

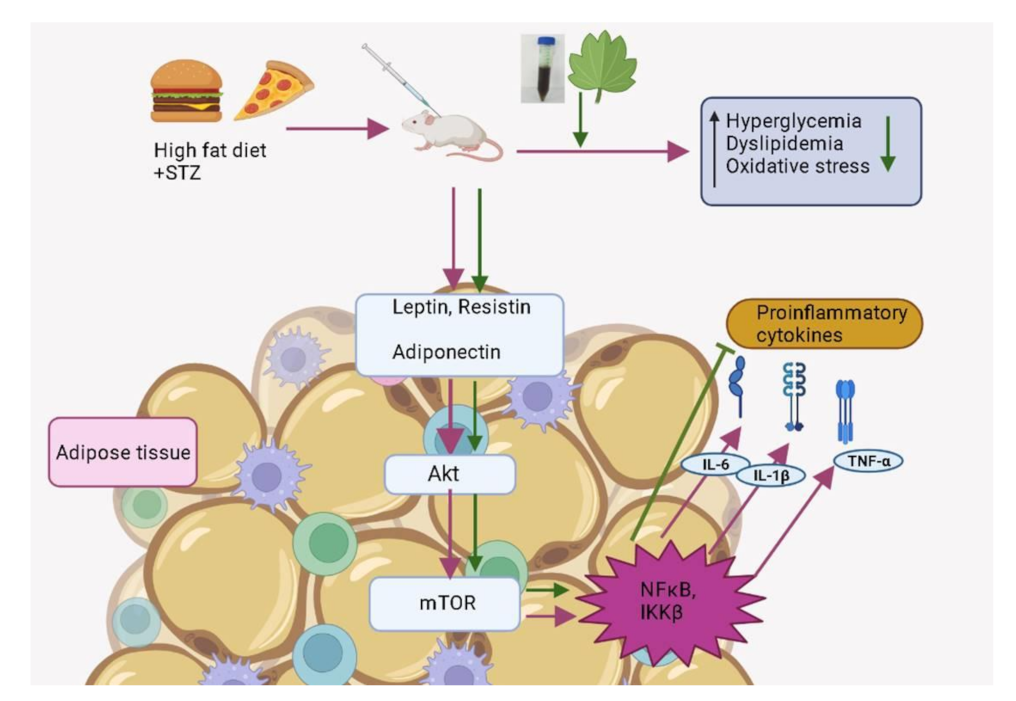
Carica Papaya Reduces High Fat Diet and Streptozotocin-Induced Development of Inflammation in Adipocyte via IL-1β/IL-6/TNF-α Mediated Signaling Mechanisms in Type-2 Diabetic Rats
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Collection and Authentication of Plant Material
2.3. Preparation of Ethanolic Extract of C. papaya
2.4. Animals
2.5. Induction of T2DM
2.6. Experimental Design
2.7. Serum Adiponectin, Leptin and Resistin
2.8. Oxidative Stress Markers
2.9. Enzymatic Antioxidants
2.10. Total RNA, cDNA Synthesis and Real-Time PCR
2.11. Histopathology
2.12. Immunohistochemical Analysis
2.13. Statistical Analysis
2.14. Molecular Docking
2.14.1. Preparation of Protein
2.14.2. Ligand Preparation
2.14.3. Molecular Docking
2.14.4. Investigation of Potential Compounds’ Molecular Dynamics and Simulation and Receptor Complex
2.14.5. Binding Energy Calculations
3. Results
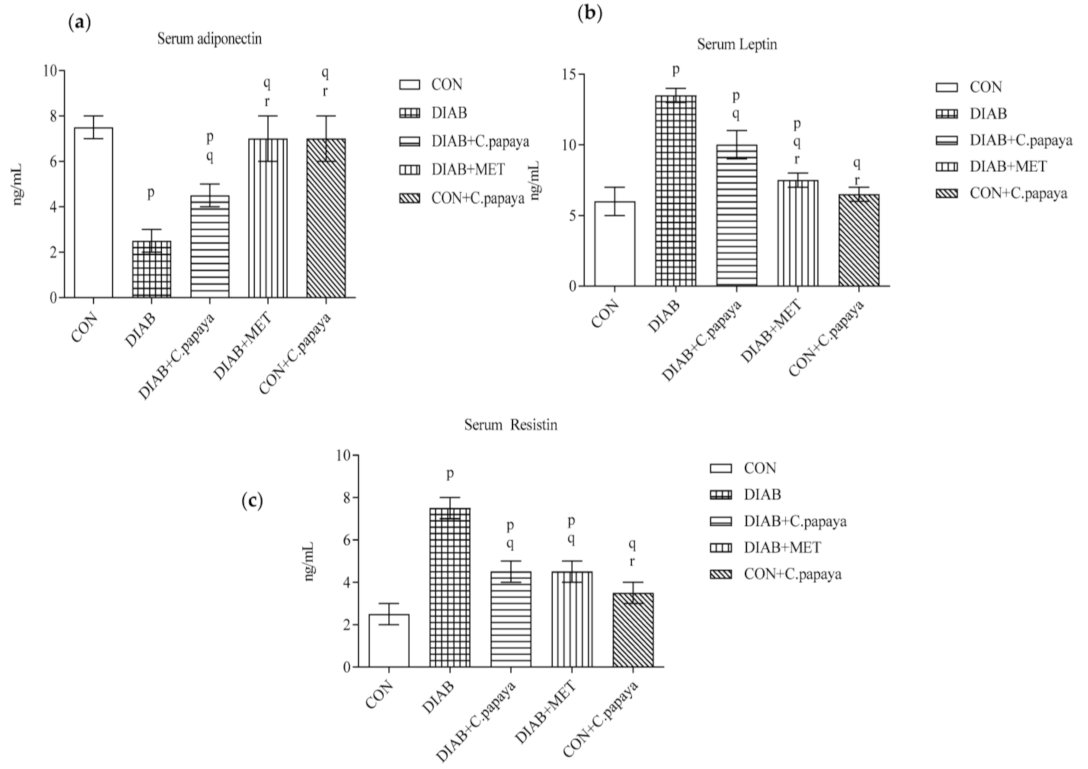
3.1. Effect of C. papaya on Serum Adiponectin, Leptin and Resistin
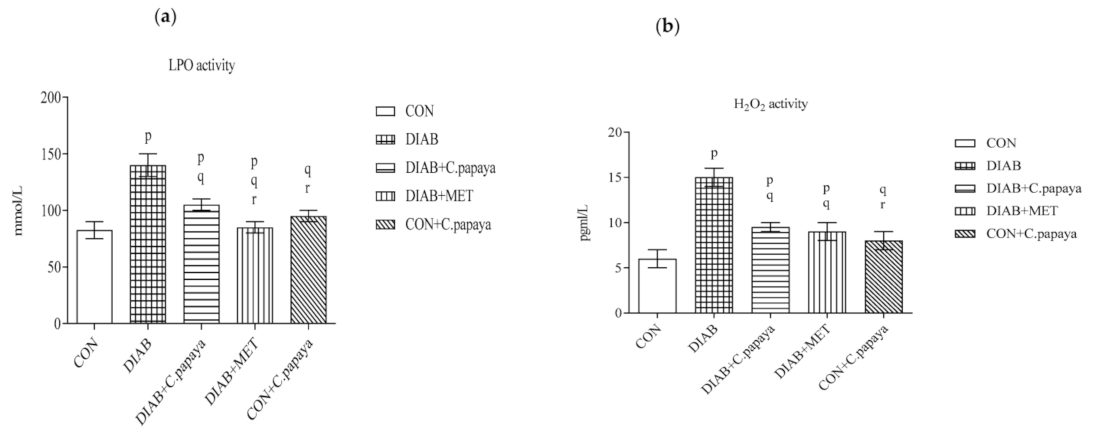
3.2. Result of C. papaya on Oxidative Stress Markers
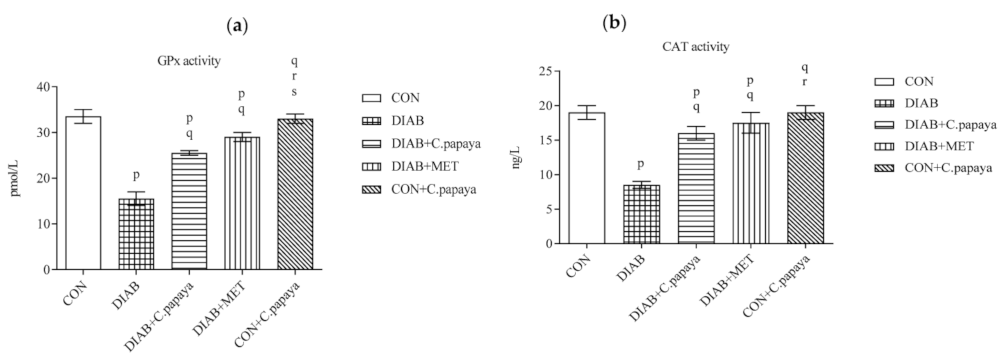
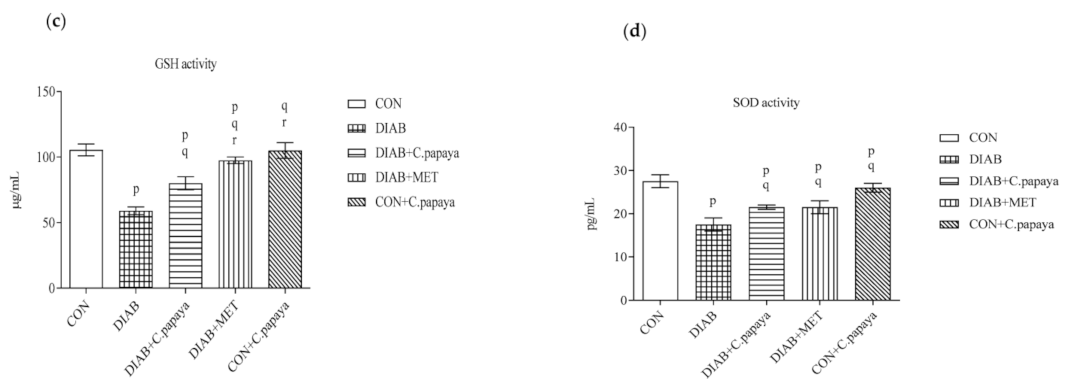
3.3. Result of C. papaya on Enzymatic Antioxidants
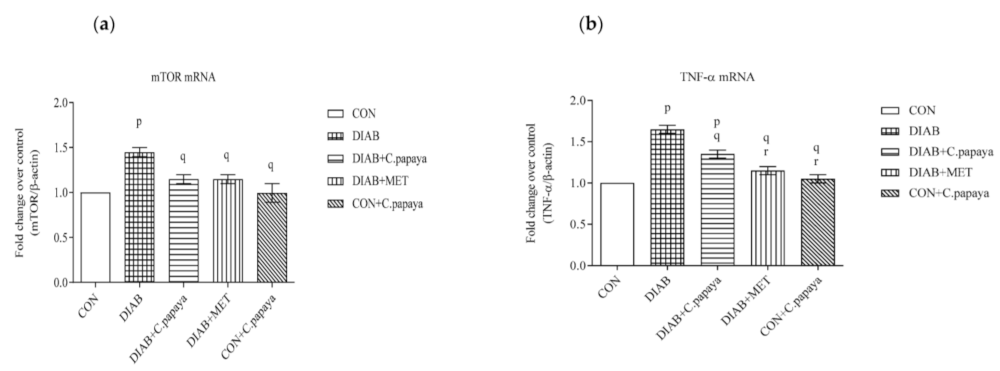
3.4. Effect of C. papaya on mRNA Expression of mTOR, TNF-α, IL-1β, IL-6 and IKKβ in Adipose Tissue
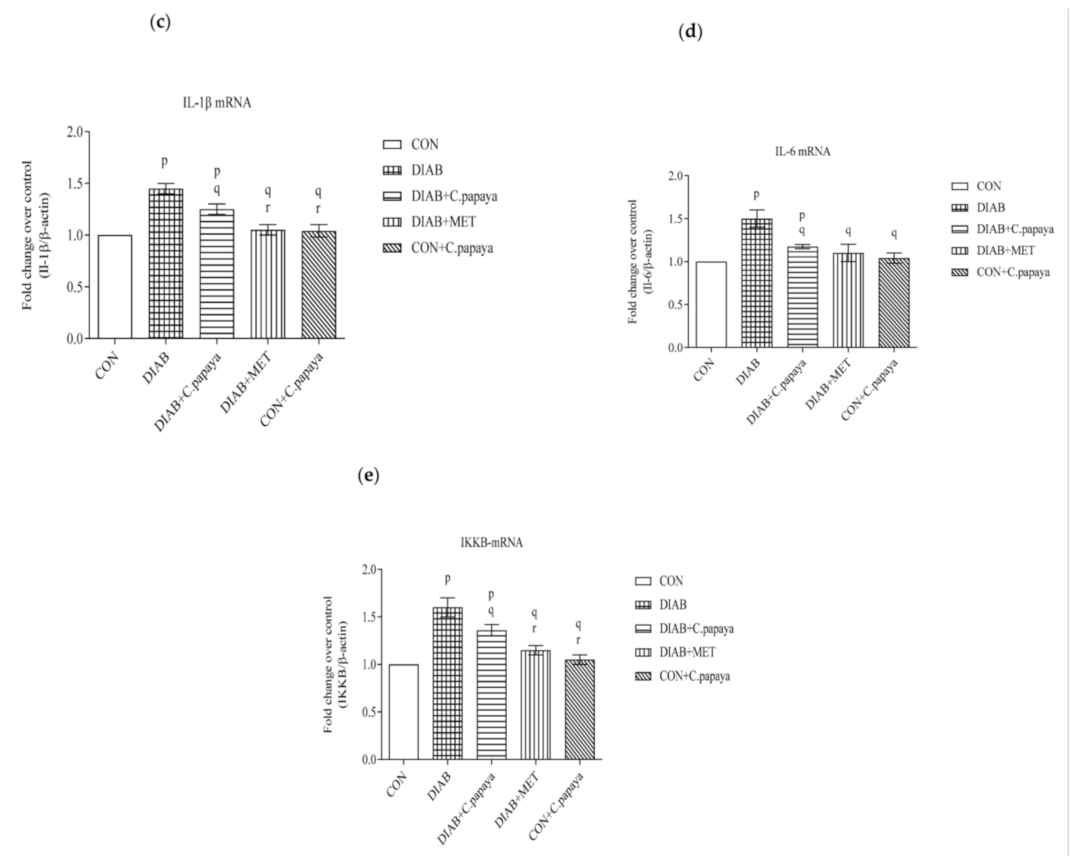
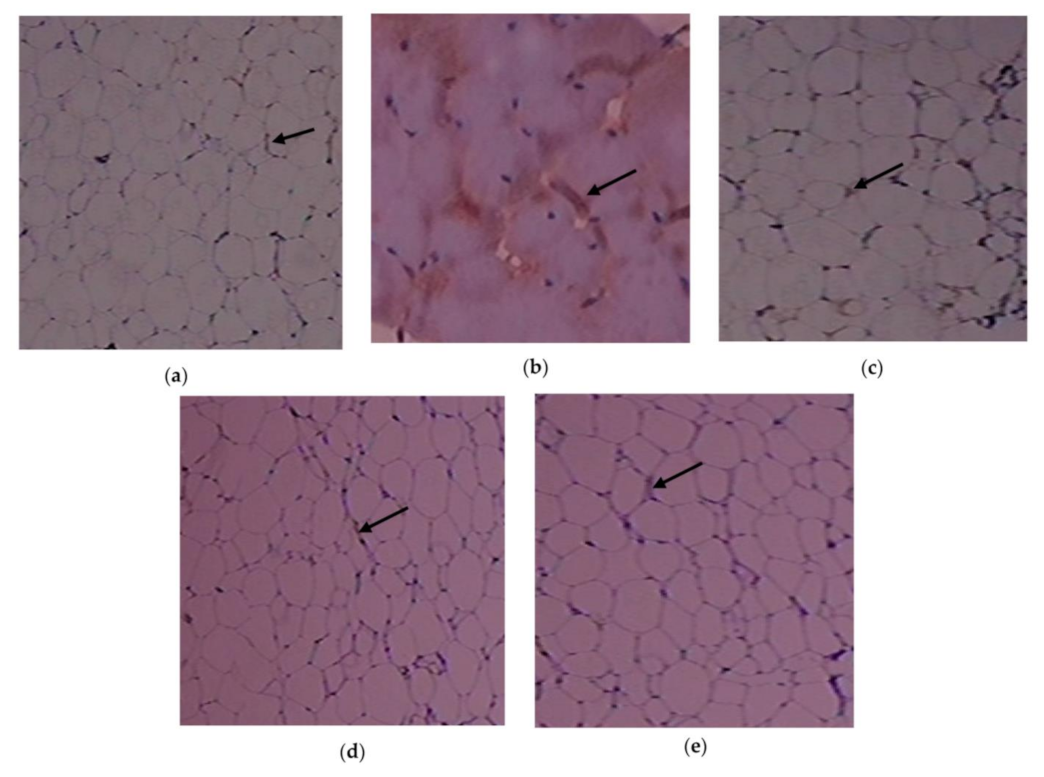
3.5. Result of C. papaya on the Histopathological Changes in the Adipose Tissue
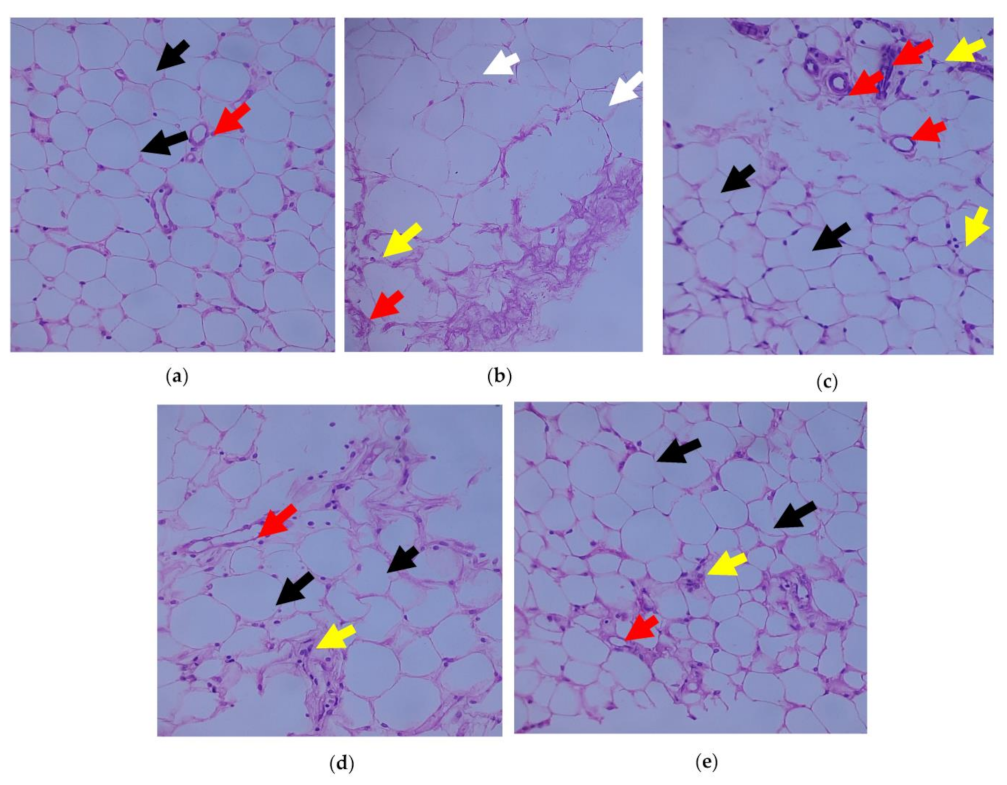
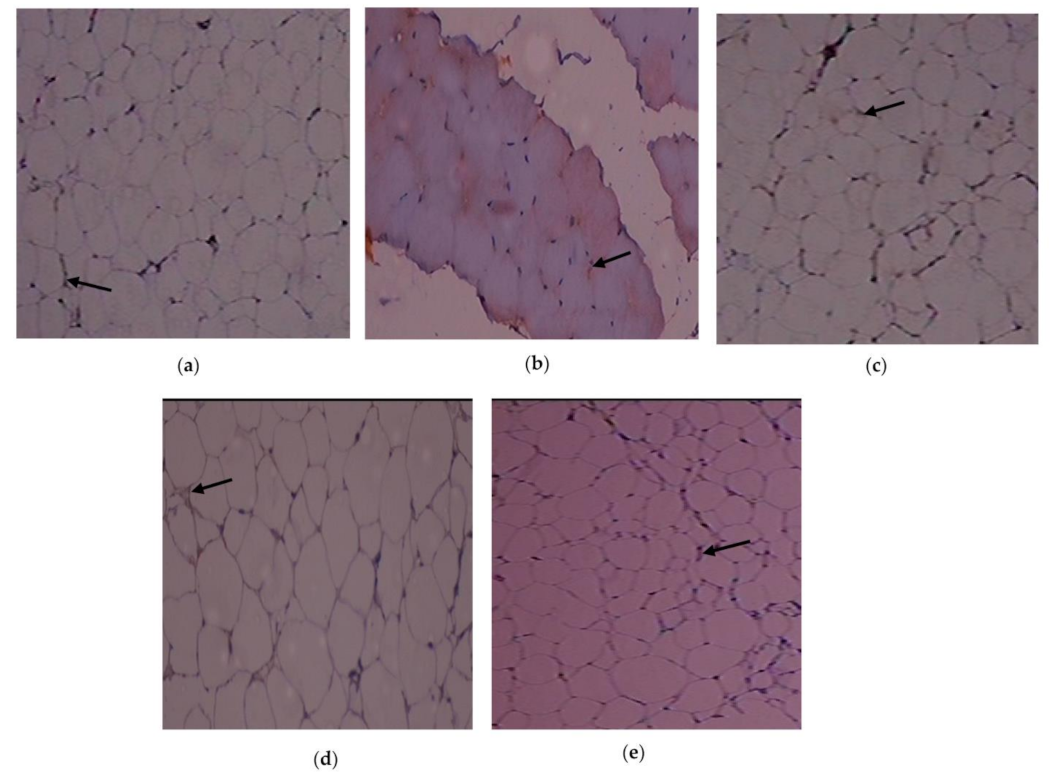
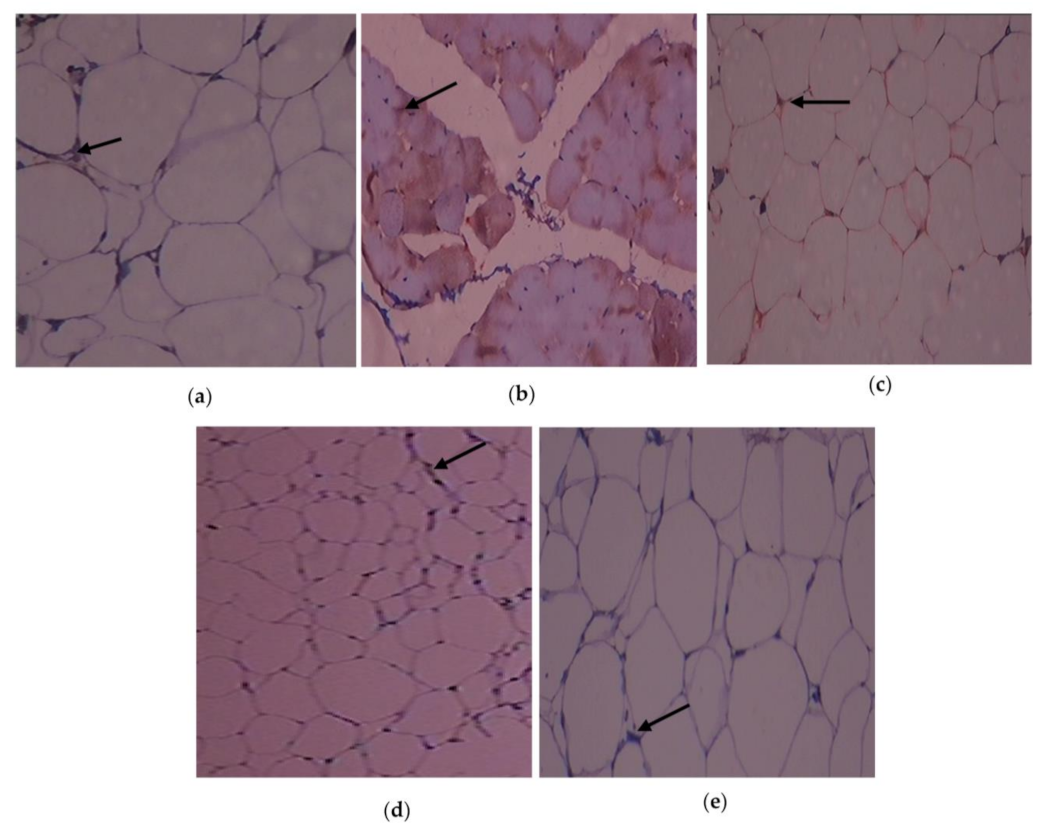
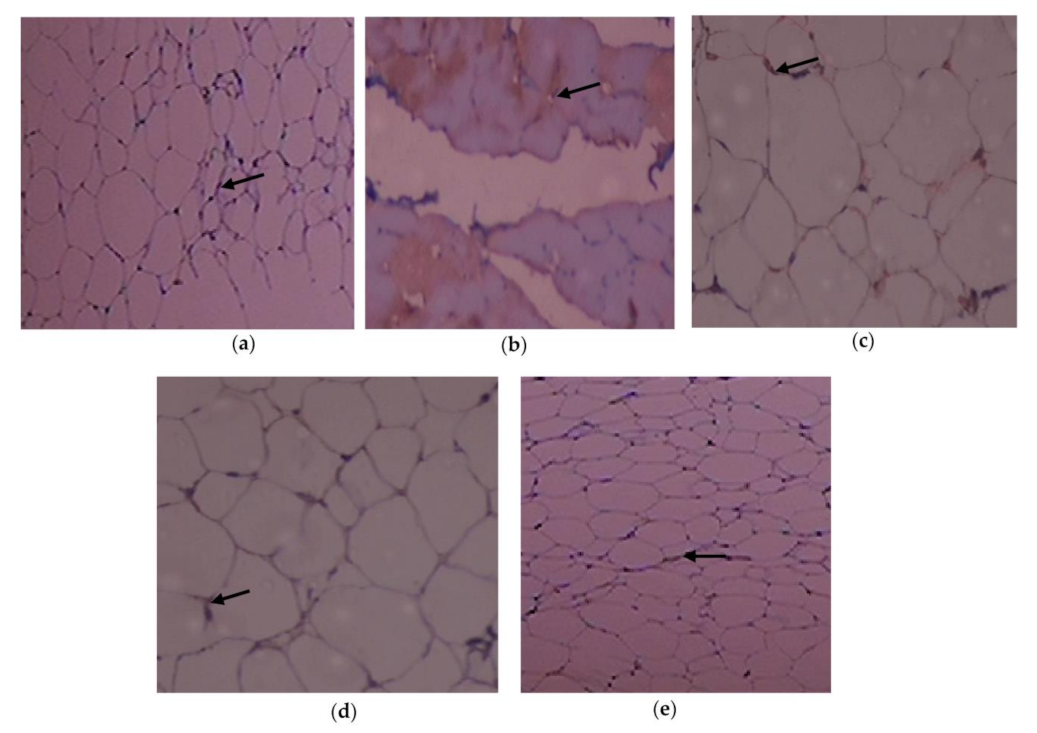
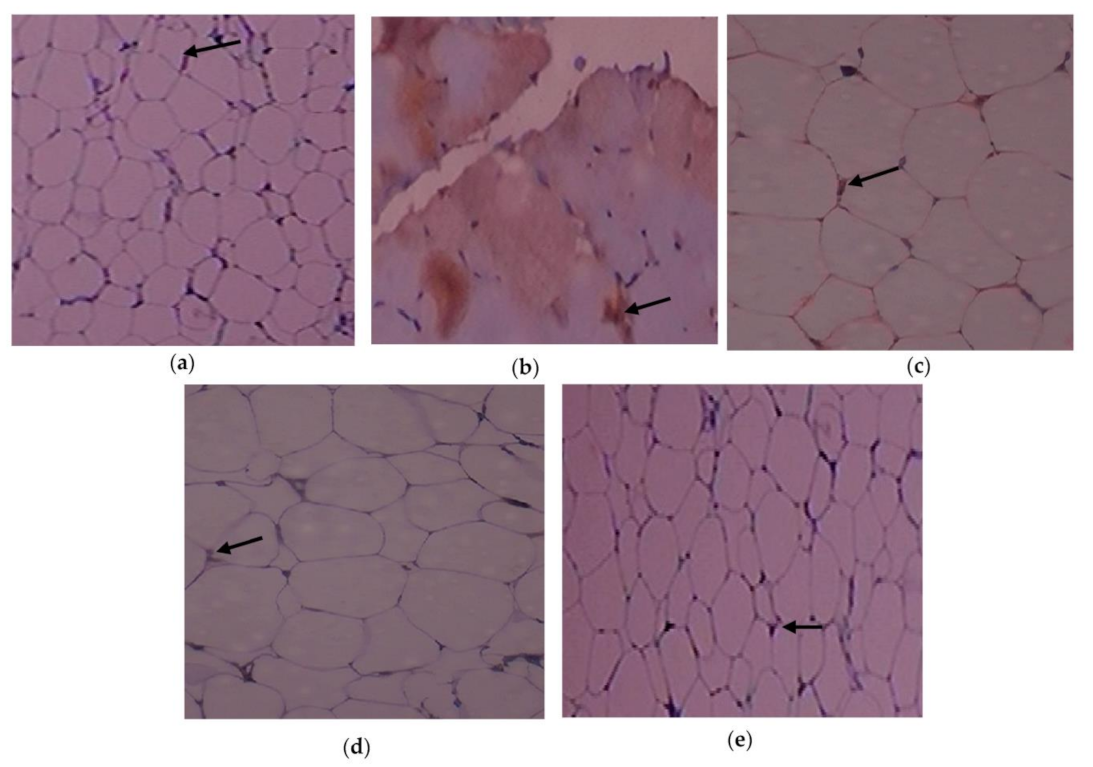
3.6. Outcome of C. papaya on the Immunohistochemical Changes in the Adipose Tissue
3.7. Molecular Docking
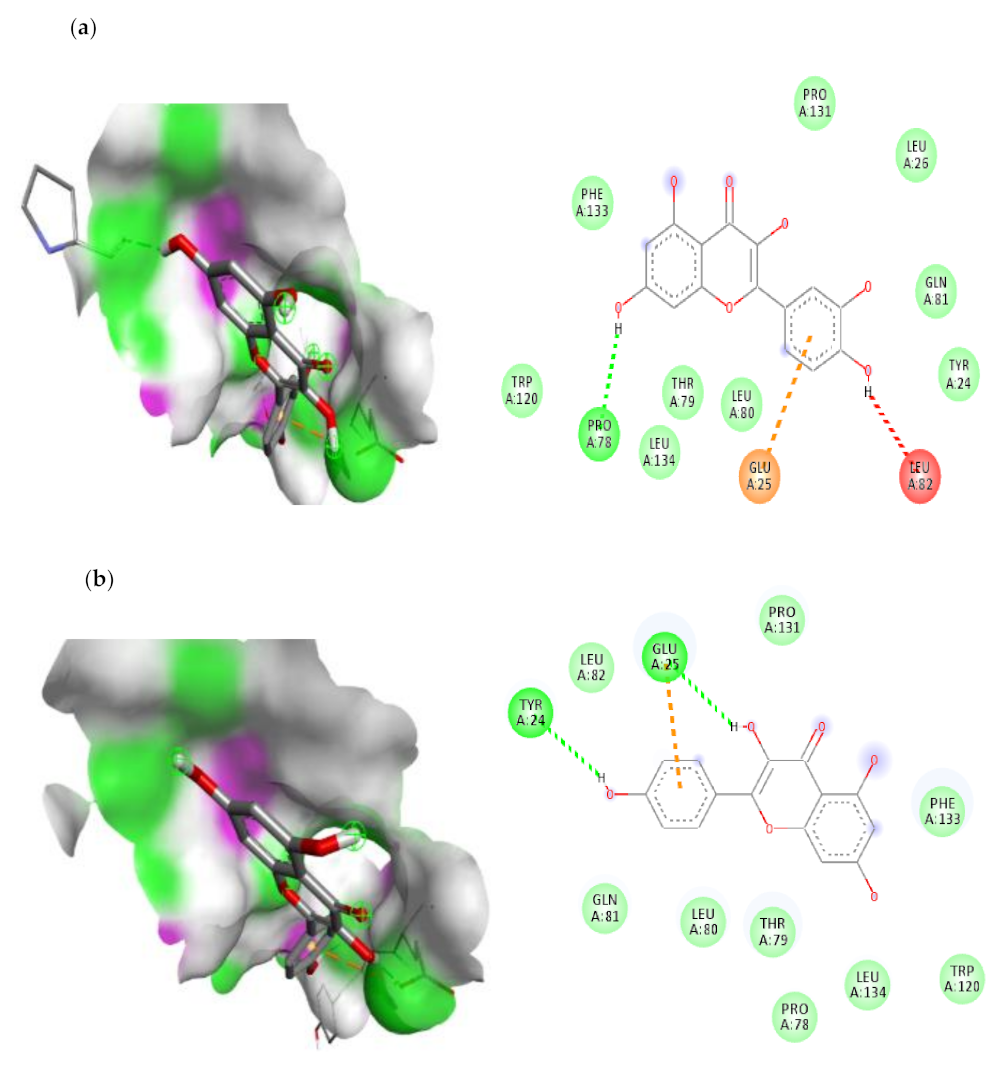
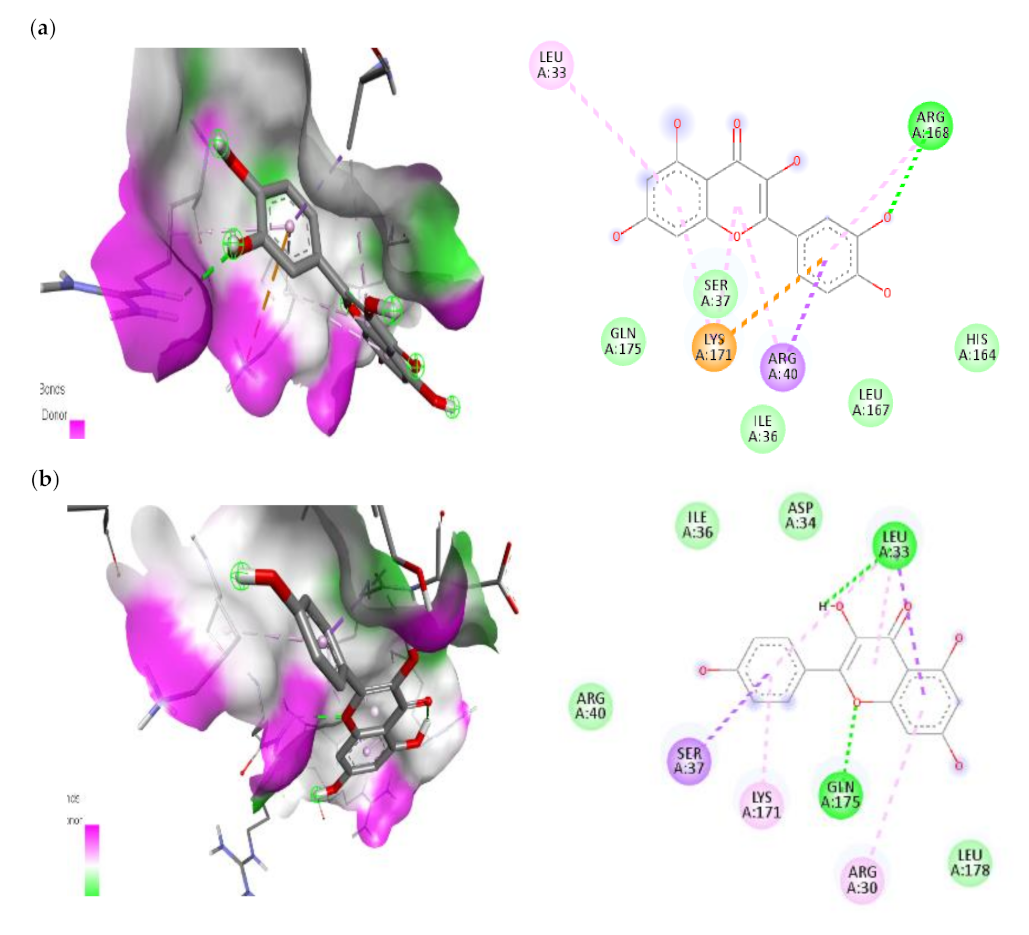
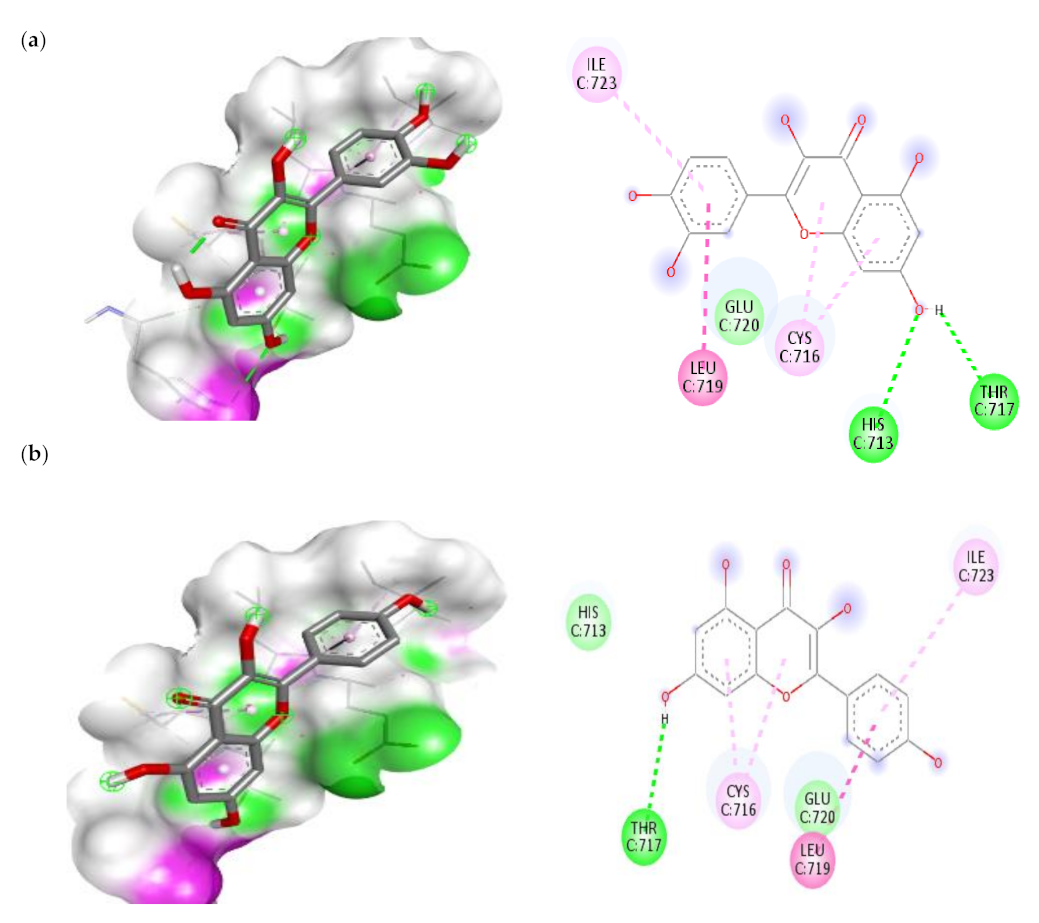
3.7.1. Molecular Docking Results of IL-1β, IL-6 and IKKβ
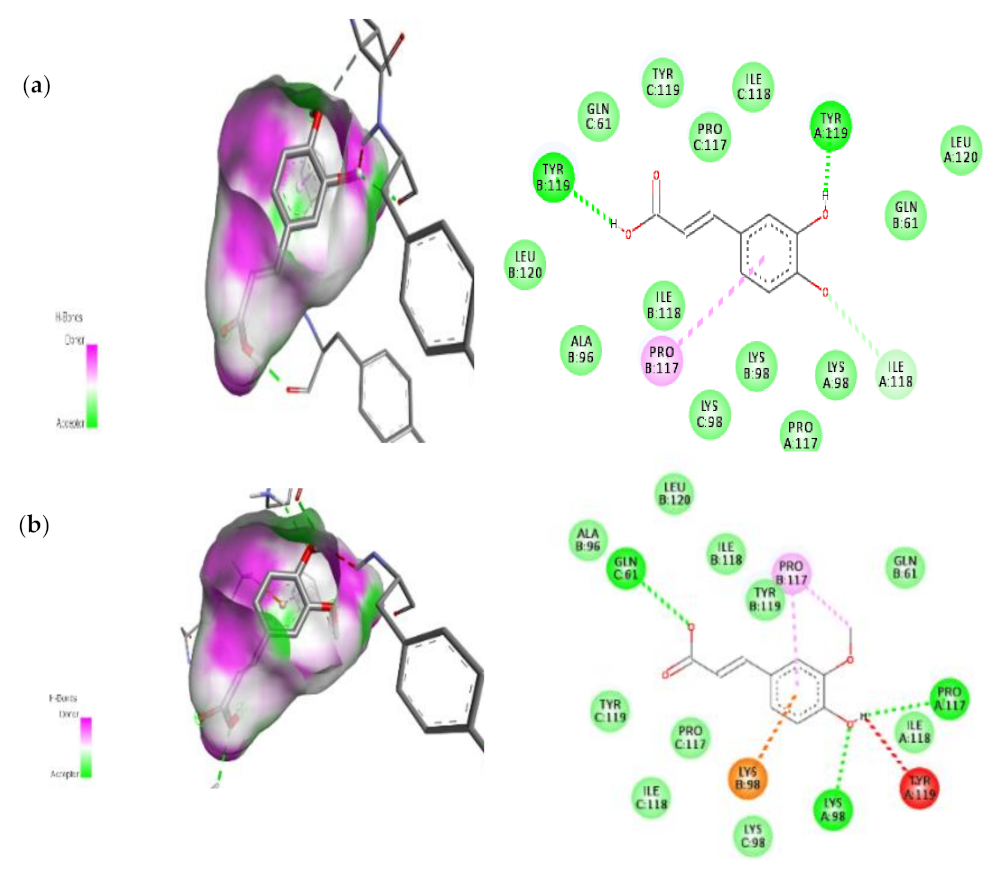
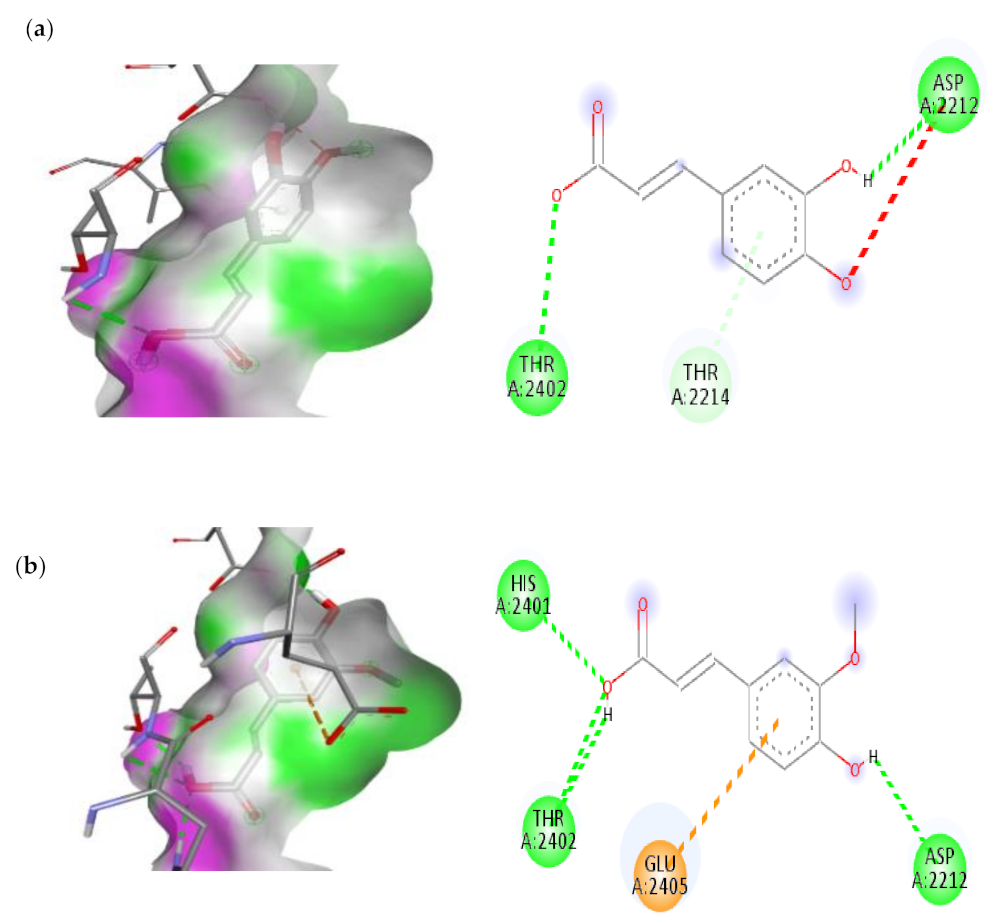
3.7.2. Molecular Docking Results of TNF Alpha and mTOR
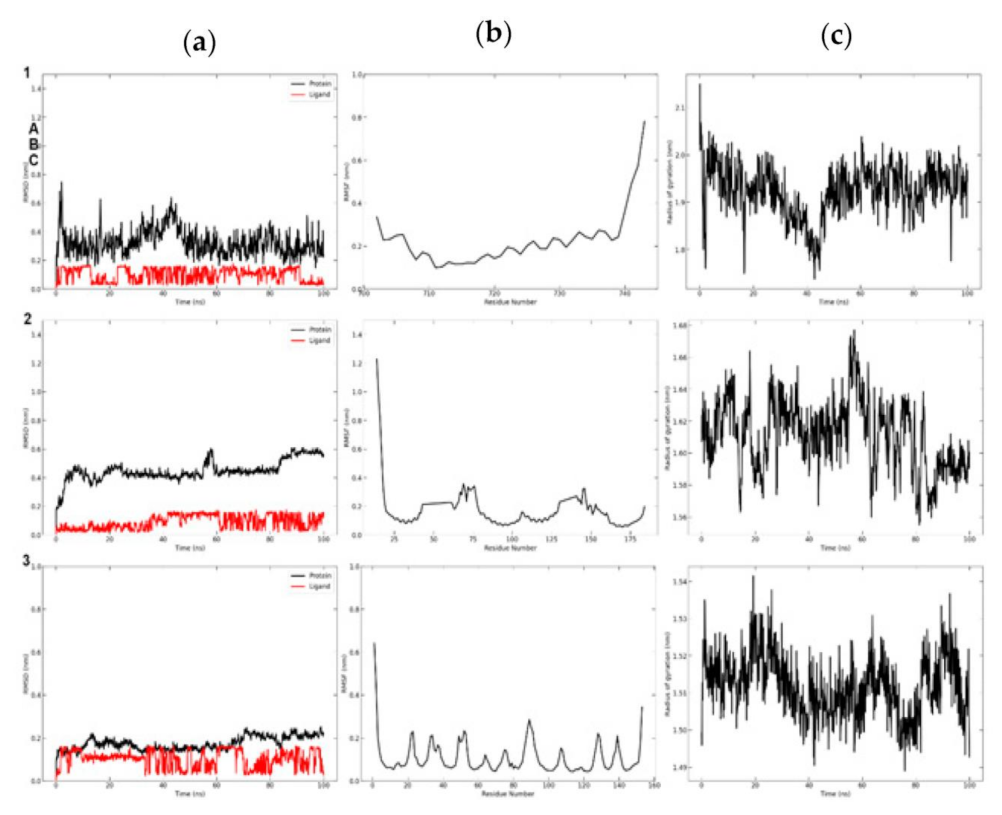
3.8. Molecular Dynamics Simulation
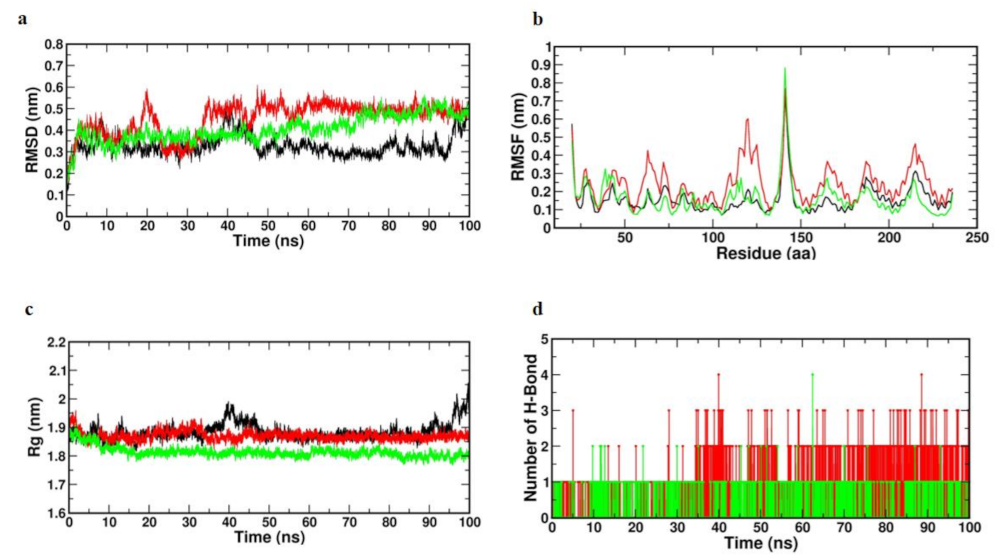
3.8.1. Trajectory Analysis of TNF-Alpha with Selected Compounds Complexes
3.8.2. Molecular Dynamic Simulation of mTOR
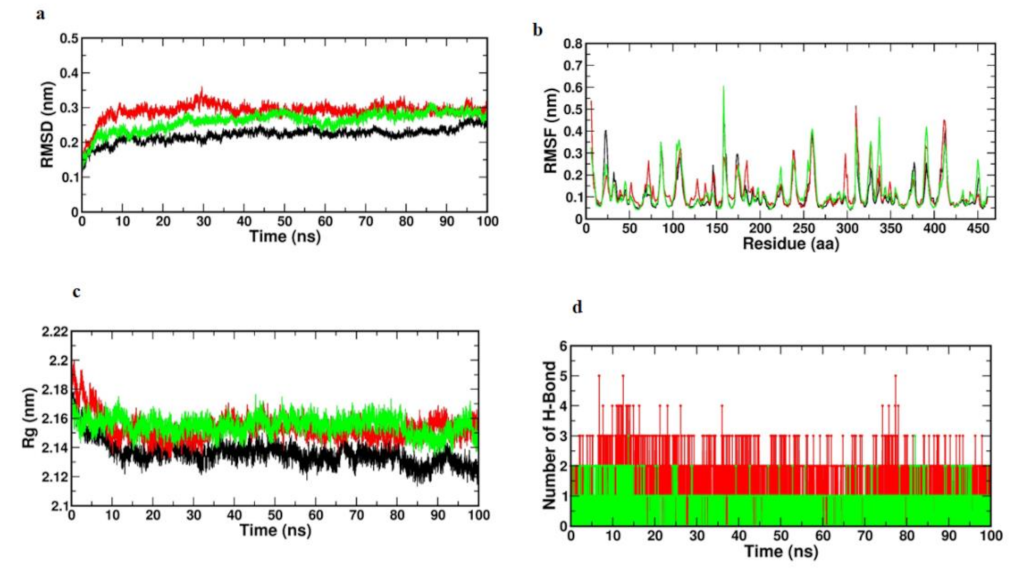
3.9. Molecular Dynamics Simulation of IL-1Beta, IL-6 and IKKβ
3.9.1. Root Mean Square Deviations RMSD
3.9.2. Root Mean Square Fluctuations RMSF
3.9.3. Radius of Gyration (Rg)
