
Ginkgo biloba: Antioxidant Activity and In Silico Central Nervous System Potential

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Studies of Similar Structures
2.2. Molecular Docking Studies
2.3. ADMETx Studies
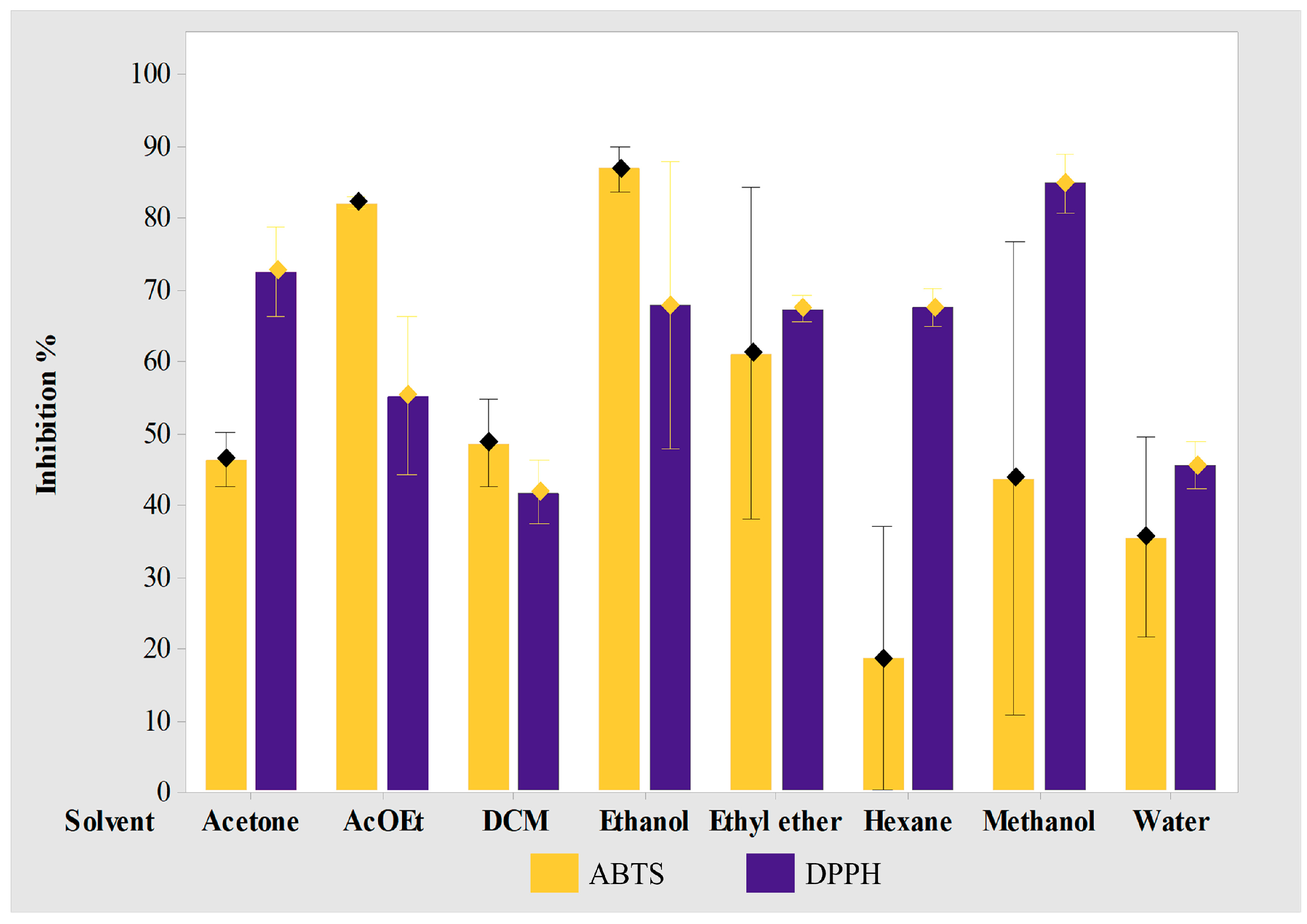
2.4. Extraction and Antioxidant Studies
3. Results and Discussion
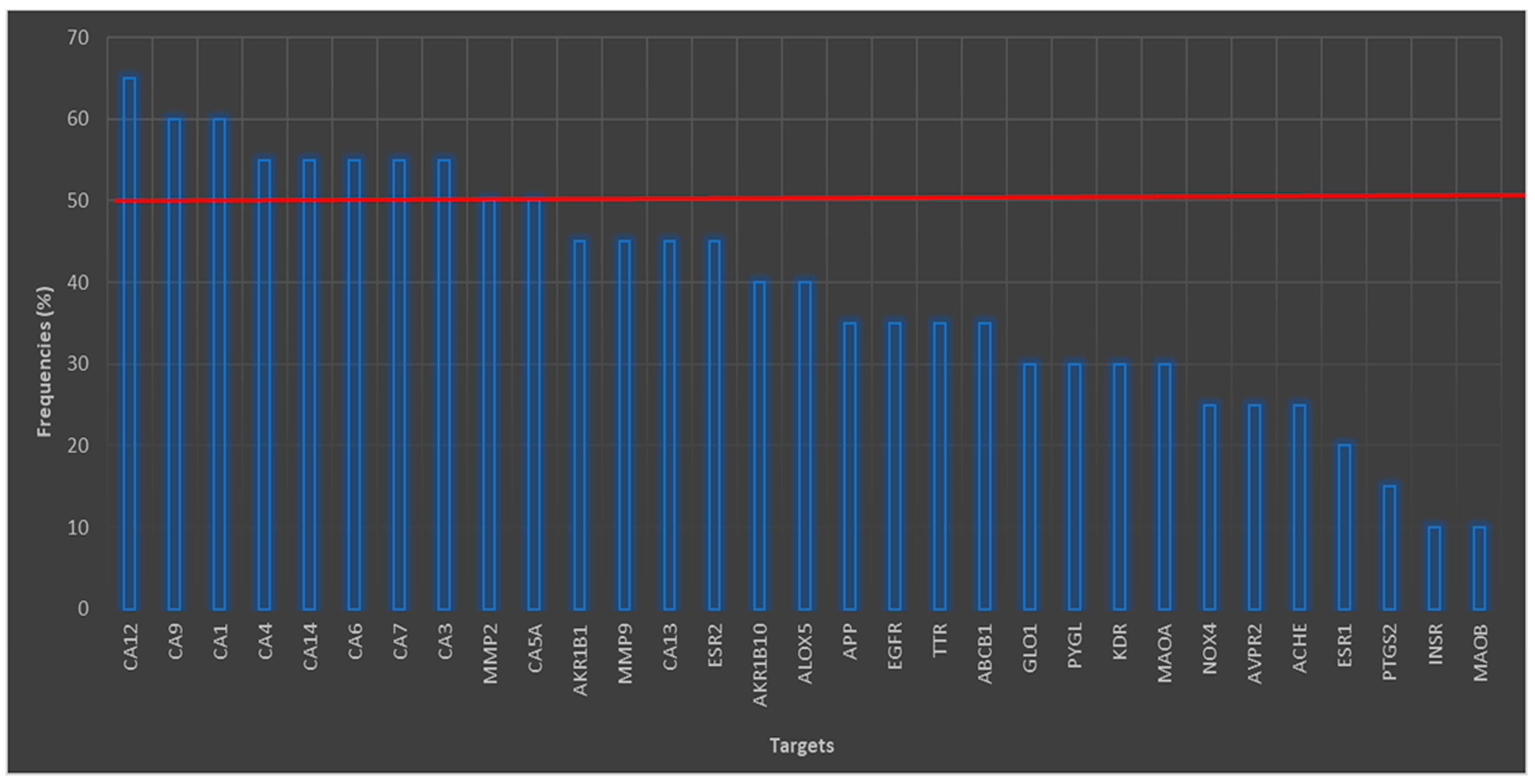
3.1. Analyses for Similar Structures
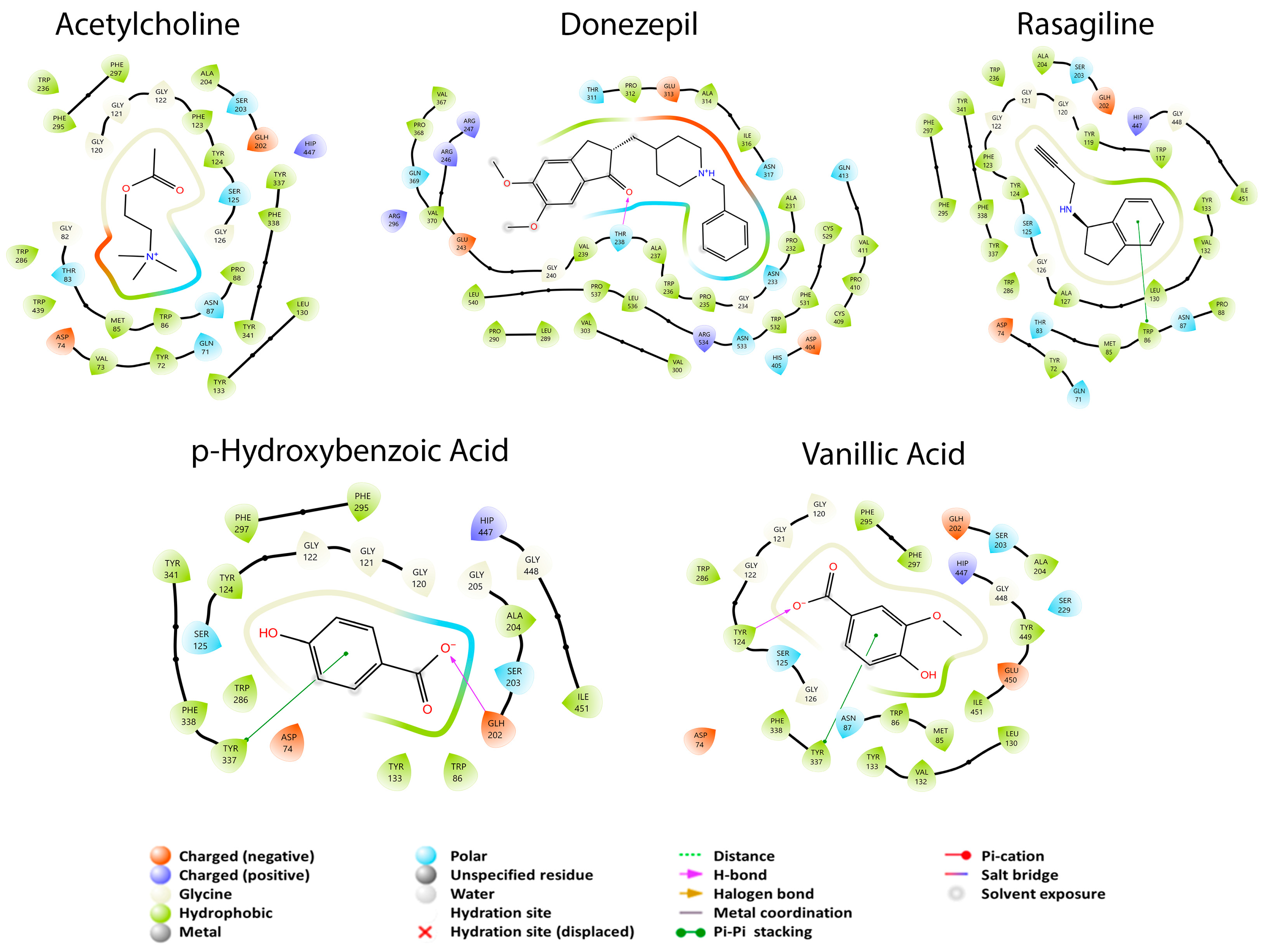
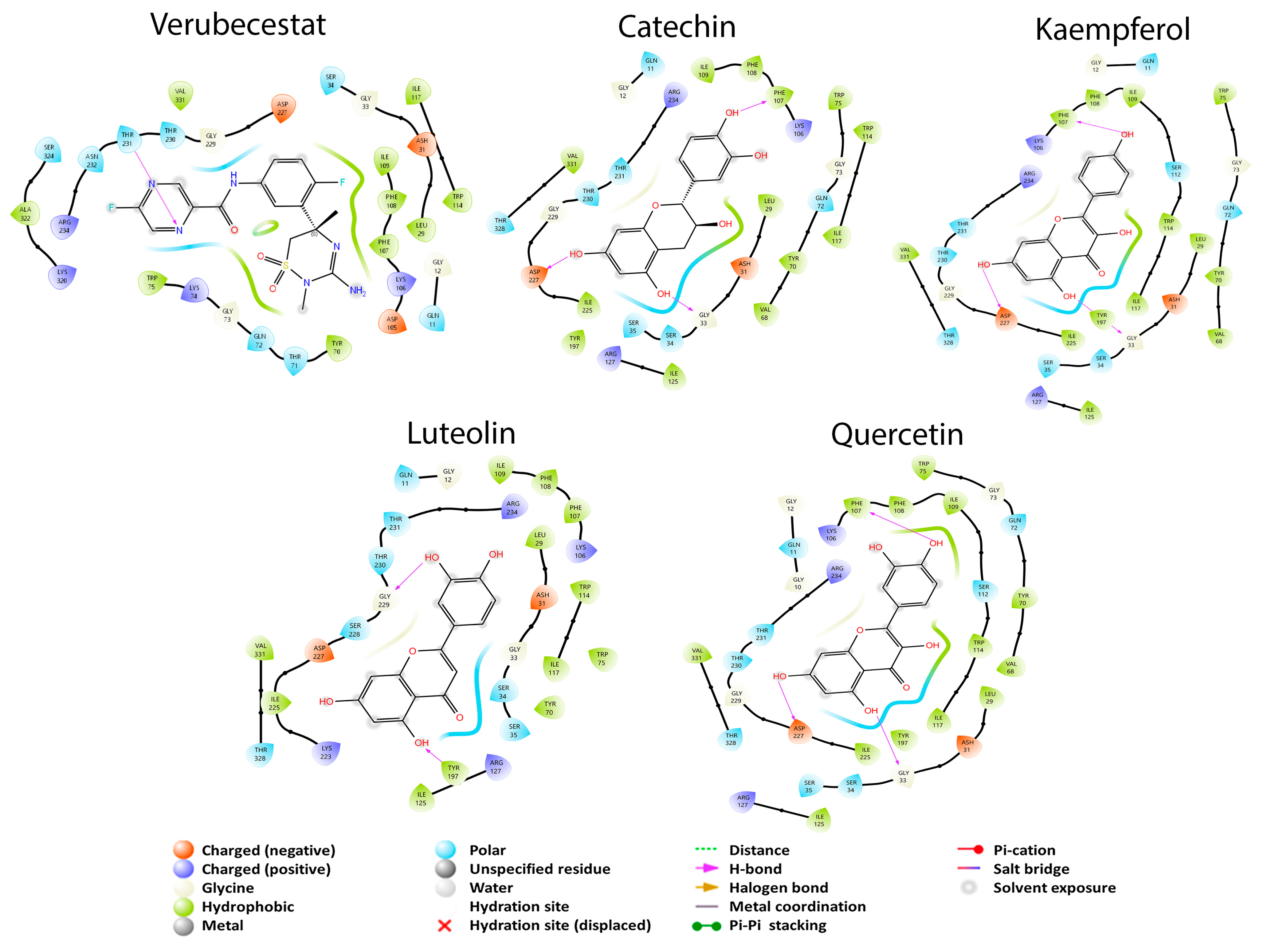
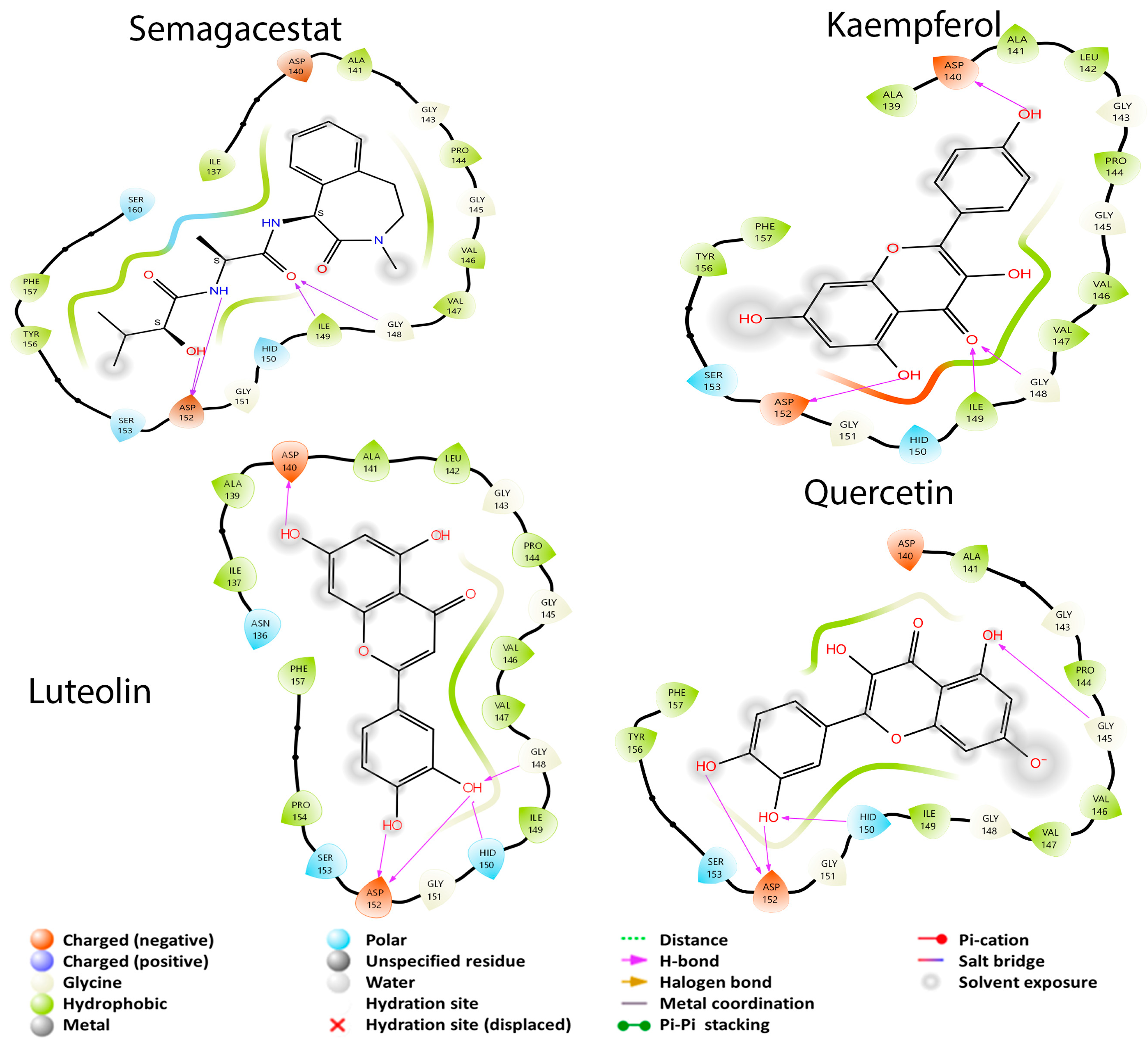
3.2. Molecular Docking Studies
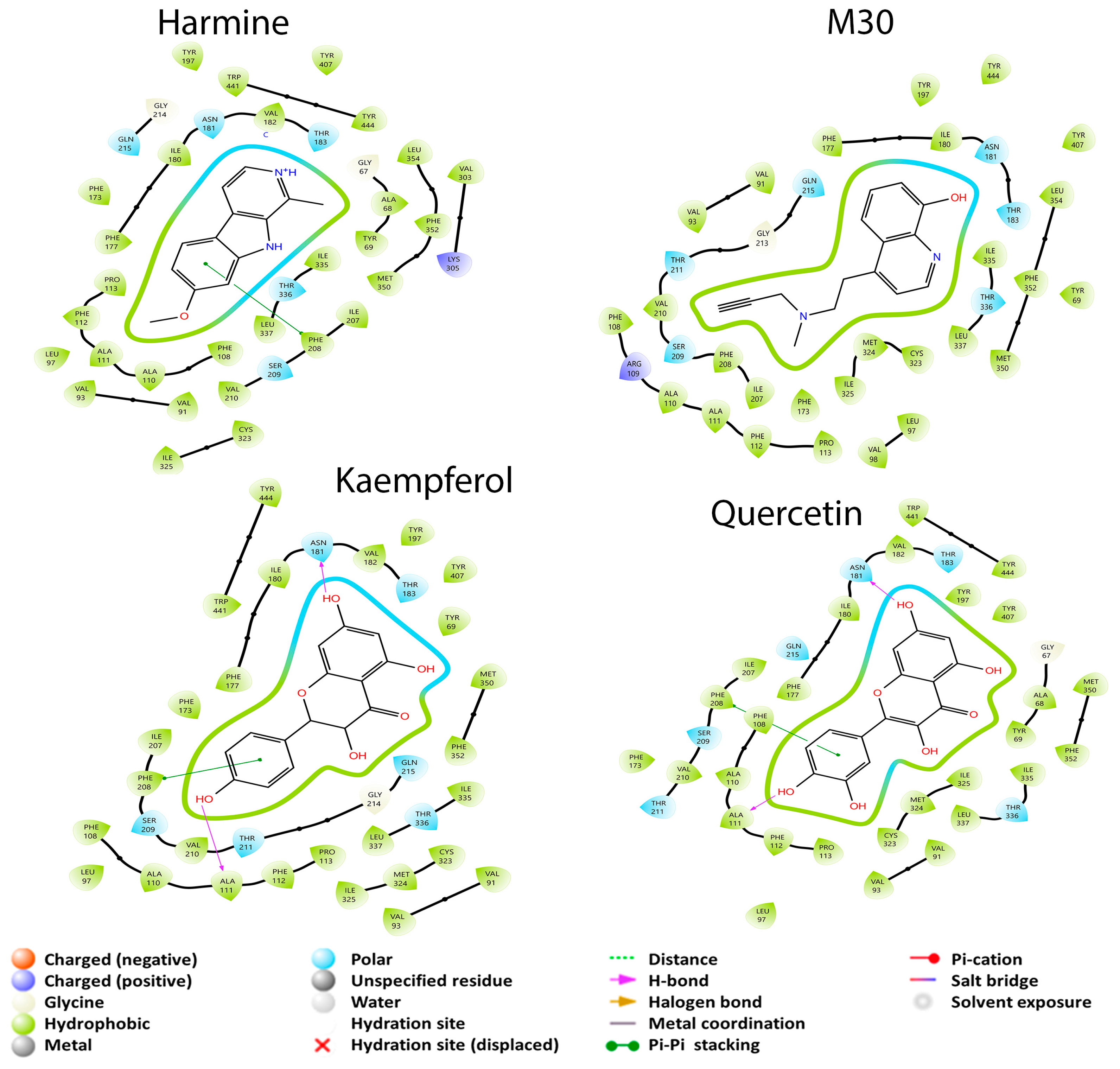
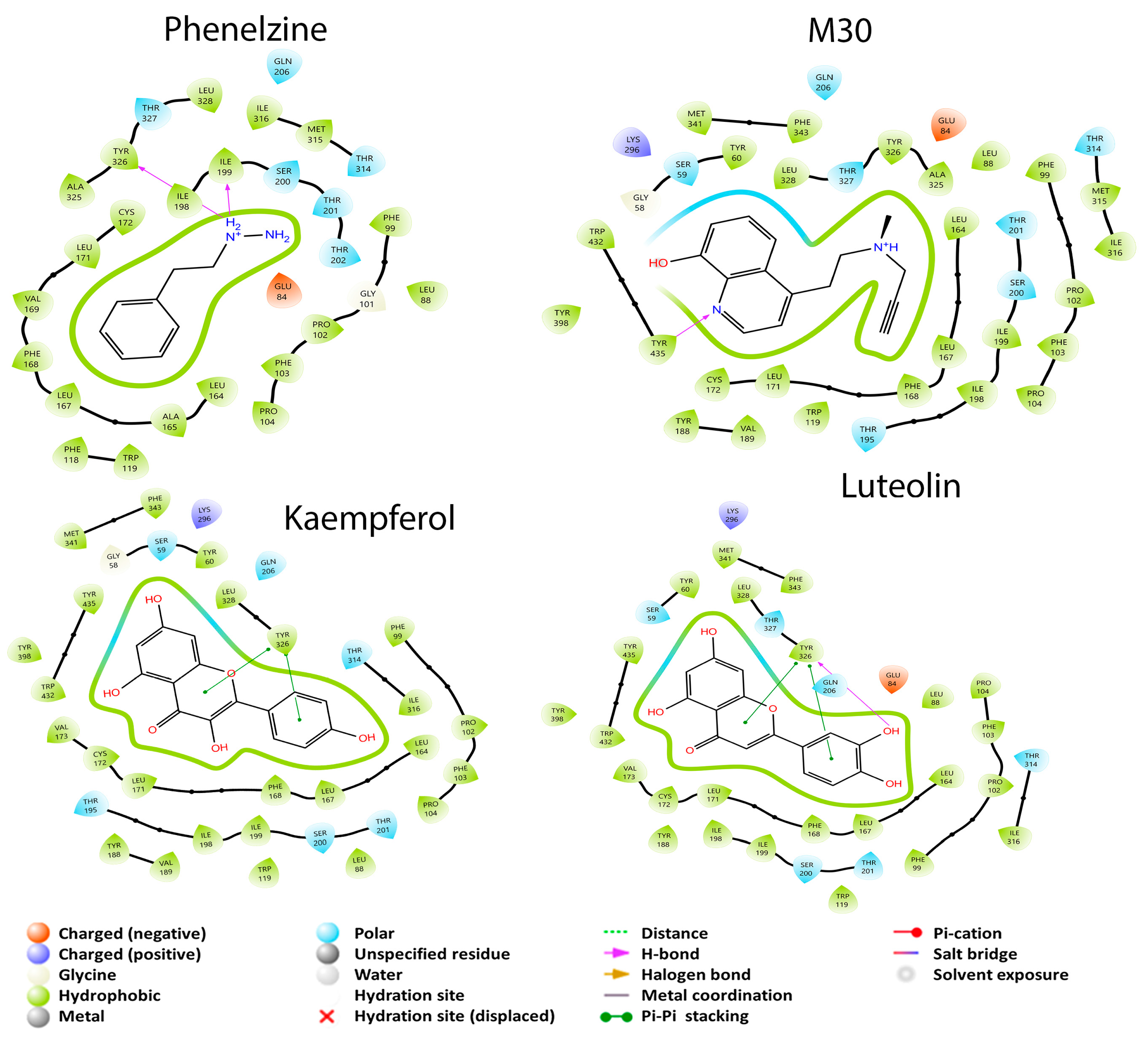
3.2.1. Monoamine Oxidase Pathway
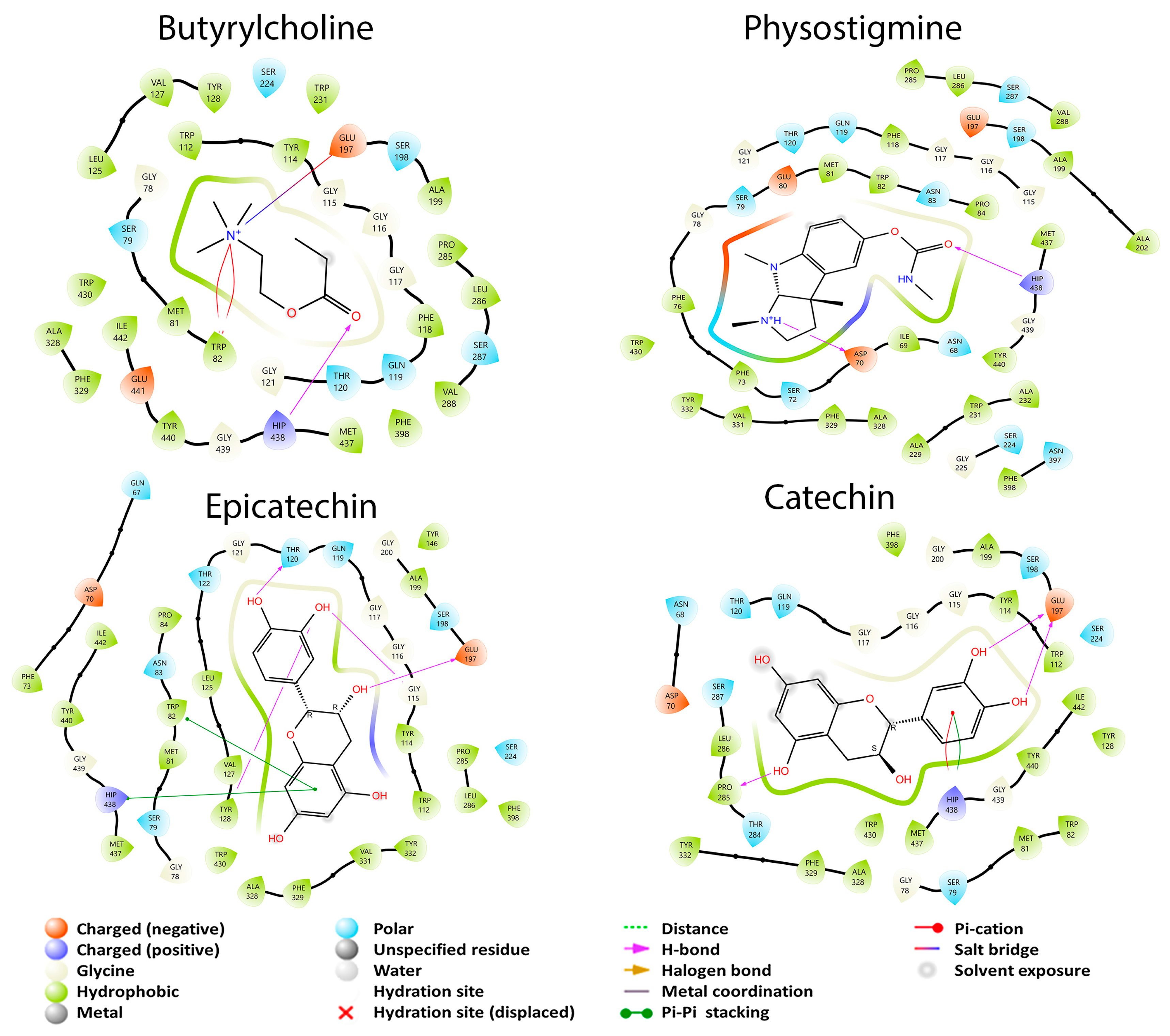
3.2.2. Enzymatic Cholinergic Pathway
3.2.3. APP Secretase Regulation
3.3. ADMETx Studies
3.4. Extraction and Antioxidant Studies
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, S.K.; Srivastav, S.; Castellani, R.J.; Plascencia-Villa, G.; Perry, G. Neuroprotective and Antioxidant Effect of Ginkgo biloba Extract Against AD and Other Neurological Disorders. Neurotherapeutics 2019, 16, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Al-kuraishy, H.M.; Al-Gareeb, A.I.; Kaushik, A.; Kujawska, M.; Batiha, G.E.S. Ginkgo biloba in the management of the COVID-19 severity. Arch. Pharm. 2022, 355, 2200188. [Google Scholar] [CrossRef] [PubMed]
- Labkovich, M.; Jacobs, E.B.; Bhargava, S.; Pasquale, L.R.; Ritch, R. Ginkgo biloba Extract in Ophthalmic and Systemic Disease, with a Focus on Normal-Tension Glaucoma. Asia Pac. J. Ophthalmol. 2020, 9, 215–225. [Google Scholar] [CrossRef]
- Mei, N.; Guo, X.; Ren, Z.; Kobayashi, D.; Wada, K.; Guo, L. Review of Ginkgo biloba-induced toxicity, from experimental studies to human case reports. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2017, 35, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Gregory, J.; Vengalasetti, Y.V.; Bredesen, D.E.; Rao, R.V. Biomolecules Neuroprotective Herbs for the Management of Alzheimer’s Disease. Biomolecules 2021, 11, 543. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Zhang, J.; Wang, S. Advances in the chemical constituents and chemical analysis of Ginkgo biloba leaf, extract, and phytopharmaceuticals. J. Pharm. Biomed. Anal. 2021, 193, 113704. [Google Scholar] [CrossRef]
- Liu, H.; Ye, M.; Guo, H. An Updated Review of Randomized Clinical Trials Testing the Improvement of Cognitive Function of Ginkgo biloba Extract in Healthy People and Alzheimer’s Patients. Front. Pharmacol. 2020, 10, 1688. [Google Scholar] [CrossRef]
- Brailowsky, S.; Montiel, T.; Hernández-Echeagaray, E.; Flores-Hernández, J.; Hernández-Pineda, R. Effects of a Ginkgo biloba extract on two models of cortical hemiplegia in rats. Restor. Neurol. Neurosci. 1991, 3, 267–274. [Google Scholar] [CrossRef]
- Ban, C.; Park, J.B.; Cho, S.; Kim, H.R.; Kim, Y.J.; Bae, H.; Kim, C.; Kang, H.; Jang, D.; Shin, Y.S.; et al. Characterization of Ginkgo biloba Leaf Flavonoids as Neuroexocytosis Regulators. Molecules 2020, 25, 1829. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, C.W.; Shi, J.; Lin, Z.X. Effects of Ginkgo biloba on dementia: An overview of systematic reviews. J. Ethnopharmacol. 2017, 195, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Horbowicz, M.; Wiczkowski, W.; Góraj-Koniarska, J.; Miyamoto, K.; Ueda, J.; Saniewski, M. Effect of Methyl Jasmonate on the Terpene Trilactones, Flavonoids, and Phenolic Acids in Ginkgo biloba L. Leaves: Relevance to Leaf Senescence. Molecules 2021, 26, 4682. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, C.; Han, Y.; Meng, X.; Zhang, Y.; Chu, H.; Ma, H. Gingko biloba Extract (EGb) Inhibits Oxidative Stress in Neuro 2A Cells Overexpressing APPsw. BioMed Res. Int. 2019, 2019, 7034983. [Google Scholar] [CrossRef] [PubMed]
