 7, 100774. [Google Scholar] [CrossRef]
- Rizzo, V.; Ferlazzo, N.; Curro, M.; Isola, G.; Matarese, M.; Bertuccio, M.P.; Caccamo, D.; Matarese, G.; Ientile, R. Baicalin-Induced Autophagy Preserved LPS-Stimulated Intestinal Cells from Inflammation and Alterations of Paracellular Permeability. Int. J. Mol. Sci. 2021, 22, 2315. [Google Scholar] [CrossRef]
- Risitano, R.; Curro, M.; Cirmi, S.; Ferlazzo, N.; Campiglia, P.; Caccamo, D.; Ientile, R.; Navarra, M. Flavonoid fraction of Bergamot juice reduces LPS-induced inflammatory response through SIRT1-mediated NF-kappaB inhibition in THP-1 monocytes. PLoS ONE 2014, 9, e107431. [Google Scholar] [CrossRef]
- Stolfi, C.; Maresca, C.; Monteleone, G.; Laudisi, F. Implication of Intestinal Barrier Dysfunction in Gut Dysbiosis and Diseases. Biomedicines 2022, 10, 289. [Google Scholar] [CrossRef]
- Di Mauro, R.; Cantarella, G.; Bernardini, R.; Di Rosa, M.; Barbagallo, I.; Distefano, A.; Longhitano, L.; Vicario, N.; Nicolosi, D.; Lazzarino, G.; et al. The Biochemical and Pharmacological Properties of Ozone: The Smell of Protection in Acute and Chronic Diseases. Int. J. Mol. Sci. 2019, 20, 634. [Google Scholar] [CrossRef]
- Kim, H.S.; Noh, S.U.; Han, Y.W.; Kim, K.M.; Kang, H.; Kim, H.O.; Park, Y.M. Therapeutic effects of topical application of ozone on acute cutaneous wound healing. J. Korean Med. Sci. 2009, 24, 368–374. [Google Scholar] [CrossRef]
- Pai, S.A.; Gagangras, S.A.; Kulkarni, S.S.; Majumdar, A.S. Potential of ozonated sesame oil to augment wound healing in rats. Indian J. Pharm. Sci. 2014, 76, 87–92. [Google Scholar]
- Altinel, O.; Demirbas, S.; Cakir, E.; Yaman, H.; Ozerhan, I.H.; Duran, E.; Cayci, T.; Akgul, E.O.; Ersoz, N.; Uysal, B.; et al. Comparison of hyperbaric oxygen and medical ozone therapies in a rat model of experimental distal colitis. Scand. J. Clin. Lab. Invest. 2011, 71, 185–192. [Google Scholar] [CrossRef]
- Geng, Y.; Wang, W.; Ma, Q.; Peng, L.Q.; Liang, Z.H. Ozone therapy combined with sulfasalazine delivered via a colon therapy system for treatment of ulcerative colitis. Nan Fang Yi Ke Da Xue Xue Bao 2010, 30, 2683–2685. [Google Scholar]
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef]
- Yamamoto-Furusho, J.K. Inflammatory bowel disease therapy: Blockade of cytokines and cytokine signaling pathways. Curr. Opin. Gastroenterol. 2018, 34, 187–193. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Eslamian, G.; Ardehali, S.H.; Hajimohammadebrahim-Ketabforoush, M.; Vahdat Shariatpanahi, Z. Association of intestinal permeability with admission vitamin D deficiency in patients who are critically ill. J. Investig. Med. 2020, 68, 397–402. [Google Scholar] [CrossRef]
- Malickova, K.; Francova, I.; Lukas, M.; Kolar, M.; Kralikova, E.; Bortlik, M.; Duricova, D.; Stepankova, L.; Zvolska, K.; Pankova, A.; et al. Fecal zonulin is elevated in Crohn’s disease and in cigarette smokers. Pract. Lab. Med. 2017, 9, 39–44. [Google Scholar] [CrossRef]
- Sturgeon, C.; Lan, J.; Fasano, A. Zonulin transgenic mice show altered gut permeability and increased morbidity/mortality in the DSS colitis model. Ann. N. Y. Acad. Sci. 2017, 1397, 130–142. [Google Scholar] [CrossRef]