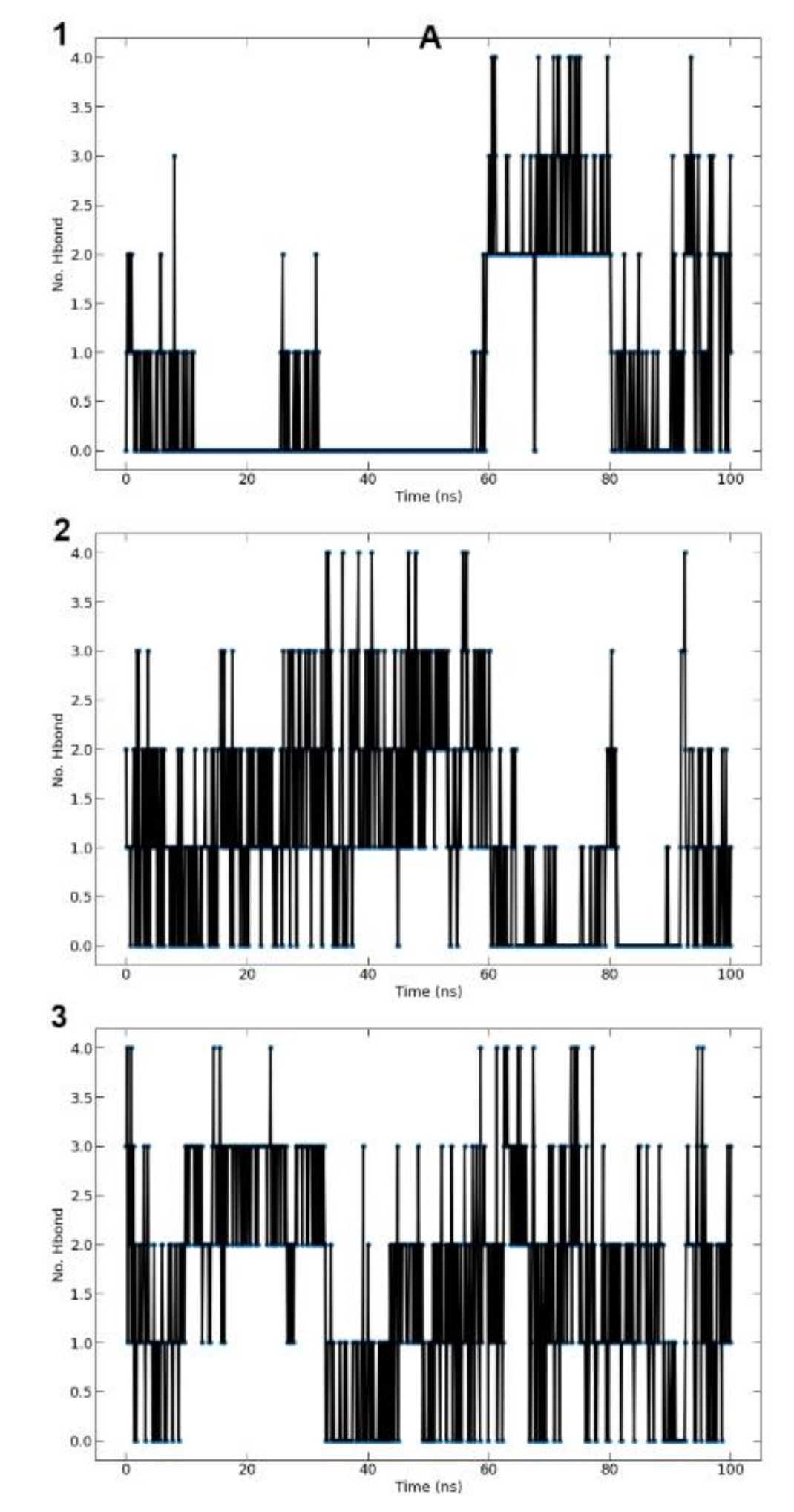
3.9.4. Hydrogen Bonds (Protein-Ligand)
3.9.5. MMPBSA Binding Energy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naser, K.A.; Gruber, A.; Thomson, G.A. The emerging pandemic of obesity and diabetes: Are we doing enough to prevent a disaster? Int. J. Clin. Pract. 2006, 60, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.; Heymsfield, S.B.; Heo, M.; Jebb, S.A.; Murgatroyd, P.R.; Sakamoto, Y. Healthy percentage body fat ranges: An approach for developing guidelines based on body mass index. Am. J. Clin. Nutr. 2000, 72, 694–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, J.M.; Cooper, M.E. Mechanisms of Diabetic Complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose tissue in obesity-related inflammation and insulin resistance: Cells, cytokines, and chemokines. ISRN Inflamm. 2013, 2013, 139239. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; McGuinness, O.P. Inflammation during obesity is not all bad: Evidence from animal and human studies. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E466–E477. [Google Scholar] [CrossRef] [Green Version]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Rehman, K.; Akash, M.S. Mechanisms of inflammatory responses and development of insulin resistance: How are they interlinked? J. Biomed. Sci. 2016, 23, 87. [Google Scholar] [CrossRef] [Green Version]
- Akash, M.S.; Rehman, K.; Chen, S. Role of inflammatory mechanisms in pathogenesis of type 2 diabetes mellitus. J. Cell. Biochem. 2013, 114, 525–531. [Google Scholar] [CrossRef]
- Sjoholm, A.; Nystrom, T. Inflammation and the etiology of type 2 diabetes. Diabetes Metab. Res. Rev. 2006, 22, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb Perspect. Biol. 2014, 6, a009191. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.W.; Egan, L.; Li, Z.W.; Greten, F.R.; Kagnoff, M.F.; Karin, M. The two faces of IKK and NF-κB inhibition: Prevention of systemic inflammation but increased local injury following intestinal ischemia-reperfusion. Nat. Med. 2003, 9, 575–581. [Google Scholar] [CrossRef]
- Maeda, S.; Chang, L.; Li, Z.W.; Luo, J.L.; Leffert, H.; Karin, M. IKKβ is required for prevention of apoptosis mediated by cell-bound but not by circulating TNFα. Immunity 2003, 19, 725–737. [Google Scholar] [CrossRef] [Green Version]
- Bing, C. Is interleukin-1β a culprit in macrophage-adipocyte crosstalk in obesity? Adipocyte 2015, 4, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Mannaa, M.; Krämer, S.; Boschmann, M.; Gollasch, M. mTOR and regulation of energy homeostasis in humans. J. Mol. Med. 2013, 91, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Kothari, V.; Galdo, J.A.; Mathews, S.T. Hypoglycemic agents and potential anti-inflammatory activity. J. Inflamm. Res. 2016, 9, 27–38. [Google Scholar] [PubMed] [Green Version]