- Janßen, I.M.; Sturtz, S.; Skipka, G.; Zentner, A.; Garrido, M.V.; Busse, R. Ginkgo biloba in Alzheimer’s disease: A systematic review. Wien. Med. Wochenschr. 2010, 160, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, M.; Ohta, Y.; Shimizu, M.; Maruyama, J.; Mochizuki, M. Meta-analysis of the efficacy and safety of Ginkgo biloba extract for the treatment of dementia. J. Pharm. Health Care Sci. 2015, 1, 14. [Google Scholar] [CrossRef]
- Kandiah, N.; Ong, P.A.; Yuda, T.; Ng, L.L.; Mamun, K.; Merchant, R.A.; Chen, C.; Dominguez, J.; Marasigan, S.; Ampil, E.; et al. Treatment of dementia and mild cognitive impairment with or without cerebrovascular disease: Expert consensus on the use of Ginkgo biloba extract, EGb 761®. CNS Neurosci. Ther. 2019, 25, 288–298. [Google Scholar] [CrossRef]
- Canevelli, M.; Adali, N.; Kelaiditi, E.; Cantet, C.; Ousset, P.J.; Cesari, M. Effects of Gingko biloba supplementation in Alzheimer’s disease patients receiving cholinesterase inhibitors: Data from the ICTUS study. Phytomedicine 2014, 21, 888–892. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Carrasco-Carballo, A.; Mendoza-Lara, D.F.; Rojas-Morales, J.A.; Altriste, V.; Merino-Montiel, P.; Luna, F.; Sandoval-Ramirez, J. In silico Study of Coumarins Derivatives with Potential Use in Systemic Diseases. BRIAC 2022, 13, 240. [Google Scholar] [CrossRef]
- Filimonov, D.A.; Lagunin, A.A.; Gloriozova, T.A.; Rudik, A.V.; Druzhilovskii, D.S.; Pogodin, P.V.; Poroikov, V.V. Prediction of the biological activity spectra of organic compounds using the pass online web resource. Chem. Heterocycl. Compd. 2014, 50, 444–457. [Google Scholar] [CrossRef]
- Schrödinger, L. Schrödinger Release 2022-4: MacroModel; EE. UU: New York, NY, USA, 2021. [Google Scholar]
- Schrödinger, L. Schrödinger Release 2022-4: LigPrep; EE. UU: New York, NY, USA, 2021. [Google Scholar]
