
Stability of Dengue 2 Nonstructural Glycoprotein 1 (NS1) Is Affected by the Nature of Basic Residue at Position NS1-324

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Antibodies
2.2. Vector Plasmids Expressing Recombinant DENV-2 NS1 Proteins
2.3. Immunoblot Assay
2.4. Proteasome Inhibition Assay
2.5. RT-qPCR
2.6. Measurement of ROS
2.7. Statistical Analysis
3. Results
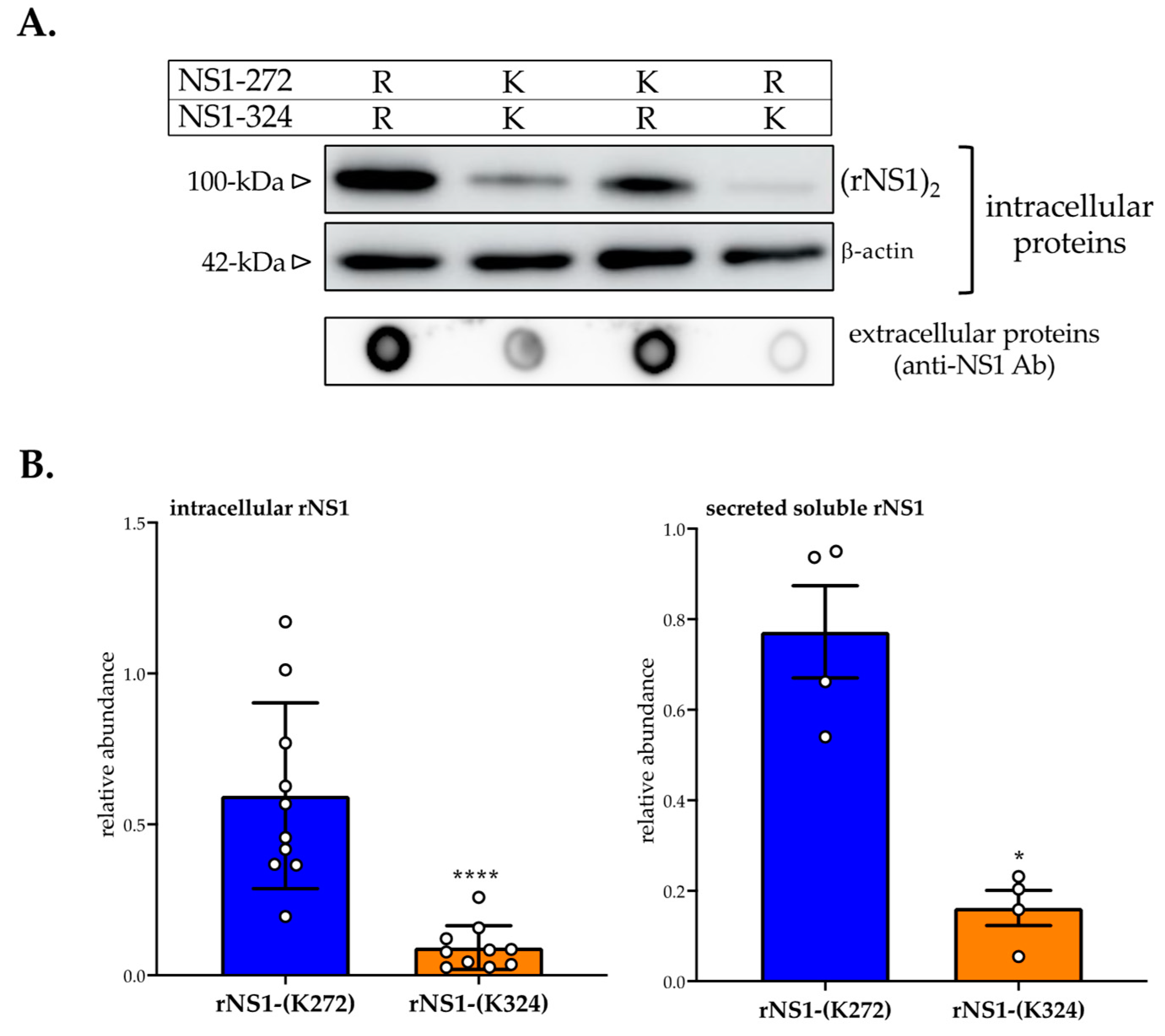
3.1. Effect of Amino-Acid Substitutions R272K and R324K on DES-14 rNS1 Protein
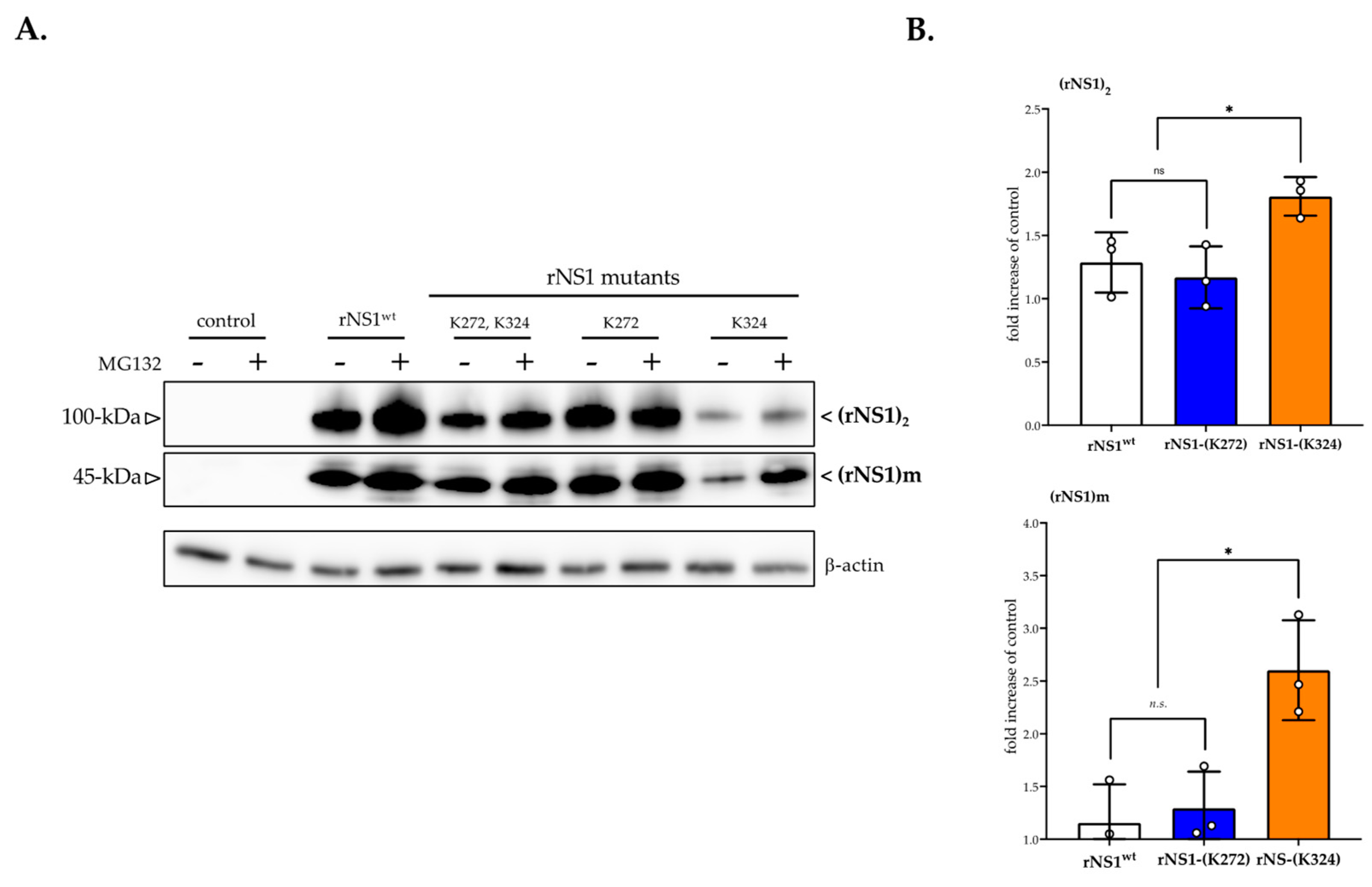
3.2. Effect of Proteasome Inhibition on rNS1 Protein Expression in Huh7 Cells
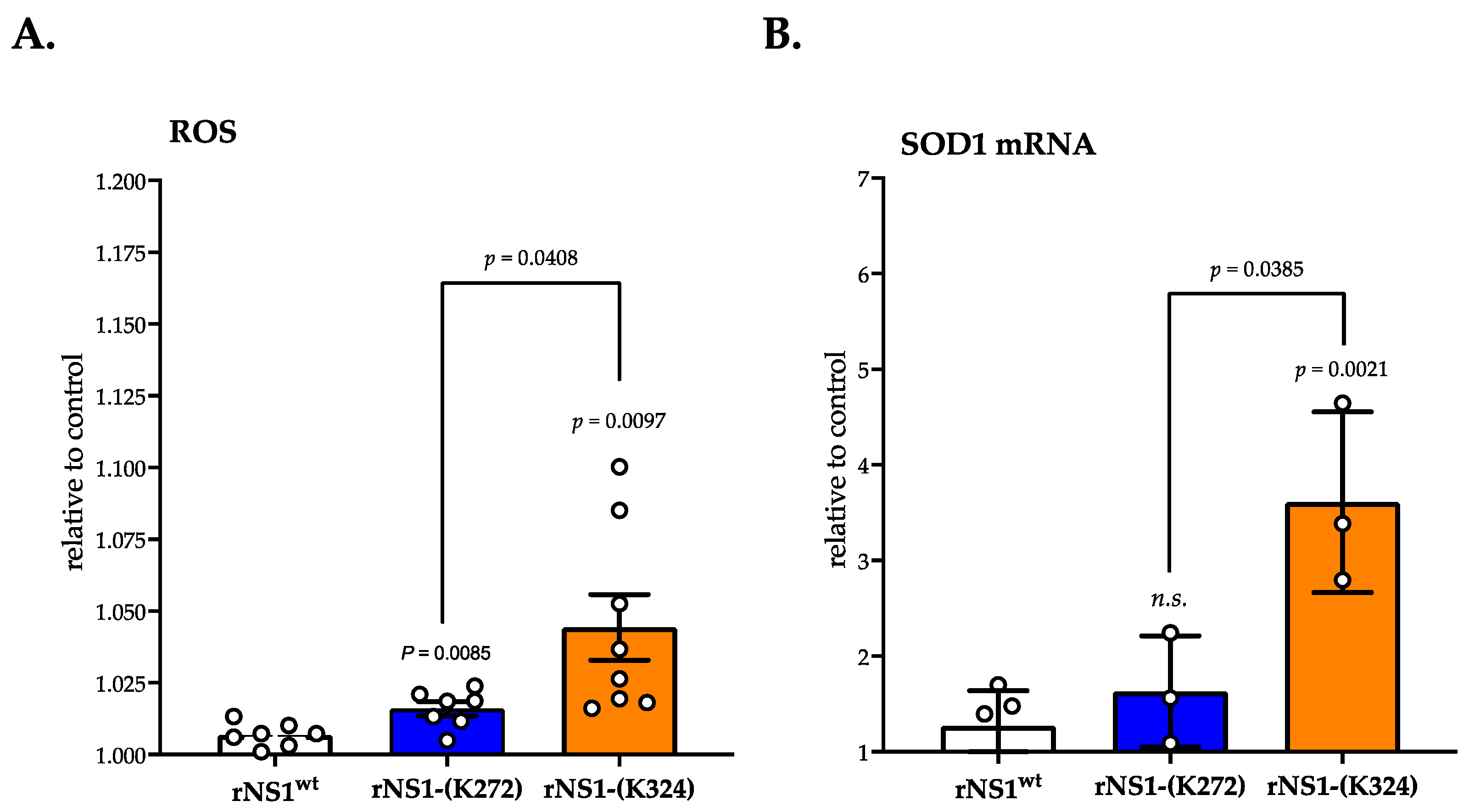
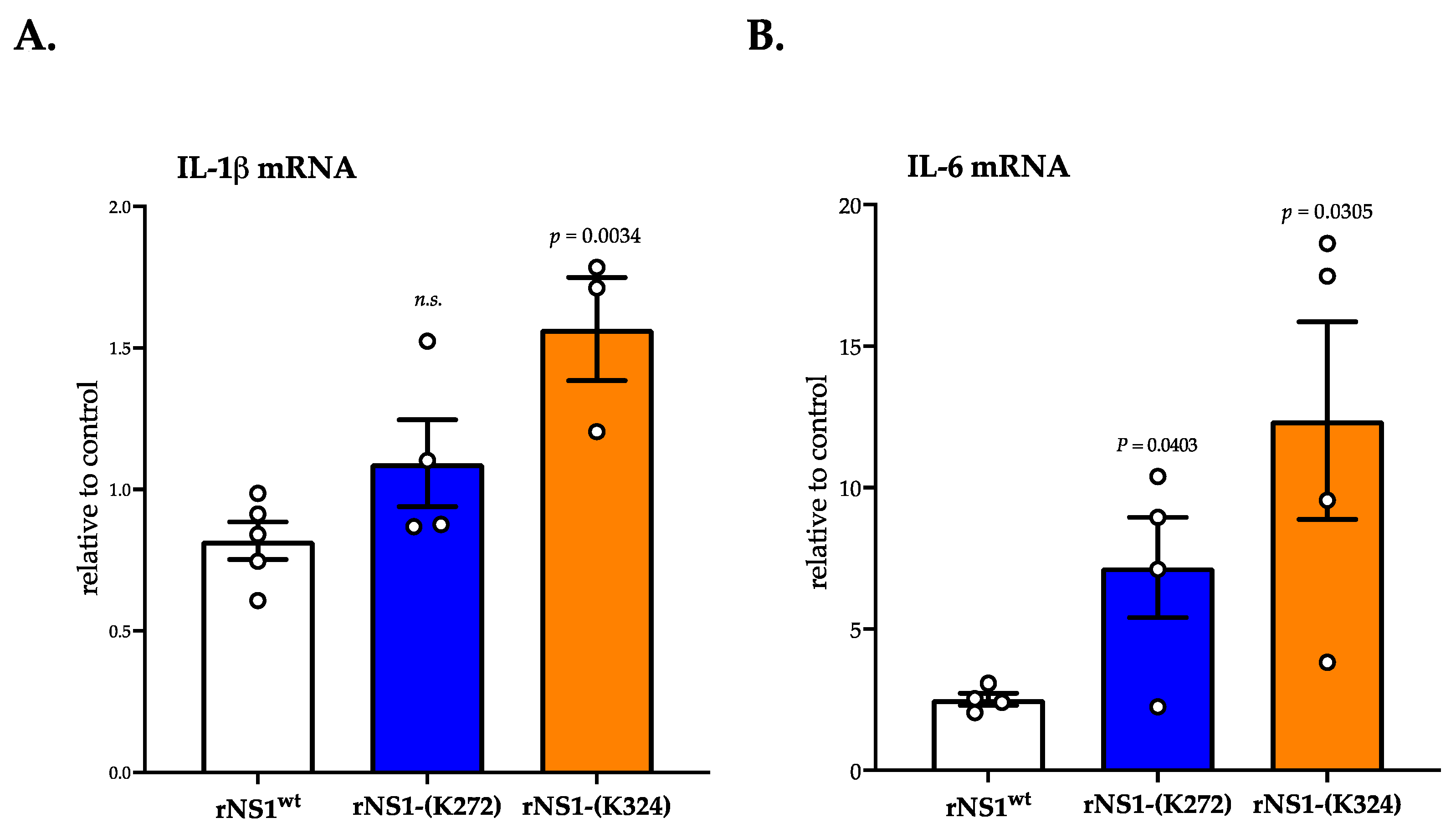
3.3. Ability of rNS1-(Lys324) Mutant to Trigger Oxidative Stress and Pro-Inflammatory Cytokine Transcription Activation
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ab | Antibody |
| DCFH-DA | Dichlorodihydrofluorescein diacetate |
| DENV | Dengue virus |
| DES-14 | DENV-2 strain D2-K2_RIJ059/Dar es Salaam 2014 |
| DMEM | Dulbecco’s Modified Eagle Medium |
| ECL | Enhanced chemiluminescent |
| HRP | Horseradish-peroxidase |
| Il-1β | Interleukine 1β |
| IL-6 | Interleukine 6 |
| kDa | kiloDalton |
| mAb | Monoclonal antibody |
| mRNA | Messager Ribonucleic acid |
| NS1 | Non-structural protein 1 |
| pAb | Polyclonal antibody |
| rNS1 | Recombinant non-structural protein 1 |
| ROS | Reactive Oxidative Species |
| RT-qPCR | Reverse Transcription- quantitative Polymerase Chain Reaction |