- Owoyele, B.V.; Adebukola, O.M.; Funmilayo, A.A.; Soladoye, A.O. Anti-inflammatory activities of ethanolic extract of Carica papaya leaves. Inflammopharmacology 2008, 16, 168–173. [Google Scholar] [CrossRef]
- Pandey, S.; Cabot, P.J.; Shaw, P.N.; Hewavitharana, A.K. Anti-inflammatory and immunomodulatory properties of Carica papaya. J. Immunotoxicol. 2016, 13, 590–602. [Google Scholar] [CrossRef] [Green Version]
- Amazu, L.U.; Azikiwe, C.C.A.; Njoku, C.J.; Osuala, F.N.; Nwosu, P.J.C.; Ajugwo, A.O.; Enye, J.C. Antiinflammatory activity of the methanolic extract of the seeds of Carica papaya in experimental animals. Asian Pac. J. Trop. Med. 2010, 3, 884–886. [Google Scholar] [CrossRef] [Green Version]
- Chao, P.C.; Li, Y.; Chang, C.H.; Shieh, J.P.; Cheng, J.T.; Cheng, K.C. Investigation of insulin resistance in the popularly used four rat models of type-2 diabetes. Biomed. Pharmacother. 2018, 101, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Li, J.; Zhuang, Q.; Zhang, X.; Yuan, A.; Shen, L.; Kang, K.; Qu, B.; Tang, Y.; Pu, J.; et al. MiR-125a-5p ameliorates monocrotaline-induced pulmonary arterial hypertension by targeting the TGF-β1 and IL-6/STAT3 signaling pathways. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.T.; Zhao, X.; Yaster, M.; Tao, Y.X. Expression and distribution of mTOR, p70S6K, 4E-BP1, and their phosphorylated counterparts in rat dorsal root ganglion and spinal cord dorsal horn. Brain Res. 2010, 1336, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.A.; Abdelhalim, M.A.; Alhomida, A.S.; Al Ayed, M.S. Transient increase in IL-1β, IL-6 and TNF-α gene expression in rat liver exposed to gold nanoparticles. Genet. Mol. Res. 2013, 12, 5851–5857. [Google Scholar] [CrossRef]
- Sabouri, M.; Kordi, M.; Shabkhiz, F.; Taghibeikzadehbadr, P.; Geramian, Z.S. Moderate treadmill exercise improves spatial learning and memory deficits possibly via changing PDE-5, IL-1 β and pCREB expression. Exp. Gerontol. 2020, 139, 111056. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.; Zhao, F.; Wang, W. Use of Bao Gui capsule in treatment of a polycystic ovary syndrome rat model. Mol. Med. Rep. 2020, 21, 1461–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabe, M. Techniques Histologiques, 6th ed.; Massie e Cie: Paris, France, 1968; 1113p. [Google Scholar]
- Christy, J.; Shankari, S.; Majeed, I.; Anand, D.A. Deciphering the Synergistic Mechanism of Cortistatin towards Cancer Targets using Network Pharmacology Approach. Indian J. Pharm. Educ. Res. 2021, 55, 1017–1027. [Google Scholar] [CrossRef]