- Cheung, J.; Gary, E.N.; Shiomi, K.; Rosenberry, T.L. Structures of human acetylcholinesterase bound to dihydrotanshinone i and territrem B show peripheral site flexibility. ACS Med. Chem. Lett. 2013, 4, 1091–1096. [Google Scholar] [CrossRef]
- Nicolet, Y.; Lockridge, O.; Masson, P.; Fontecilla-Camps, J.C.; Nachon, F. Crystal Structure of Human Butyrylcholinesterase and of Its Complexes with Substrate and Products. J. Biol. Chem. 2003, 278, 41141–41147. [Google Scholar] [CrossRef] [PubMed]
- Son, S.Y.; Ma, J.; Kondou, Y.; Yoshimura, M.; Yamashita, E.; Tsukihara, T. Structure of human monoamine oxidase A at 2.2-Å resolution: The control of opening the entry for substrates/inhibitors. Proc. Natl. Acad. Sci. USA 2008, 105, 5739–5744. [Google Scholar] [CrossRef]
- Binda, C.; Wang, J.; Pisani, L.; Caccia, C.; Carotti, A.; Salvati, P.; Edmondson, D.E.; Mattevi, A. Structures of human monoamine oxidase B complexes with selective noncovalent inhibitors: Safinamide and coumarin analogs. J. Med. Chem. 2007, 50, 5848–5852. [Google Scholar] [CrossRef]
- Low, J.D.; Bartberger, M.D.; Cheng, Y.; Whittington, D.; Xue, Q.; Wood, S.; Allen, J.R.; Minatti, A.E. Diastereoselective synthesis of fused cyclopropyl-3-amino-2,4-oxazine β-amyloid cleaving enzyme (BACE) inhibitors and their biological evaluation. Bioorg. Med. Chem. Lett. 2018, 28, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.C.; Rajendra, E.; Yang, G.; Shi, Y.; Scheres, S.H.W. Sampling the conformational space of the catalytic subunit of human g-secretase. eLife 2015, 4, e11182. [Google Scholar] [CrossRef]
- Schrödinger, L. Schrödinger Release 2022-4: Schrödinger KNIME Extensions; EE. UU: New York, NY, USA, 2021. [Google Scholar]
- Schrödinger, L. Schrödinger Release 2022-4: Glide; EE. UU: New York, NY, USA, 2021. [Google Scholar]
- Schrödinger, L. Schrödinger Release 2022-4: QikProp; EE. UU: New York, NY, USA, 2021. [Google Scholar]