- Weber, C.R.; Nalle, S.C.; Tretiakova, M.; Rubin, D.T.; Turner, J.R. Claudin-1 and claudin-2 expression is elevated in inflammatory bowel disease and may contribute to early neoplastic transformation. Lab. Invest. 2008, 88, 1110–1120. [Google Scholar] [CrossRef]
- Guihot, G.; Guimbaud, R.; Bertrand, V.; Narcy-Lambare, B.; Couturier, D.; Duee, P.H.; Chaussade, S.; Blachier, F. Inducible nitric oxide synthase activity in colon biopsies from inflammatory areas: Correlation with inflammation intensity in patients with ulcerative colitis but not with Crohn’s disease. Amino Acids 2000, 18, 229–237. [Google Scholar] [CrossRef]
- Rafa, H.; Saoula, H.; Belkhelfa, M.; Medjeber, O.; Soufli, I.; Toumi, R.; de Launoit, Y.; Morales, O.; Nakmouche, M.; Delhem, N.; et al. IL-23/IL-17A axis correlates with the nitric oxide pathway in inflammatory bowel disease: Immunomodulatory effect of retinoic acid. J. Interferon Cytokine Res. 2013, 33, 355–368. [Google Scholar] [CrossRef]
- Soufli, I.; Toumi, R.; Rafa, H.; Touil-Boukoffa, C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J. Gastrointest. Pharmacol. Ther. 2016, 7, 353–360. [Google Scholar] [CrossRef]
- Mao, J.W.; Tang, H.Y.; Tan, X.Y.; Wang, Y.D. Effect of Etiasa on the expression of matrix metalloproteinase-2 and tumor necrosis factor-alpha in a rat model of ulcerative colitis. Mol. Med. Rep. 2012, 6, 996–1000. [Google Scholar] [CrossRef]
- Sim, W.H.; Wagner, J.; Cameron, D.J.; Catto-Smith, A.G.; Bishop, R.F.; Kirkwood, C.D. Expression profile of genes involved in pathogenesis of pediatric Crohn’s disease. J. Gastroenterol. Hepatol. 2012, 27, 1083–1093. [Google Scholar] [CrossRef]
- Ravi, A.; Garg, P.; Sitaraman, S.V. Matrix metalloproteinases in inflammatory bowel disease: Matrix metalloproteinases in inflammatory bowel disease: Boon or a bane? Inflamm. Bowel. Dis. 2007, 13, 97–107. [Google Scholar] [CrossRef]
- Galie, M.; Costanzo, M.; Nodari, A.; Boschi, F.; Calderan, L.; Mannucci, S.; Covi, V.; Tabaracci, G.; Malatesta, M. Mild ozonisation activates antioxidant cell response by the Keap1/Nrf2 dependent pathway. Free Radic. Biol. Med. 2018, 124, 114–121. [Google Scholar] [CrossRef]
- Li, W.; Kong, A.N. Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. 2009, 48, 91–104. [Google Scholar] [CrossRef]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2-an update. Free Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Sohn, D.H.; Jin, X.Y.; Kim, S.W.; Choi, S.C.; Seo, G.S. 2′,4′,6′-tris(methoxymethoxy) chalcone protects against trinitrobenzene sulfonic acid-induced colitis and blocks tumor necrosis factor-alpha-induced intestinal epithelial inflammation via heme oxygenase 1-dependent and independent pathways. Biochem. Pharmacol. 2007, 74, 870–880. [Google Scholar] [CrossRef]