- Christy, J.; Harini; Vasudevan, S.; Lingesan, P.; Anand, D.A. Deciphering the molecular interplay between pelvic inflammatory disease (PID) and ovarian cancer (OC)—A network biology approach. Gene Rep. 2021, 25, 101405. [Google Scholar] [CrossRef]
- Schüttelkopf, A.W.; van Aalten, D.M. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef] [Green Version]
- Ost, A.; Svensson, K.; Ruishalme, I.; Brännmark, C.; Franck, N.; Krook, H.; Sandström, P.; Kjolhede, P.; Strålfors, P. Attenuated mTOR Signaling and Enhanced Autophagy in Adipocytes from Obese Patients with Type 2 Diabetes. Mol. Med. 2010, 16, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, W.; Karch, R.; Knapp, B.; Ilieva, N. Relaxation estimation of RMSD in molecular dynamics immunosimulations. Comput. Math. Methods Med. 2012, 2012, 173521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Scherer, P.E. Adipokines as novel biomarkers and regulators of the metabolic syndrome. Ann. N. Y. Acad. Sci. 2011, 1212, E1–E19. [Google Scholar] [CrossRef]
- Halberg, N.; Wernstedt-Asterholm, I.; Scherer, P.E. The adipocyte as an endocrine cell. Endocrinol. Metab. Clin. North Am. 2008, 37, 753–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Scheja, L.; Heeren, J. The endocrine function of adipose tissues in health and cardiometabolic disease. Nat. Rev. Endocrinol. 2019, 15, 507–524. [Google Scholar] [CrossRef]
- Havel, P.J. Update on adipocyte hormones: Regulation of energy balance and carbohydrate/lipid metabolism. Diabetes 2004, 53, S143–S151. [Google Scholar] [CrossRef] [Green Version]
- Lucas, S.; Verwaerde, C.; Wolowczuk, I. Is the adipose tissue the key road to inflammation? Immunol. Immunogenet. Insights 2009, 1, III-S2145. [Google Scholar] [CrossRef] [Green Version]
- Tilg, H.; Moschen, A.R. Inflammatory mechanisms in the regulation of insulin resistance. Mol. Med. 2008, 14, 222–231. [Google Scholar] [CrossRef]
- Jayaraman, S.; Devarajan, N.; Rajagopal, P.; Babu, S.; Ganesan, S.K.; Veeraraghavan, V.P.; Palanisamy, C.P.; Cui, B.; Periyasamy, V.; Chandrasekar, K. β-Sitosterol Circumvents Obesity Induced Inflammation and Insulin Resistance by Down-Regulating IKKβ/NF-κB and JNK Signaling Pathway in Adipocytes of Type 2 Diabetic Rats. Molecules 2021, 26, 2101. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [Green Version]