- Lagunin, A.; Zakharov, A.; Filimonov, D.; Poroikov, V. QSAR Modelling of Rat Acute Toxicity on the Basis of PASS Prediction. Mol. Inform. 2011, 30, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Rivera, J.A.; Herrera-García, M.; López-Castillo, G.N.; Sandoval-Ramírez, J.; Carrasco-Carballo, A. Epoxide of diosgenin acetate synthesis: DoE for diastereoselective yield optimization, antimicroibial, antioxidant and antiproliferative effects. Steroids 2023, 197, 109256. [Google Scholar] [CrossRef]
- Soto-Ortiz, S.; López-Castillo, G.N.; Alatriste, V.; Marin-Torres, L.J.; Morales-Sandoval, I.P.; Carrasco-Carballo, A. Loeselia mexicana: Antioxidant and antimicrobial properties by Soxhlet differential extraction. GSC Biol. Pharm. Sci. 2023, 2023, 115–120. [Google Scholar] [CrossRef]
- Monroy, L.E.; Vargas, M.P.S. Anhidrasa carbónica, nuevas perspectivas. Neumol. Cirugía Tórax 2010, 69, 200–209. Available online: https://www.medigraphic.com/pdfs/neumo/nt-2010/nt104d.pdf (accessed on 1 September 2023).
- Jasiecki, J.; Szczoczarz, A.; Cysewski, D.; Lewandowski, K.; Skowron, P.; Waleron, K.; Wasąg, B. Butyrylcholinesterase-Protein Interactions in Human Serum. Int. J. Mol. Sci. 2021, 22, 662. [Google Scholar] [CrossRef]
- Baumkötter, F.; Schimidt, N.; Vargas, C.; Schilling, S.; Weber, R.; Wagner, K.; Fiedler, S.; Klug, W.; Radzimanowsli, J.; Nickolaus, S.; et al. Amyloid Precursor Protein Dimerization and Synaptogenic Function Depend on Copper Binding to the Growth Factor-Like Domain. J. Neurosci. 2014, 34, 11159–11172. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Brindisi, M.; Tang, J. Developing β-secretase inhibitors for treatment of Alzheimer’s disease. J. Neurochem. 2012, 120, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, M.S. γ-Secretase inhibitors and modulators for Alzheimer’s disease. J. Neurochem. 2012, 120, 89–98. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Oxidative Stress, Amyloid-β Peptide, and Altered Key Molecular Pathways in the Pathogenesis and Progression of Alzheimer’s Disease. J. Alzheimers Dis. 2018, 62, 1345–1367. [Google Scholar] [CrossRef]