| RUN-18 | DENV-2 strain RUN-18 (or RUJul, Genbank accession number MN272404) |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SOD1 | Superoxide dismutase 1 |
| SWIO | Southwestern Indian Ocean |
| UPS | Ubiquitin-proteasome system |
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Special Programme for Research and Training in Tropical Diseases and World Health Organization, Eds. Dengue: Guidelines for Diagnosis, Treatment, Prevention, and Control, New ed; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Gubler, D.J. Dengue/Dengue Haemorrhagic Fever: History and Current Status. In Novartis Foundation Symposia; Bock, G., Goode, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2008; pp. 3–22. [Google Scholar] [CrossRef]
- Guy, B.; Jackson, N. Dengue vaccine: Hypotheses to understand CYD-TDV-induced protection. Nat. Rev. Microbiol. 2016, 14, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Rauscher, M.; Kudela, M.; Pang, H. Clinical Safety Experience of TAK-003 for Dengue Fever: A New Tetravalent Live Attenuated Vaccine Candidate. Clin. Infect. Dis. 2023, 76, e1350–e1359. [Google Scholar] [CrossRef] [PubMed]
- Pascalis, H.; Turpin, J.; Roche, M.; Krejbich, P.; Gadea, G.; Nten, C.A.; Desprès, P.; Mavingui, P. The epidemic of Dengue virus type-2 Cosmopolitan genotype on Reunion Island relates to its active circulation in the Southwestern Indian Ocean neighboring islands. Heliyon 2019, 5, e01455. [Google Scholar] [CrossRef] [PubMed]