- Jaganathan, R.; Ravindran, R.; Dhanasekaran, S. Emerging role of adipocytokines in type 2 diabetes as mediators of insulin resistance and cardiovascular disease. Can. J. Diabetes 2018, 42, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H. Adipocytokines in obesity and metabolic disease. J. Endocrinol. 2014, 220, T47–T59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Acta 2013, 17, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Ziemke, F.; Mantzoros, C.S. Adiponectin in insulin resistance: Lessons from translational research. Am. J. Clin. Nutr. 2010, 91, 258S–261S. [Google Scholar] [CrossRef] [Green Version]
- Aleidi, S.; Issa, A.; Bustanji, H.; Khalil, M.; Bustanji, Y. Adiponectin serum levels correlate with insulin resistance in type 2 diabetic patients. Saudi Pharm. J. 2015, 23, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Reilly, M.P.; Iqbal, N.; Schutta, M.; Wolfe, M.L.; Scally, M.; Localio, A.R.; Rader, D.J.; Kimmel, S.E. Plasma leptin levels are associated with coronary atherosclerosis in type 2 diabetes. J. Clin. Endocrinol. Metab. 2004, 89, 3872–3878. [Google Scholar] [CrossRef]
- Heidemann, C.; Sun, Q.; van Dam, R.M.; Meigs, J.B.; Zhang, C.; Tworoger, S.S.; Mantzoros, C.S.; Hu, F.B. Total and high-molecular-weight adiponectin and resistin in relation to the risk for type 2 diabetes in women. Ann. Intern. Med. 2008, 149, 307–316. [Google Scholar] [CrossRef]
- Ghanbari, M.; Lamuki, M.S.; Habibi, E.; Sadeghimahalli, F. Artemisia annua L. Extracts Improved Insulin Resistance via Changing Adiponectin, Leptin and Resistin Production in HFD/STZ Diabetic Mice. J. Pharmacopunct. 2022, 25, 130–137. [Google Scholar] [CrossRef]
- Ansari, S.; Bari, A.; Ullah, R.; Mathanmohun, M.; Veeraraghavan, V.P.; Sun, Z. Gold nanoparticles synthesized with Smilax glabra rhizome modulates the anti-obesity parameters in high-fat diet and streptozotocin induced obese diabetes rat model. J. Photochem. Photobiol. B Biol. 2019, 201, 111643. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.; Polotsky, V.Y. Leptin and Leptin Resistance in the Pathogenesis of Obstructive Sleep Apnea: A Possible Link to Oxidative Stress and Cardiovascular Complications. Oxid. Med. Cell. Longev. 2018, 2018, 5137947. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, Q.; Cai, D.; Guo, H.; Fang, J.; Cui, H.; Gou, L.; Deng, J.; Wang, Z.; Zuo, Z. Resistin, a Novel Host Defense Peptide of Innate Immunity. Front. Immunol. 2021, 12, 699807. [Google Scholar] [CrossRef] [PubMed]
- Lay, S.L.; Simard, G.; Martinez, M.C. Oxidative stress and metabolic pathologies: From an adipocentric point of view. Oxid. Med. Cell. Longev. 2014, 2014, 908539. [Google Scholar] [CrossRef] [Green Version]
