
Water Extract of Portulaca Oleracea Inhibits PEDV Infection-Induced Pyrolysis by Caspase-1/GSDMD

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Virus
2.2. CCK-8 Assay
2.3. Measurement of LDH Release and ATP Production
2.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.5. Immunofluorescence
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
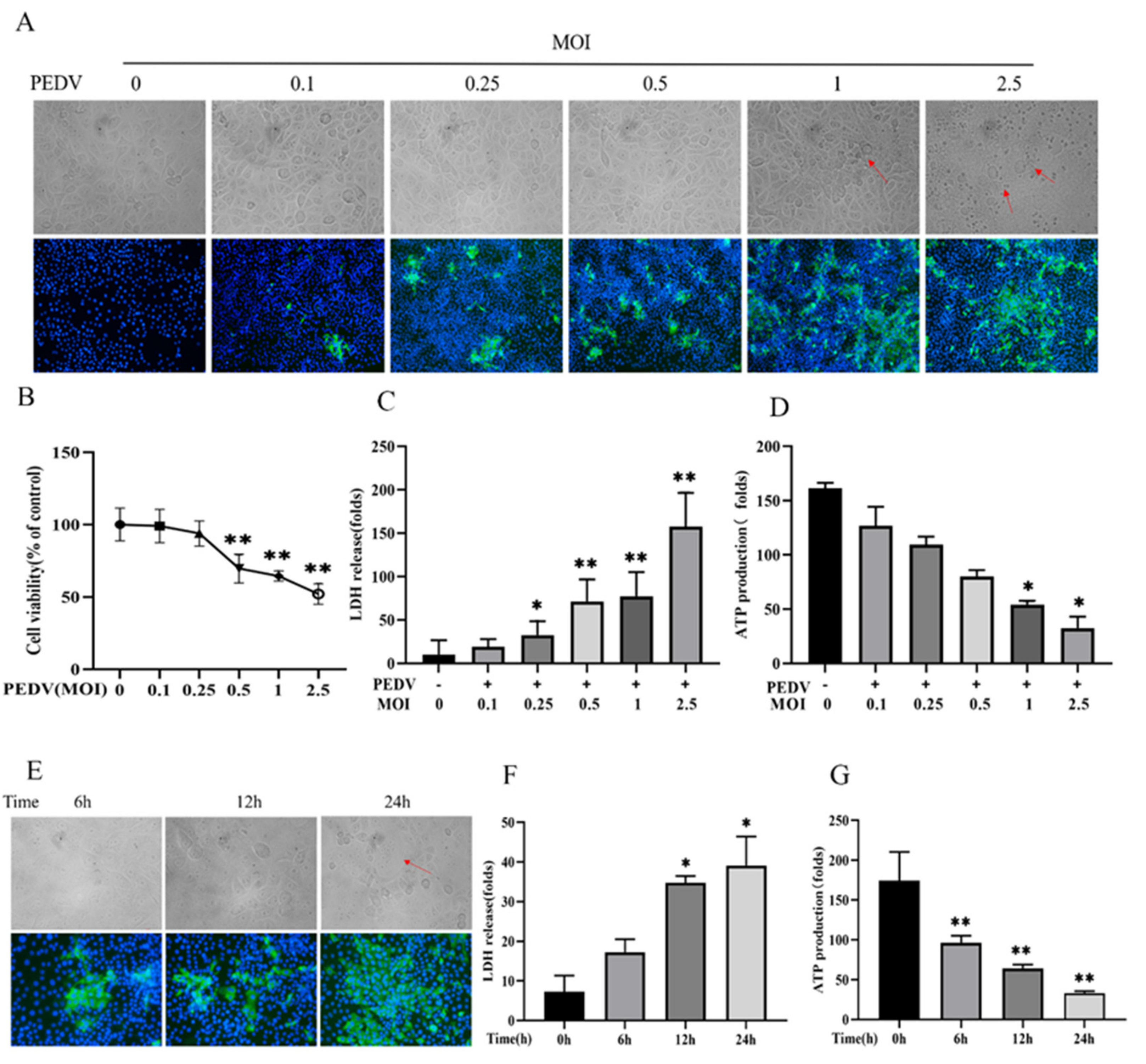
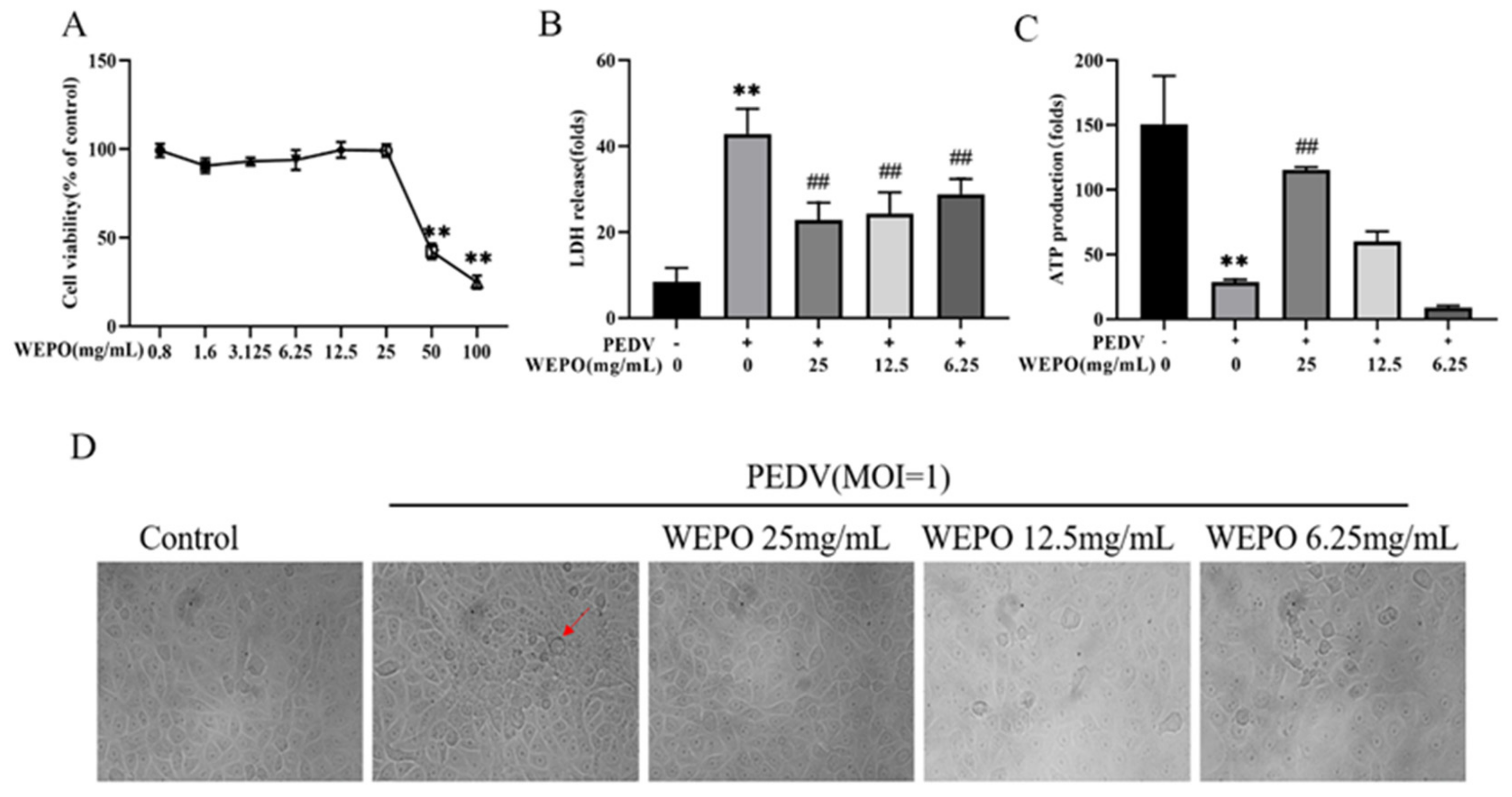
3.1. WEPO Attenuated PEDV-Infected Vero Cell Injury
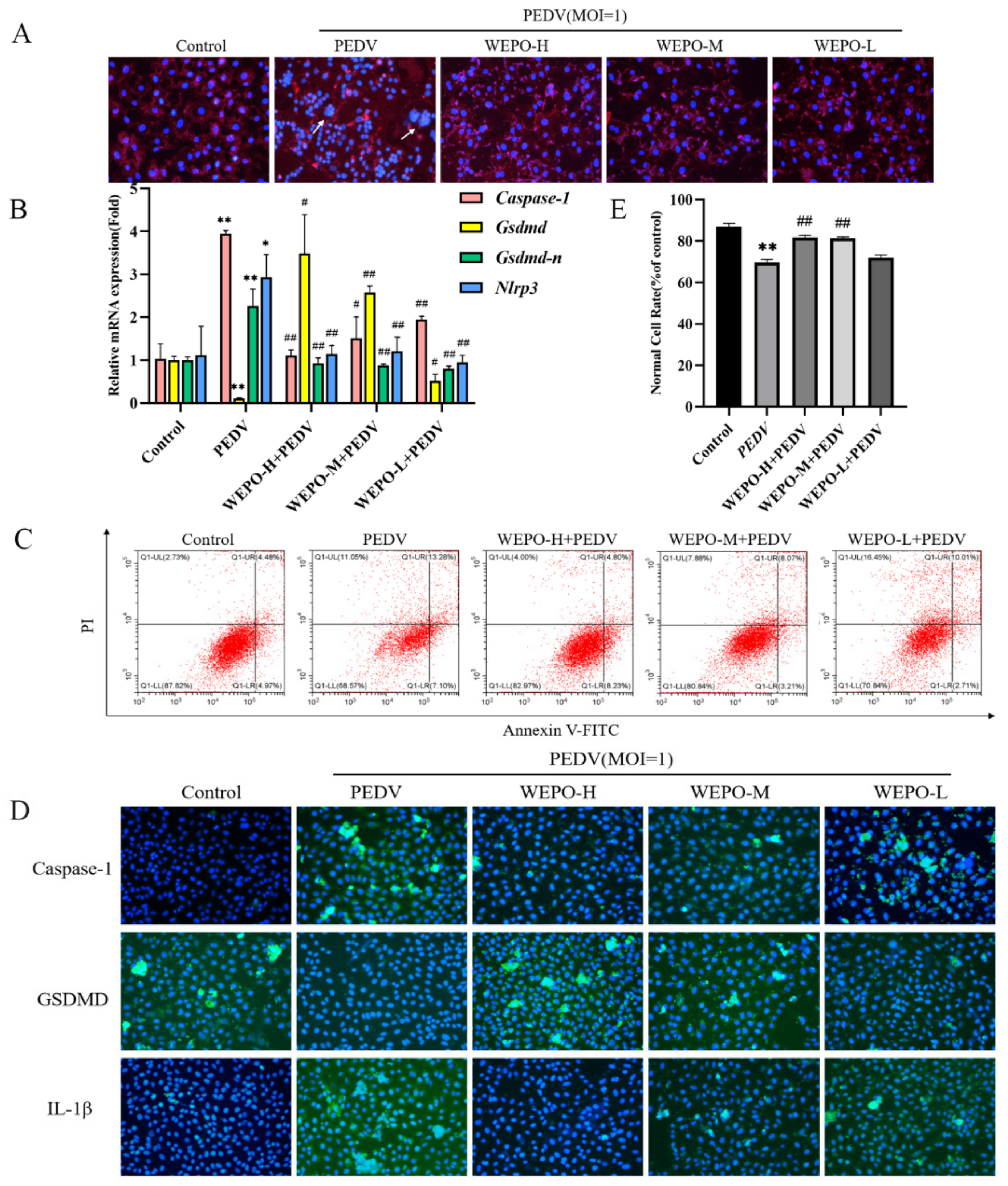
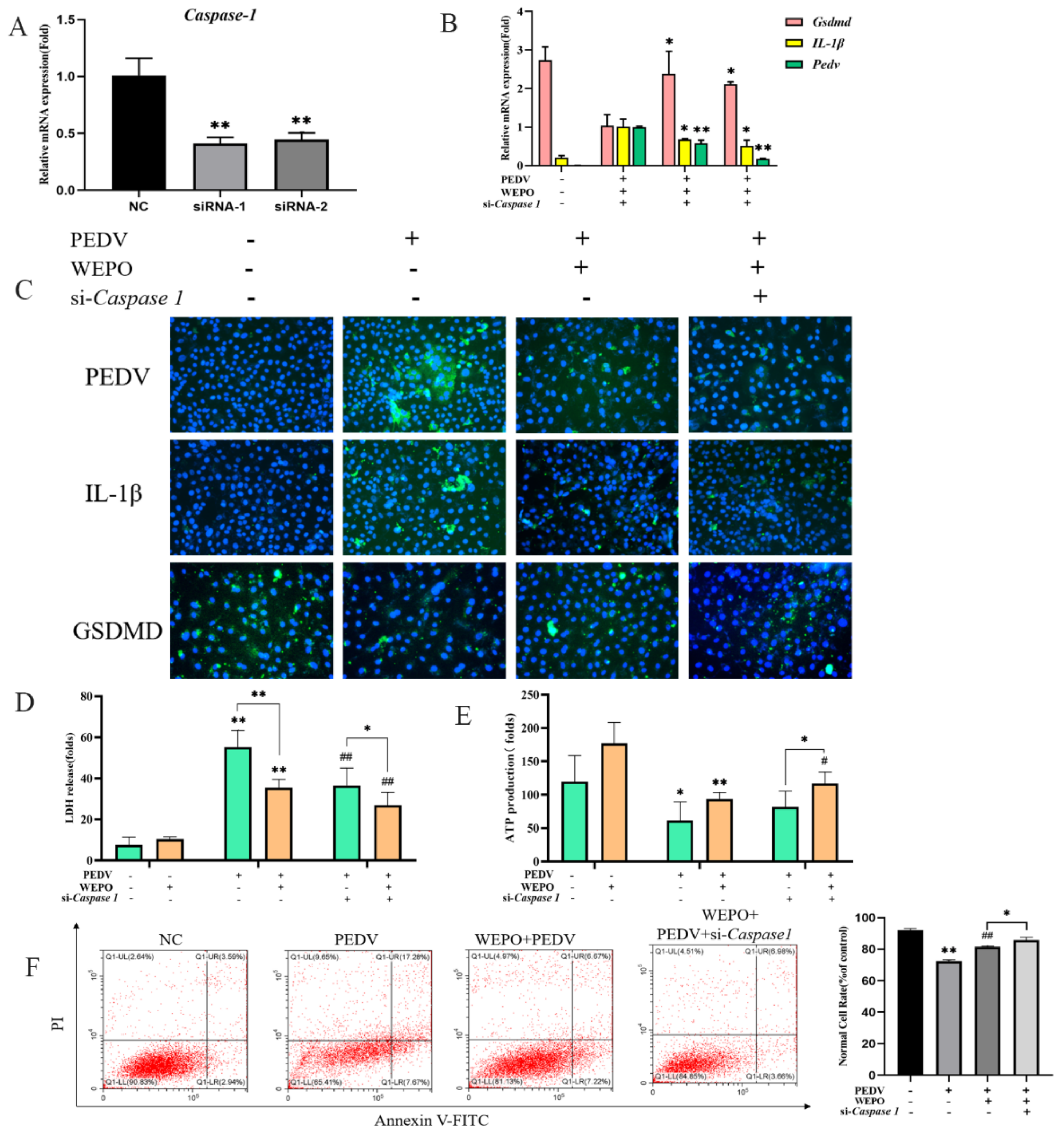
3.2. WEPO Inhibited Caspase-1/GSDMD-Mediated Pyroptosis in PEDV-Infected Vero Cells
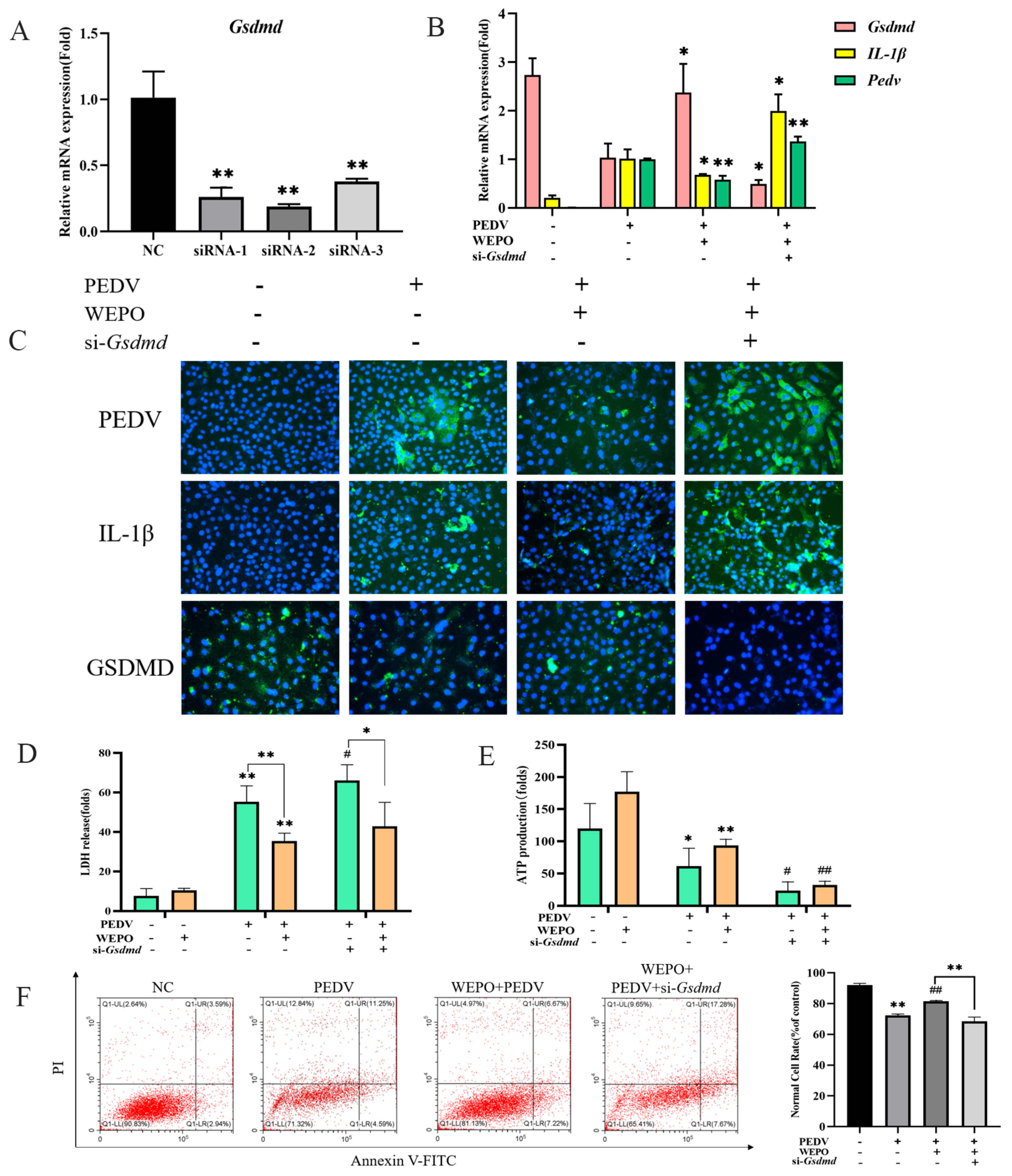
3.3. GSDMD Participates in the Pyroptosis of WEPO-Inhibited PEDV-Infected Vero Cells
3.4. Caspase-1 Is Needed for GSDMD-N Induction and Pyroptosis of WEPO in Response to PEDV Infection
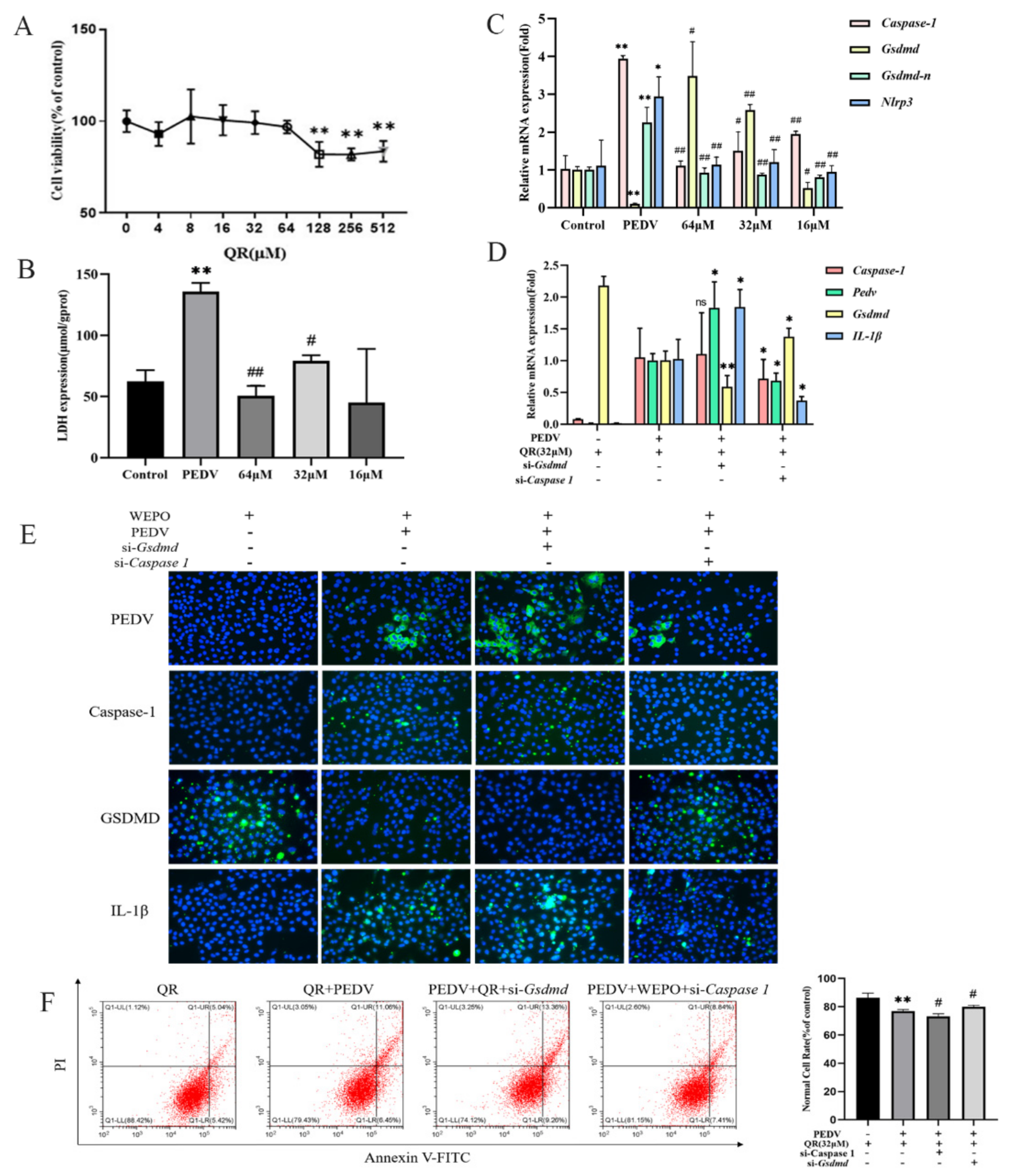
3.5. Quercetin, as One of the Main Components of Purslane, Participates in Cell Pyroptosis Mediated by Caspase-1/GSDMD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sueyoshi, M.; Tsuda, T.; Yamazaki, K.; Yoshida, K.; Nakazawa, M.; Sato, K.; Minami, T.; Iwashita, K.; Watanabe, M.; Suzuki, Y. An immunohistochemical investigation of porcine epidemic diarrhoea. J. Comp. Pathol. 