- Hafsia, S.; Haramboure, M.; Wilkinson, D.A.; Baldet, T.; Yemadje-Menudier, L.; Vincent, M.; Tran, A.; Atyame, C.; Mavingui, P. Overview of dengue outbreaks in the southwestern Indian Ocean and analysis of factors involved in the shift toward endemicity in Reunion Island: A systematic review. PLoS Negl. Trop. Dis. 2022, 16, e0010547. [Google Scholar] [CrossRef]
- Ogire, E.; Diaz, O.; Vidalain, P.-O.; Lotteau, V.; Desprès, P.; Roche, M. Instability of the NS1 Glycoprotein from La Reunion 2018 Dengue 2 Virus (Cosmopolitan-1 Genotype) in Huh7 Cells Is Due to Lysine Residues on Positions 272 and 324. IJMS 2021, 22, 1951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Rangarajan, E.S.; Izard, T.; Farzan, M.; Choe, H. The D614G mutation in the SARS-CoV-2 spike protein reduces S1 shedding and increases infectivity. Microbiology 2022. preprint. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; VanLandingham, D.L.; McGee, C.E.; Higgs, S. A Single Mutation in Chikungunya Virus Affects Vector Specificity and Epidemic Potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef]
- Hawman, D.W.; Carpentier, K.S.; Fox, J.M.; May, N.A.; Sanders, W.; Montgomery, S.A.; Moorman, N.J.; Diamond, M.S.; Morrison, T.E. Mutations in the E2 Glycoprotein and the 3′ Untranslated Region Enhance Chikungunya Virus Virulence in Mice. J. Virol. 2017, 91, e00816-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sannigrahi, A.; Maity, P.; Karmakar, S.; Chattopadhyay, K. Interaction of KMP-11 with Phospholipid Membranes and Its Implications in Leishmaniasis: Effects of Single Tryptophan Mutations and Cholesterol. J. Phys. Chem. B 2017, 121, 1824–1834. [Google Scholar] [CrossRef] [PubMed]