- Othman, M.S.; Khaled, A.M.; Al-Bagawi, A.H.; Fareid, M.A.; Ghany, R.A.; Habotta, O.A.; Moneim, A.E.A. Hepatorenal protective efficacy of flavonoids from Ocimum basilicum extract in diabetic albino rats: A focus on hypoglycemic, antioxidant, anti-inflammatory and anti-apoptotic activities. Biomed. Pharmacother. 2021, 144, 112287. [Google Scholar] [CrossRef]
- Sadek, K.M. Antioxidant and immunostimulant effect of Carica papaya linn. Aqueous extract in acrylamide intoxicated rats. Acta Inform. Med. 2012, 20, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Nain, P.; Saini, V.; Sharma, S.; Nain, J. Antidiabetic and antioxidant potential of Emblica officinalis Gaertn. leaves extract in streptozotocin-induced type-2 diabetes mellitus (T2DM) rats. J. Ethnopharmacol. 2012, 142, 65–71. [Google Scholar] [CrossRef]
- Gouranton, E.; Thabuis, C.; Riollet, C.; Malezet-Desmoulins, C.; El Yazidi, C.; Amiot, M.J.; Borel, P.; Landrier, J.F. Lycopene inhibits proinflammatory cytokine and chemokine expression in adipose tissue. J. Nutr. Biochem. 2011, 22, 642–648. [Google Scholar] [CrossRef]
- Kowalska, K.; Olejnik, A.; Zielińska-Wasielica, J.; Olkowicz, M. Raspberry (Rubus idaeus L.) fruit extract decreases oxidation markers, improves lipid metabolism and reduces adipose tissue inflammation in hypertrophied 3T3-L1 adipocytes. J. Funct. Foods 2019, 62, 103568. [Google Scholar] [CrossRef]
- Shokouh, P.; Jeppesen, P.B.; Christiansen, C.B.; Mellbye, F.B.; Hermansen, K.; Gregersen, S. Efficacy of arabica versus robusta coffee in improving weight, insulin resistance, and liver steatosis in a rat model of type-2 diabetes. Nutrients 2019, 11, 2074. [Google Scholar] [CrossRef] [Green Version]
- McIver, C.M.; Wycherley, T.P.; Clifton, P.M. MTOR signaling and ubiquitin-proteosome gene expression in the preservation of fat free mass following high protein, calorie restricted weight loss. Nutr. Metab. 2012, 9, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Wang, L.; Chen, Z.; Hou, D.; Xue, Y.; Diao, X.; Shen, Q. Foxtail millet improves blood glucose metabolism in diabetic rats through pi3k/akt and nf-κb signaling pathways mediated by gut microbiota. Nutrients 2021, 13, 1837. [Google Scholar] [CrossRef] [PubMed]
- Shabani, M.; Sadeghi, A.; Hosseini, H.; Teimouri, M.; Khorzoughi, R.B.; Pasalar, P.; Meshkani, R. Resveratrol alleviates obesity-induced skeletal muscle inflammation via decreasing M1 macrophage polarization and increasing the regulatory T cell population. Sci. Rep. 2020, 10, 3791. [Google Scholar] [CrossRef] [Green Version]