- Hur, J.Y. γ-Secretase in Alzheimer’s disease. Exp. Mol. Med. 2022, 54, 433–446. [Google Scholar] [CrossRef]
- Shih, J.C.; Isoenzymes, M.O. Functions and Targets for Behavior and Cancer Therapy. J. Neural Transm. 2018, 125, 1553–1566. [Google Scholar] [CrossRef]
- Kushal, S.; Wang, W.; Vaikari, V.P.; Kota, R.; Chen, K.; Yeh, T.S.; Jhaveri, N.; Groshen, S.L.; Olenyuk, B.Z.; Chen, T.C.; et al. Monoamine oxidase A (MAO A) inhibitors decrease glioma progression. Oncotarget 2016, 7, 13842–13853. [Google Scholar] [CrossRef]
- Hagenow, J.; Hagenow, S.; Grau, K.; Khanfar, M.; Hefke, L.; Proschak, E.; Stark, H. Reversible Small Molecule Inhibitors of MAO A and MAO B with Anilide Motifs. Drug Des. Dev. Ther. 2020, 14, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Margret, A.A.; Begum, T.N.; Parthasarathy, S.; Suvaithenamudhan, S. A Strategy to Employ Clitoria ternatea as a Prospective Brain Drug Confronting Monoamine Oxidase (MAO) Against Neurodegenerative Diseases and Depression. Nat. Prod. Bioprospect. 2015, 5, 293–306. [Google Scholar] [CrossRef]
- Tan, Y.-Y.; Jenner, P.; Chen, S.-D. Monoamine Oxidase-B Inhibitors for the Treatment of Parkinson’s Disease: Past, Present, and Future. J. Parkinsons Dis. 2022, 12, 477–493. [Google Scholar] [CrossRef]
- Eckroat, T.J.; Manross, D.L.; Cowan, S.C.; Tacrine-Based, M. Multitarget-Directed Acetylcholinesterase Inhibitors 2015–Present: Synthesis and Biological Activity. Int. J. Mol. Sci. 2020, 21, 5965. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Chávez, G.; Salceda, R. Enzimas Polifuncionales: El Caso de la Acetilcolinesterasa. REB 2008, 27, 44–51. Available online: https://www.redalyc.org/pdf/490/49011451002.pdf (accessed on 1 September 2023).