- Sannigrahi, A.; Chall, S.; Jawed, J.J.; Kundu, A.; Majumdar, S.; Chattopadhyay, K. Nanoparticle Induced Conformational Switch Between α-Helix and β-Sheet Attenuates Immunogenic Response of MPT63. Langmuir 2018, 34, 8807–8817. [Google Scholar] [CrossRef] [PubMed]
- Sannigrahi, A.; Nandi, I.; Chall, S.; Jawed, J.J.; Halder, A.; Majumdar, S.; Karmakar, S.; Chattopadhyay, K. Conformational Switch Driven Membrane Pore Formation by Mycobacterium Secretory Protein MPT63 Induces Macrophage Cell Death. ACS Chem. Biol. 2019, 14, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Figueira-Mansur, J.; Aguilera, E.A.; Stoque, R.M.; Ventura, G.T.; Mohana-Borges, R. Mutations in the dimer interfaces of the dengue virus capsid protein affect structural stability and impair RNA-capsid interaction. Sci. Rep. 2019, 9, 2829. [Google Scholar] [CrossRef] [PubMed]
- Biering, S.B.; Akey, D.L.; Wong, M.P.; Brown, W.C.; Lo, N.T.N.; Puerta-Guardo, H.; de Sousa, F.T.G.; Wang, C.; Konwerski, J.R.; Espinosa, D.A.; et al. Structural basis for antibody inhibition of flavivirus NS1–triggered endothelial dysfunction. Science 2021, 371, 194–200. [Google Scholar] [CrossRef]
- Vairo, F.; Mboera, L.E.; De Nardo, P.; Oriyo, N.M.; Meschi, S.; Rumisha, S.F.; Colavita, F.; Mhina, A.; Carletti, F.; Mwakapeje, E.; et al. Clinical, Virologic, and Epidemiologic Characteristics of Dengue Outbreak, Dar es Salaam, Tanzania, 2014. Emerg. Infect. Dis. 2016, 22, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, M.I.G.; Vargas-Cuartas, O.; Gallego-Gomez, J.C.; Shi, P.