- Moruzzi, M.; Klöting, N.; Blüher, M.; Martinelli, I.; Tayebati, S.K.; Gabrielli, M.G.; Roy, P.; Micioni Di Bonaventura, M.V.; Cifani, C.; Lupidi, G.; et al. Tart Cherry Juice and Seeds Affect Pro-Inflammatory Markers in Visceral Adipose Tissue of High-Fat Diet Obese Rats. Molecules 2021, 26, 1403. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Shankar, E.; Fu, P.; MacLennan, G.T.; Gupta, S. Suppression of NF-κB and NF-κB-Regulated Gene Expression by Apigenin through IκBα and IKK Pathway in TRAMP Mice. PLoS ONE 2015, 10, e0138710. [Google Scholar] [CrossRef] [Green Version]
- Mopuri, R.; Ganjayi, M.; Banavathy, K.S.; Parim, B.N.; Meriga, B. Evaluation of anti-obesity activities of ethanolic extract of Terminalia paniculate bark on high fat diet-induced obese rats. BMC Complement. Altern. Med. 2015, 15, 76. [Google Scholar] [CrossRef] [Green Version]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S1. No | Name of Gene | Gene Specific Primers | Reference |
|---|---|---|---|
| 1 | Beta actin | Sense primer: 5′-CGC GAG TAC AAC CTT CTT GC-3′ Anti-sense primer: 5′-CGT CAT CCA TGG CGA ACT GG-3′ | [23] |
| 2 | mTOR | Sense primer: 5′-TTG AGG TTG CTA TGA CCA GAG AGA A-3′ Anti-sense primer: 5′-TTA CCA GAA AGG ACA CCA GCC AAT G-3′ | [24] |
| 3 | TNF-α | Sense primer: 5′-ACT GAA CTT CGG GGT GAT TG-3′ Anti-sense primer: 5′-GCT TGG TGG TTT GCT ACG AC-3′ | [25] |
| 4 | IL-1β | Sense primer: 5′-TAC CTA TGT CTG GCC CGT GGA G-3′ Anti-sense primer: 5′-ATC ATC CCA CGA GTC ACA CAG G-3′ | [26] |
| 5 | IL-6 | Sense primer: 5′-TTC TCT CCG CAA GAG ACT TCC-3′ Anti-sense primer: 5′-TTC TGA CAG TGC ATC ATC GCT-3′ | [23] |
| 6 | IKKβ | Sense primer: 5′-GCA CCC TGG CCT TTG AAT G-3′ Anti-sense primer: 5′-TCC GTT CAA GTC CTC GCT AAC A-3′ | [27] |
| Sl. No | Compound Name |
|---|---|
| i. | Transferulic acid |
| ii. | Quercetin |
| iii. | Rutin |
| iv. | Chlorogenic acid |
| v. | Kaempferol |
| vi. | Protocatechuic acid |
| vii. | Caffeic acid |
| viii. | p-coumaric acid |
| Sl. No | Compound Name | Binding Energy kcal/mol | Interacting Residues |
|---|---|---|---|
| IL-1β | |||
| 1. | Quercetin | −6.3 | PRO-78 (H-bond)GLU-25(Pi-Anion) |
| 2. | Kaempferol | −6.3 | TYR-24(H-bond) GLU-25(H-bond) |
| IL-6 | |||
| 1. | Quercetin | −6.1 | ARG-168(H-bond) ARG-40(Pi-Sigma) SER-37(Pi-Alkyl) LYS-171(Pi-Cation) LEU-33(Pi-Alkyl) |
| 2. | Kaempferol | −5.7 | LEU-33(H-bond) GLN-175(H-bond) SER-37(Pi-Sigma) ARG-30(Pi-Alkyl) LYS-171(Pi-Alkyl) |
| IKKβ | |||
| 1. | Quercetin | −5 | HIS-713(H-bond) THR-717(H-bond) LEU-719(Pi-Amide) CYS-716(Pi-Alkyl) ILE-723(Pi-Alkyl) |
| 2. | Kaempferol | −5 | THR-717(H-bond) LEU-719(Pi-Amide) |
| Sl No. | Compound Name | Binding Energy kcal/mol | Interacting Residues |
|---|---|---|---|
| TNF-α | |||
| 1. | Caffeic acid | −6.7 | TYR-119(H-Bond) PRO-117(Pi-alkyl) |
| 2. | Transferulic acid | −6.6 | GLN-61(H-Bond) LYS-98((Pi-alkyl)) TYR-119(H-Bond) LYS-98(Pi-cation) |