1995, 113, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Shibata, I.; Tsuda, T.; Mori, M.; Ono, M.; Uruno, K. Isolation of porcine epidemic diarrhea virus in porcine cell cultures and experimental infection of pigs of different ages. Vet. Microbiol. 2000, 72, 173–182. [Google Scholar] [CrossRef]
- Ruiqin, S. Outbreak of Porcine Epidemic Diarrhea in Suckling Piglets, China. In Emerging Infectious Diseases; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019. [Google Scholar]
- Zhang, Q.-Z.; Shi, K.; Yoo, D. Suppression of type I interferon production by porcine epidemic diarrhea virus and degradation of CREB-binding protein by nsp1. Virology 2016, 489, 252–268. [Google Scholar] [CrossRef]
- Li, S.; Yang, J.; Zhu, Z.; Zheng, H. Porcine Epidemic Diarrhea Virus and the Host Innate Immune Response. Pathogens 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Rui, Z.; Xiao-Lin, W.U.; Wen-Wei, L.; Cardiology, D.O.; Hospital, X.C. Relationship of NLRP3 Inflammasome with TLR3, PKR and the Research Progress in the Atherosclerosis. Med. Recapitul. 2016, 22, 685–687. [Google Scholar]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine epidemic diarrhea virus (PEDV): An update on etiology, transmission, pathogenesis, and prevention and control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, L.; Xie, Y.; Chen, Z.; Yang, S.; Yin, B.; Li, G.; Guo, H.; Lin, S.; Wu, J. Antiviral activity of Portulaca oleracea L. extracts against porcine epidemic diarrhea virus by partial suppression on myd88/NF-κb activation in vitro. Microb. Pathog. 2021, 154, 104832. [Google Scholar] [CrossRef]
- Zhou, Y.X.; Xin, H.L.; Rahman, K.; Wang, S.J.; Peng, C.; Zhang, H. Portulaca oleracea L.: A review of phytochemistry and pharmacological effects. Biomed. Res. Int. 2015, 2015, 925631. [Google Scholar] [CrossRef]
- Zhu, H.B.; Wang, Y.Z.; Liu, Y.X.; Xia, Y.L.; Tang, T. Analysis of flavonoids in Portulaca oleracea L. by UV-vis spectrophotometry with comparative study on different extraction technologies. Food Anal. Methods 2010, 3, 90–97. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, Y.; Zhang, D.; Wu, X.; Yu, J.; Zhang, Z.; Li, Y.; Sun, C.; Tang, Z.; Liu, L. Protective Effects of Polysaccharide Extracted from Portulacae oleracea L. on Colitis Induced by Dextran Sulfate Sodium. J. Med. Food 2020, 23, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.J.; Bhardwaj, J.; Puri, S.K. mRNA expression of cytokines and its impact on outcomes after infection with lethal and nonlethal Plasmodium vinckei parasites. Parasitol. Res. 2012, 110, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.S.; Zhao, Y.H.; Zhong, Y.; Li, X.Z.; Pu, J.X.; Luo, Y.C.; Zhou, Q.L. Neferine inhibits LPS-ATP-induced endothelial cell pyroptosis via regulation of ROS/NLRP3/Caspase-1 signaling pathway. Inflamm. Res. 2019, 68, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sun, Y.; Song, M.; Song, Y.; Fang, Y.; Zhang, Q.; Li, X.; Song, N.; Ding, J.; Lu, M.; et al. NLRP3/caspase-1/GSDMD-mediated pyroptosis exerts a crucial role in astrocyte pathological injury in mouse model of depression. JCI Insight 2021, 6, e146852. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Luo, B.; Wu, X.; Guan, F.; Yu, X.; Zhao, L.; Ke, X.; Wu, J.; Yuan, J. Cisplatin Induces Pyroptosis via Activation of MEG3/NLRP3/caspase-1/GSDMD Pathway in Triple-Negative Breast Cancer. Int. J. Biol. Sci. 2021, 17, 2606–2621. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, X.; Jia, G.; Li, T.; Li, Y.; Zhao, R.; Wang, Y. Network pharmacology based research into the effect and potential mechanism of Portulaca oleracea L. polysaccharide against ulcerative colitis. Comput. Biol. Med. 2023, 161, 106999. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Hu, Z.; Fan, M.; Shao, Z.; Yu, Q.; Li, X. Development of an indirect ELISA to detect PEDV specific IgA antibody based on a PEDV epidemic strain. BMC Vet. Res. 2022, 18, 319. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.M.; Hou, Y.; Marthaler, D.G.; Gao, X.; Liu, X.; Zheng, L.; Saif, L.J.; Wang, Q. Attenuation of an original US porcine epidemic diarrhea virus strain PC22A via serial cell culture passage. Vet. Microbiol. 2017, 201, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yin, B.; Lian, R.; Zhang, L.; Liu, Y.; Li, Z.; Sun, C.; Liu, Y.; Shifa, L.G.; Fu, J.; et al. Anti-colitis Mechanism of Herba Portulacae Based on Network Pharmacology. World Chin. Med. 2020, 15, 3748–3760. (In Chinese) [Google Scholar]
- Kuriakose, T.; Zheng, M.; Neale, G.; Kanneganti, T.D. IRF1 Is a Transcriptional Regulator of ZBP1 Promoting NLRP3 Inflammasome Activation and Cell Death during Influenza Virus Infection. J. Immunol. 2018, 200, 1489–1495. [Google Scholar] [CrossRef]
- Deng, X.; Zou, W.; Xiong, M.; Wang, Z.; Qiu, J. Human Parvovirus Infection of Human Airway Epithelia Induces Pyroptotic Cell Death by Inhibiting Apoptosis. J. Virol. 2017, 91, e01533-17. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Gao, J.; Taxman, D.J.; Ting, J.P.; Su, L. HIV-1 Infection Induces Interleukin-1β Production via TLR8 Protein-dependent and NLRP3 Inflammasome Mechanisms in Human Monocytes. J. Biol. Chem. 2014, 289, 21716–21726. [Google Scholar] [CrossRef] [PubMed]
- Doitsh, G.; Galloway, N.L.K.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Munoz-Arias, I. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, C. Porcine epidemic diarrhea virus induces caspase-independent apoptosis through activation of mitochondrial apoptosis-inducing factor. Virology 2014, 460–461, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Cao, Y.; Wu, S.; Wu, Z.; Bao, W. miR-129a-3p Inhibits PEDV Replication by Targeting the EDA-Mediated NF-κB Pathway in IPEC-J2 Cells. Int. J. Mol. Sci. 2021, 22, 8133. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, H.; Li, X.; Yi, B.; Huang, L.; Hu, Z.; Li, A.; Du, J.; Li, Y.; Zhang, W. Vitamin D/VDR attenuate cisplatin-induced AKI by down-regulating NLRP3/Caspase-1/GSDMD pyroptosis pathway. J. Steroid. Biochem. Mol. Biol. 2021, 206, 105789. [Google Scholar] [CrossRef]
- Fan, Y.M.; Wei, Y.Y.; Wang, H.R.; Yu, G.; Zhang, Y.N.; Hao, Z. Inhibitory effect of Portulaca oleracea L. aqueous extract and juice on NLRP3 inflammasome activation in an ulcerative colitis mouse model. Environ. Sci. Pollut. Res. Int. 2023, 30, 86380–86394. [Google Scholar] [CrossRef]
- Yu, R.; Zhong, J.; Zhou, Q.; Ren, W.; Liu, Z.; Bian, Y. Kaempferol prevents angiogenesis of rat intestinal microvascular endothelial cells induced by LPS and TNF-α via inhibiting VEGF/Akt/p38 signaling pathways and maintaining gut-vascular barrier integrity. Chem. Biol. Interact. 2022, 366, 110135. [Google Scholar] [CrossRef]
- Zhang, H.X.; Li, Y.Y.; Liu, Z.J.; Wang, J.F. Quercetin effectively improves LPS-induced intestinal inflammation, pyroptosis, and disruption of the barrier function through the TLR4/NF-κB/NLRP3 signaling pathway in vivo and in vitro. Food Nutr. Res. 2022, 66, 8948. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Li, X.; Zhang, L.; Ren, L. Flavonoids as Potential Antiviral Agents for Porcine Viruses. Pharmaceutics 2022, 14, 1793. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, Y.; Yang, S.; Yin, B.; Zhao, Z.; Huang, Z.; Wu, J.; Lin, S.; Wang, X. Water Extract of Portulaca Oleracea Inhibits PEDV Infection-Induced Pyrolysis by Caspase-1/GSDMD. Curr. Issues Mol. Biol. 2023, 45, 10211-10224. https://doi.org/10.3390/cimb45120637
Zhang Y, Liu Y, Yang S, Yin B, Zhao Z, Huang Z, Wu J, Lin S, Wang X. Water Extract of Portulaca Oleracea Inhibits PEDV Infection-Induced Pyrolysis by Caspase-1/GSDMD. Current Issues in Molecular Biology. 2023; 45(12):10211-10224. https://doi.org/10.3390/cimb45120637
Chicago/Turabian StyleZhang, Yu, Yueyue Liu, Shifa Yang, Bin Yin, Zengcheng Zhao, Zhongli Huang, Jiaqiang Wu, Shuqian Lin, and Xin Wang. 2023. "Water Extract of Portulaca Oleracea Inhibits PEDV Infection-Induced Pyrolysis by Caspase-1/GSDMD" Current Issues in Molecular Biology 45, no. 12: 10211-10224. https://doi.org/10.3390/cimb45120637