- Piotrowska, M.; Swierczynski, M.; Fichna, J.; Piechota-Polanczyk, A. The Nrf2 in the pathophysiology of the intestine: Molecular mechanisms and therapeutic implications for inflammatory bowel diseases. Pharmacol. Res. 2021, 163, 105243. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.R.; Bain, C.C.; Fenton, T.M.; Kelly, A.; Brown, S.L.; Ivens, A.C.; Travis, M.A.; Cook, P.C.; MacDonald, A.S. Dynamics of Colon Monocyte and Macrophage Activation During Colitis. Front. Immunol. 2018, 9, 2764. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Lin, Y.H.; Bi, L.H.; Wang, J.D.; Bai, Y.; Liu, S.D. Effects of interleukin-4 or interleukin-10 gene therapy on trinitrobenzenesulfonic acid-induced murine colitis. BMC Gastroenterol. 2013, 13, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayme, T.S.; Leung, G.; Wang, A.; Workentine, M.L.; Rajeev, S.; Shute, A.; Callejas, B.E.; Mancini, N.; Beck, P.L.; Panaccione, R.; et al. Human interleukin-4-treated regulatory macrophages promote epithelial wound healing and reduce colitis in a mouse model. Sci. Adv. 2020, 6, eaba4376. [Google Scholar] [CrossRef] [PubMed]
- Bamias, G.; Clark, D.J.; Rivera-Nieves, J. Leukocyte traffic blockade as a therapeutic strategy in inflammatory bowel disease. Curr. Drug Targets 2013, 14, 1490–1500. [Google Scholar] [CrossRef]
- Bailey, M.J.; Naik, N.N.; Wild, L.E.; Patterson, W.B.; Alderete, T.L. Exposure to air pollutants and the gut microbiota: A potential link between exposure, obesity, and type 2 diabetes. Gut Microbes 2020, 11, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- Snow, S.J.; Broniowska, K.; Karoly, E.D.; Henriquez, A.R.; Phillips, P.M.; Ledbetter, A.D.; Schladweiler, M.C.; Miller, C.N.; Gordon, C.J.; Kodavanti, U.P. Offspring susceptibility to metabolic alterations due to maternal high-fat diet and the impact of inhaled ozone used as a stressor. Sci. Rep. 2020, 10, 16353. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer Sequence 5′ > 3′ | |
|---|---|---|
| Forward | Reverse | |
| TNF-α | GTGAGGAGGACGAACATC | GAGCCAGAAGAGGTTGAG |
| IL-1β | GCTTATTACAGTGGCAATGA | TAGTGGTGGTCGGAGATT |
| IL-4 | CCACGGACACAAGTGCGATAT | GGCAGCAAAGATGTCTGTTACG |
| NOS2 | TGACCTCCTAACAAGTAGCA | CAGCAGCAAGTTCCATCT |
| MMP2 | TGATCTTGACCAGAATACCATCGA | GGCTTGCGAGGGAAGAAGTT |
| ZO1 | TCATCCCAAATAAGAACAGAGC | GAAGAACAACCCTTTCATAAGC |
| CLDN1 | GTTGGGCTTCATTCTCGCCTT | CCTGGGCGGTCACGATGTTGTC |
| ICAM | CCCATGAAACCGAACACAC | ACTCTGTTCAGTGTGGCACC |
| β-actin | TTGTTACAGGAAGTCCCTTGCC | ATGCTATCACCTCCCCTGTGTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertuccio, M.P.; Rizzo, V.; Arena, S.; Trainito, A.; Montalto, A.S.; Caccamo, D.; Currò, M.; Romeo, C.; Impellizzeri, P. Ozoile Reduces the LPS-Induced Inflammatory Response in Colonic Epithelial Cells and THP-1 Monocytes. Curr. Issues Mol. Biol. 2023, 45, 1333-1348. https://doi.org/10.3390/cimb45020087
Bertuccio MP, Rizzo V, Arena S, Trainito A, Montalto AS, Caccamo D, Currò M, Romeo C, Impellizzeri P. Ozoile Reduces the LPS-Induced Inflammatory Response in Colonic Epithelial Cells and THP-1 Monocytes. Current Issues in Molecular Biology. 2023; 45(2):1333-1348. https://doi.org/10.3390/cimb45020087
Chicago/Turabian StyleBertuccio, Maria Paola, Valentina Rizzo, Salvatore Arena, Alessandra Trainito, Angela Simona Montalto, Daniela Caccamo, Monica Currò, Carmelo Romeo, and Pietro Impellizzeri. 2023. "Ozoile Reduces the LPS-Induced Inflammatory Response in Colonic Epithelial Cells and THP-1 Monocytes" Current Issues in Molecular Biology 45, no. 2: 1333-1348. https://doi.org/10.3390/cimb45020087