| mTOR | |||
| 1. | Caffeic acid | −6.5 | ASP-2212(H-Bond) THR-2402(H-Bond) |
| 2. | Transferulic acid | −6.4 | ASP-2212(H-Bond) HIS-2401(H-Bond) THR-2402(H-Bond) GLU-2405(Pi-Anion) |
| Compounds Name | VdW Energy (kJ mol−1) | Electrostatic Energy (kJ mol−1) | Polar Solvation Energy (kJ mol−1) | SASA Energy (kJ mol−1) | ∆G |
|---|---|---|---|---|---|
| TNF-α | |||||
| Caffeic acid | 127.312 ± 6.976 | −3.326 ± 2.756 | 57.699 ± 8.530 | −12.101 ± 0.693 | −85.040 ± 9.971 |
| Trans-Ferulic Acid | −140.334 ± 8.549 | −2.877 ± 2.564 | 90.000 ± 16.505 | −13.070 ± 0.740 | −66.281 ± 16.686 |
| mTOR | |||||
| Caffeic acid | −133.129 ± 8.888 | −4.746 ± 2.692 | 49.135 ± 8.077 | −11.507 ± 0.635 | −100.248 ± 9.760 |
| Trans-Ferulic Acid | −135.313 ± 9.188 | −5.112 ± 4.418 | 59.421 ± 22.356 | −12.723 ± 0.635 | −93.726 ± 24.063 |
| Complex | Van Der Waal Energy | Electrostatic Energy | Polar Solvation Energy | SASA Energy | |
|---|---|---|---|---|---|
| 3BRV | −70.444 ± 38.771 | −47.325 ± 30.022 | −60.150 ± 68.143 | 43.763 ± 40.074 | −6.732 ± 3.409 |
| 4NI9 | −66.310 ± 42.238 | −74.532 ± 31.307 | −67.017 ± 70.124 | 84.696 ± 49.905 | −9.457 ± 3.103 |
| 9ILB | −78.544 ± 25.532 | −71.504 ± 25.995 | −96.621 ± 47.023 | 98.843 ± 48.350 | −9.263 ± 3.531 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebecca Roy, J.; Janaki, C.S.; Jayaraman, S.; Periyasamy, V.; Balaji, T.; Vijayamalathi, M.; Veeraraghavan, V.P.; Krishnamoorthy, K.; Prasad, M. Carica Papaya Reduces High Fat Diet and Streptozotocin-Induced Development of Inflammation in Adipocyte via IL-1β/IL-6/TNF-α Mediated Signaling Mechanisms in Type-2 Diabetic Rats. Curr. Issues Mol. Biol. 2023, 45, 852-884. https://doi.org/10.3390/cimb45020056
Rebecca Roy J, Janaki CS, Jayaraman S, Periyasamy V, Balaji T, Vijayamalathi M, Veeraraghavan VP, Krishnamoorthy K, Prasad M. Carica Papaya Reduces High Fat Diet and Streptozotocin-Induced Development of Inflammation in Adipocyte via IL-1β/IL-6/TNF-α Mediated Signaling Mechanisms in Type-2 Diabetic Rats. Current Issues in Molecular Biology. 2023; 45(2):852-884. https://doi.org/10.3390/cimb45020056
Chicago/Turabian StyleRebecca Roy, Jeane, Coimbatore Sadagopan Janaki, Selvaraj Jayaraman, Vijayalakshmi Periyasamy, Thotakura Balaji, Madhavan Vijayamalathi, Vishnu Priya Veeraraghavan, Kalaiselvi Krishnamoorthy, and Monisha Prasad. 2023. "Carica Papaya Reduces High Fat Diet and Streptozotocin-Induced Development of Inflammation in Adipocyte via IL-1β/IL-6/TNF-α Mediated Signaling Mechanisms in Type-2 Diabetic Rats" Current Issues in Molecular Biology 45, no. 2: 852-884. https://doi.org/10.3390/cimb45020056