- Walczak-Nowicka, Ł.J.; Herbet, M. Acetylcholinesterase Inhibitors in the Treatment of Neurodegenerative Diseases and the Role of Acetylcholinesterase in their Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9290. [Google Scholar] [CrossRef]
- Pohanka, M. Inhibitors of Acetylcholinesterase and Butyrylcholinesterase Meet Immunity. Int. J. Mol. Sci. 2014, 15, 9809–9825. [Google Scholar] [CrossRef]
- Kandiah, N.; Pai, M.-C.; Senanarong, V.; Looi, I.; Ampil, E.; Park, K.W.; Karanam, A.K.; Christopher, S. Rivastigmine: The advantages of dual inhibition of acetylcholinesterase and butyrylcholinesterase and its role in subcortical vascular dementia and Parkinson’s disease dementia. Clin. Interv. Aging 2017, 18, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Delacour, H.; Dedome, E.; Courcelle, S.; Hary, B.; Ceppa, F. Butyrylcholinesterase deficiency. Ann. Biol. Clin. 2016, 74, 279–285. [Google Scholar] [CrossRef]
- Yang, G.; Zhou, R.; Gou, X.; Yan, C.; Lei, J.; Shi, Y. Structural basis of γ-secretase inhibition and modulation by small molecule drugs. Cell 2021, 184, 521–533. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Tang, J. Prospects of β-Secretase Inhibitors for the Treatment of Alzheimer’s Disease. Chem. Med. Chem. 2015, 10, 1463–1466. [Google Scholar] [CrossRef]
- Harrison, H.; Pegg, H.J.; Thompson, J.; Bates, C.; Shore, P. HIF1-alpha expressing cells induce a hypoxic-like response in neighbouring cancer cells. BMC Cancer 2018, 18, 674. [Google Scholar] [CrossRef]
- Bardaweel, S.; Aljanabi, R.; Sabbah, D.; Sweidan, K. Design, Synthesis, and Biological Evaluation of Novel MAO-A Inhibitors Targeting Lung Cancer. Molecules 2022, 27, 2887. [Google Scholar] [CrossRef]
- Azam, F.; Madi, A.M.; Ali, H.I. Molecular Docking and Prediction of Pharmacokinetic Properties of Dual Mechanism Drugs that Block MAO-B and Adenosine A2A Receptors for the Treatment of Parkinson’s Disease. J. Young Pharm. 2012, 4, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Qin, X.; Wang, J.; Zhang, M.; An, Q.; Xu, J.; Qu, X.; Cao, X.; Niu, B. Discovery of MAO-B Inhibitor with Machine Learning, Topomer CoMFA, Molecular Docking and Multi-Spectroscopy Approaches. Biomolecules 2022, 12, 1470. [Google Scholar] [CrossRef]
- Shakil, S. Molecular interaction of inhibitors with human brain butyrylcholinesterase. EXCLI J. 2021, 20, 1597–1607. [Google Scholar] [CrossRef]
- Ogata, N.; Tagishi, H.; Tsuji, M. Inhibition of Acetylcholinesterase by Wood Creosote and Simple Phenolic Compounds. Chem. Pharm. Bull. 2020, 68, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Budryn, G.; Majak, I.; Grzelczyk, J.; Szwajgier, D.; Rodríguez-Martínez, A.; Pérez-Sánchez, H. Hydroxybenzoic Acids as Acetylcholinesterase Inhibitors: Calorimetric and Docking Simulation Studies. Nutrients 2022, 14, 2476. [Google Scholar] [CrossRef] [PubMed]
- Seniya, C.; Khan, G.J.; Uchadia, K. Identification of Potential Herbal Inhibitor of Acetylcholinesterase Associated Alzheimer’s Disorders Using Molecular Docking and Molecular Dynamics Simulation. Biochem. Res. Int. 2014, 2014, 705451. [Google Scholar] [CrossRef]
- Galeana-Ascencio, R.A.; Mendieta, L.; Limón, D.I.; Gnecco, D.; Terán, J.L.; Orea, M.L.; Carrasco-Carballo, A. β-Secretase-1: In Silico Drug Reposition for Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 8164. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Youdim, K.A.; Shukitt-Hale, B.; Joseph, J.A. Flavonoids and the brain: Interactions at the blood–brain barrier and their physiological effects on the central nervous system. Free. Radic. Biol. Med. 2004, 37, 1683–1693. [Google Scholar] [CrossRef]
- Velásquez-Jiménez, D.; Corella-Salazar, D.A.; Zuñiga-Martínez, B.S.; Domínguez-Avila, J.A.; Montiel-Herrera, M.; Salazar-López, N.J.; Rodrigo-Garcia, J.; Villegas-Ochoa, M.A.; González-Aguilar, G.A. Phenolic compounds that cross the blood–brain barrier exert positive health effects as central nervous system antioxidants. Food Funct. 2021, 12, 10356–10369. [Google Scholar] [CrossRef]
- Faria, A.; Pestana, D.; Teixeira, D.; Azevedo, J.; Freitas, V.; Mateus, N.; Calhau, C. Flavonoid transport across RBE4 cells: A blood-brain barrier model. Cell. Mol. Biol. Lett. 2010, 15, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Ionescu-Tucker, A.; Cotman, C.W. Emerging roles of oxidative stress in brain aging and Alzheimer’s disease. Neurobiol. Aging 2021, 107, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Liu, J.; Wu, F.; Yew, D.T. Ginkgo biloba Extract in Alzheimer’s Disease: From Action Mechanisms to Medical Practice. Int. J. Mol. Sci. 2010, 11, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Kojder, K.; Zielonka-Brzezicka, J.; Wróbel, J.; Bosiacki, M.; Fabiańska, M.; Wróbel, M.; Sołek-Pastuszka, J.; Klimowicz, A. The Use of Ginkgo biloba L. as a Neuroprotective Agent in the Alzheimer’s Disease. Front. Pharmacol. 2021, 12, 775034. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Secondary Metabolites | References |
|---|---|---|
| Terpenoids | Bilobalide Ginkgolide A Ginkgolide B Ginkgolide C | [1,2,3,6,7,9,10] |
| Flavonoids | Epicatechin Catechin Apigenin Luteolin Quercetin Kaempferol Isorhamnetin | [1,2,6,10] |