-Y.; Padilla-Sanabria, L.; Castaño-Osorio, J.C.; Rajsbaum, R. K48-linked polyubiquitination of dengue virus NS1 protein inhibits its interaction with the viral partner NS4B. Virus Res. 2018, 246, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.E.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and Three-Dimensional Architecture of the Dengue Virus Replication and Assembly Sites. Cell Host Microbe 2009, 4, 365–375. [Google Scholar] [CrossRef]
- Mackenzie, J.; Jones, M.; Young, P.R. Immunolocalization of the Dengue Virus Nonstructural Glycoprotein NS1 Suggests a Role in Viral RNA Replication. Virology 1996, 220, 232–240. [Google Scholar] [CrossRef]
- Scaturro, P.; Cortese, M.; Chatel-Chaix, L.; Fischl, W.; Bartenschlager, R. Dengue Virus Non-structural Protein 1 Modulates Infectious Particle Production via Interaction with the Structural Proteins. PLoS Pathog. 2015, 11, e1005277. [Google Scholar] [CrossRef] [PubMed]
- Flamand, M.; Megret, F.; Mathieu, M.; Lepault, J.; Rey, F.A.; Deubel, V. Dengue Virus Type 1 Nonstructural Glycoprotein NS1 Is Secreted from Mammalian Cells as a Soluble Hexamer in a Glycosylation-Dependent Fashion. J. Virol. 1999, 73, 6104–6110. [Google Scholar] [CrossRef] [PubMed]
- Gutsche, I.; Coulibaly, F.; Voss, J.E.; Salmon, J.; D’Alayer, J.; Ermonval, M.; Larquet, E.; Charneau, P.; Krey, T.; Mégret, F.; et al. Secreted dengue virus nonstructural protein NS1 is an atypical barrel-shaped high-density lipoprotein. Proc. Natl. Acad. Sci. USA 2011, 108, 8003–8008. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.R.; Puerta-Guardo, H.; Killingbeck, S.S.; Glasner, D.R.; Hopkins, K.; Harris, E. Dengue virus NS1 triggers endothelial permeability and vascular leak that is prevented by NS1 vaccination. Sci. Transl. Med. 2015, 7, 304ra141. [Google Scholar] [CrossRef] [PubMed]
- Glasner, D.R.; Puerta-Guardo, H.; Beatty, P.R.; Harris, E. The Good, the Bad, and the Shocking: The Multiple Roles of Dengue Virus Nonstructural Protein 1 in Protection and Pathogenesis. Annu. Rev. Virol. 2018, 5, 227–253. [Google Scholar] [CrossRef] [PubMed]
- Puerta-Guardo, H.; Biering, S.B.; de Sousa, F.T.G.; Shu, J.; Glasner, D.R.; Li, J.; Blanc, S.F.; Beatty, P.R.; Harris, E. Flavivirus NS1 Triggers Tissue-Specific Disassembly of Intercellular Junctions Leading to Barrier Dysfunction and Vascular Leak in a GSK-3β-Dependent Manner. Pathogens 2022, 11, 615. [Google Scholar] [CrossRef] [PubMed]
- Flick, K.; Kaiser, P. Protein degradation and the stress response. Semin. Cell Dev. Biol. 2012, 23, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K. Integration of ER stress, oxidative stress and the inflammatory response in health and disease. Int. J. Clin. Exp. Med. 2010, 3, 33–40. [Google Scholar]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Bulua, A.C.; Simon, A.; Maddipati, R.; Pelletier, M.; Park, H.; Kim, K.-Y.; Sack, M.N.; Kastner, D.L.; Siegel, R.M. Mitochondrial reactive oxygen species promote production of proinflammatory cytokines and are elevated in TNFR1-associated periodic syndrome (TRAPS). J. Exp. Med. 2011, 208, 519–533. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Rape, M. The Ubiquitin Code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef]