| Carboxylic acids | Protocatechuic acid Caffeic acid Chlorogenic acid p-Hydroxybenzoic acid Vanillic acid p-Coumaric acid Ferulic acid | [6,11] |
| Saccharides | Glucose Rhamnose | [1,6] |
| Proteins | Diseases | Inhibitors | References |
|---|---|---|---|
| MAO-A | Alzheimer’s disease Glioma Depressive disorders | Moclobemide Clorgyline Tranylcypromine | [10,16,41,42,43,44] |
| MAO-B | Alzheimer’s disease Parkinson’s disease Depressive disorders | Rasagiline Selegiline Deprenil | [10,16,42,43,44,45] |
| AChE | Alzheimer’s disease Parkinson’s disease Huntington’s disease Depressive disorders Myasthenia gravis | Galantamine Huperzine A Rivastigmine Neostigmine Donepezil | [46,47,48,49,50] |
| BuChE | No physiological functions Alzheimer’s disease Myasthenia gravis | Neostigmine Huperzine A Rivastigmine | [35,49,51] |
| β-Secretase | Alzheimer’s disease | GRL-8234 CTS-21166 Verubecestat | [40,52,53] |
| γ-Secretase | Alzheimer’s disease | Avagacestat Semagacestat | [40,52] |
| Secondary Metabolite | MAO-A | MAO-B | AChE | BuChE | β-Secretase | γ-Secretase |
|---|---|---|---|---|---|---|
| Bilobalide | NI | NI | NI | −6.056 | −4.694 | −0.110 |
| Ginkgolide A | NI | NI | NI | −6.559 | −5.413 | NI |
| Ginkgolide B | NI | NI | NI | −6.350 | −6.132 | NI |
| Ginkgolide C | NI | NI | NI | −6.068 | −6.230 | NI |
| Isorhamnetin | −6.223 | −6.588 | NI | −6.328 | −5.835 | −5.763 |
| Kaempferol | −9.127 | −6.636 | NI | −6.160 | −6.445 | −5.955 |
| Quercetin | −7.872 | −6.626 | NI | −7.451 | −6.244 | −4.668 |
| Apigenin | −8.698 | −6.663 | NI | −6.047 | −6.359 | −5.938 |
| Vanillic acid | −6.314 | −6.227 | −6.038 | −5.990 | −4.505 | −4.754 |
| Caffeic acid | −5.089 | −5.891 | −4.843 | −5.715 | −4.564 | −5.445 |
| Catechin | −7.287 | NI | NI | −6.321 | −7.345 | −4.652 |
| Epicatechin | NI | NI | NI | −7.984 | −5.969 | −4.753 |
| Chlorogenic acid | −4.548 | −6.302 | NI | −5.773 | −4.571 | −4.721 |
| Ferulic acid | −5.650 | −6.346 | −5.373 | −5.669 | −4.805 | −4.717 |
| Luteolin | −6.587 | −6.667 | NI | −7.114 | −5.539 | −5.344 |
| p-Coumaric acid | −4.642 | −5.500 | −4.354 | −5.751 | −4.753 | −4.647 |
| p-Hydroxybenzoic acid | −6.510 | −5.902 | −6.209 | −6.294 | −4.868 | −4.828 |
| Protocatechuic acid | −6.767 | −6.079 | −5.815 | −6.488 | −4.854 | −4.718 |
| (L)-Rhamnose | −5.859 | −5.901 | −5.780 | −5.822 | −4.892 | NI |
| (D)-Glucose | −5.498 | −5.627 | −5.695 | −6.941 | −4.848 | −5.801 |
| Harmine | −6.916 | - | - | - | - | - |
| Phenelzine | - | −5.766 | - | - | - | - |
| M30 | −5.956 | −6.104 | - | - | - | - |
| Acetylcholine | - | - | −3.546 | - | - | - |
| BuCh | - | - | - | −4.341 | - | - |
| Donezepil | - | - | −4.509 | - | - | - |
| Rasagiline | - | −6.060 | −5.145 | - | - | - |
| Physostigmine | - | - | - | −5.874 | - | - |
| Verubecestat | - | - | - | - | −4.298 | - |
| Semagacestat | - | - | - | - | - | −4.446 |
| Secondary Metabolite | MW (g/mol) | Lipophilicity Log P | BBB | Water Solubility Log S | GI Absorption | Rule of Three | Rule of Five |
|---|---|---|---|---|---|---|---|
| Bilobalide | 326.3 | 0.07 | No | −1.12 | High | 0 | 0 |
| Ginkgolide A | 408.4 | 0.61 | No | −1.59 | High | 0 | 0 |
| Ginkgolide B | 424.4 | −0.2 | No | −0.77 | Low | 0 | 0 |
| Ginkgolide C | 440.4 | −0.98 | No | 0.06 | Low | 0 | 1 |
| Isorhamnetin | 290.27 | 0.85 | No | −2.14 | High | 1 | 0 |
| Kaempferol | 290.27 | 0.85 | No | −2.14 | High | 1 | 0 |
| Quercetin | 270.24 | 2.11 | No | −4.4 | High | 0 | 0 |
| Apigenin | 286.24 | 1.73 | No | −3.82 | High | 0 | 0 |
| Vanillic acid | 302.24 | 1.23 | No | −3.24 | High | 1 | 0 |
| Caffeic acid | 286.24 | 1.58 | No | −3.82 | High | 0 | 0 |
| Catechin | 316.26 | 1.65 | No | −3.94 | High | 0 | 0 |
| Epicatechin | 154.12 | 0.65 | No | −0.6 | High | 0 | 0 |
| Chlorogenic acid | 180.16 | 0.93 | No | −0.71 | High | 1 | 0 |
| Ferulic acid | 354.31 | −0.38 | No | 0.4 | Low | 1 | 1 |
| Luteolin | 138.12 | 1.05 | Yes | −1.17 | High | 0 | 0 |
| p-Coumaric acid | 168.15 | 1.08 | No | −1.32 | High | 0 | 0 |
| p-Hydroxybenzoic acid | 164.16 | 1.26 | Yes | −1.28 | High | 0 | 0 |
| Protocatechuic acid | 194.18 | 1.36 | Yes | −1.42 | High | 0 | 0 |
| L-Rhamnose | 180.16 | −2.26 | No | 2.45 | Low | 0 | 0 |
| D-Glucose | 164.16 | −1.5 | No | 1.9 | Low | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez-González, E.; Sandoval-Ramírez, J.; Flores-Hernández, J.; Carrasco-Carballo, A. Ginkgo biloba: Antioxidant Activity and In Silico Central Nervous System Potential. Curr. Issues Mol. Biol. 2023, 45, 9674-9691. https://doi.org/10.3390/cimb45120604
Suárez-González E, Sandoval-Ramírez J, Flores-Hernández J, Carrasco-Carballo A. Ginkgo biloba: Antioxidant Activity and In Silico Central Nervous System Potential. Current Issues in Molecular Biology. 2023; 45(12):9674-9691. https://doi.org/10.3390/cimb45120604
Chicago/Turabian StyleSuárez-González, Eduardo, Jesús Sandoval-Ramírez, Jorge Flores-Hernández, and Alan Carrasco-Carballo. 2023. "Ginkgo biloba: Antioxidant Activity and In Silico Central Nervous System Potential" Current Issues in Molecular Biology 45, no. 12: 9674-9691. https://doi.org/10.3390/cimb45120604