- Geiss-Friedlander, R.; Melchior, F. Concepts in sumoylation: A decade on. Nat Rev Mol Cell Biol 2007, 8, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, C.; Weinert, B.T.; Nishida, Y.; Verdin, E.; Mann, M. The growing landscape of lysine acetylation links metabolism and cell signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 536–550. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-I.; Tseng, C.-H.; Yu, C.-Y.; Lai, M.M.C. SUMO Modification Stabilizes Dengue Virus Nonstructural Protein 5 To Support Virus Replication. J. Virol. 2016, 90, 4308–4319. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogire, E.; El-Kalamouni, C.; Desprès, P.; Roche, M. Stability of Dengue 2 Nonstructural Glycoprotein 1 (NS1) Is Affected by the Nature of Basic Residue at Position NS1-324. Curr. Issues Mol. Biol. 2023, 45, 1644-1654. https://doi.org/10.3390/cimb45020106
Ogire E, El-Kalamouni C, Desprès P, Roche M. Stability of Dengue 2 Nonstructural Glycoprotein 1 (NS1) Is Affected by the Nature of Basic Residue at Position NS1-324. Current Issues in Molecular Biology. 2023; 45(2):1644-1654. https://doi.org/10.3390/cimb45020106
Chicago/Turabian StyleOgire, Eva, Chaker El-Kalamouni, Philippe Desprès, and Marjolaine Roche. 2023. "Stability of Dengue 2 Nonstructural Glycoprotein 1 (NS1) Is Affected by the Nature of Basic Residue at Position NS1-324" Current Issues in Molecular Biology 45, no. 2: 1644-1654. https://doi.org/10.3390/cimb45020106