
Integrated Analysis of Transcriptome and Metabolome Profiles in the Longissimus Dorsi Muscle of Buffalo and Cattle
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Sample Collection
2.2. Meat Quality Determination
2.3. RNA-Seq and Transcriptome Data Analysis
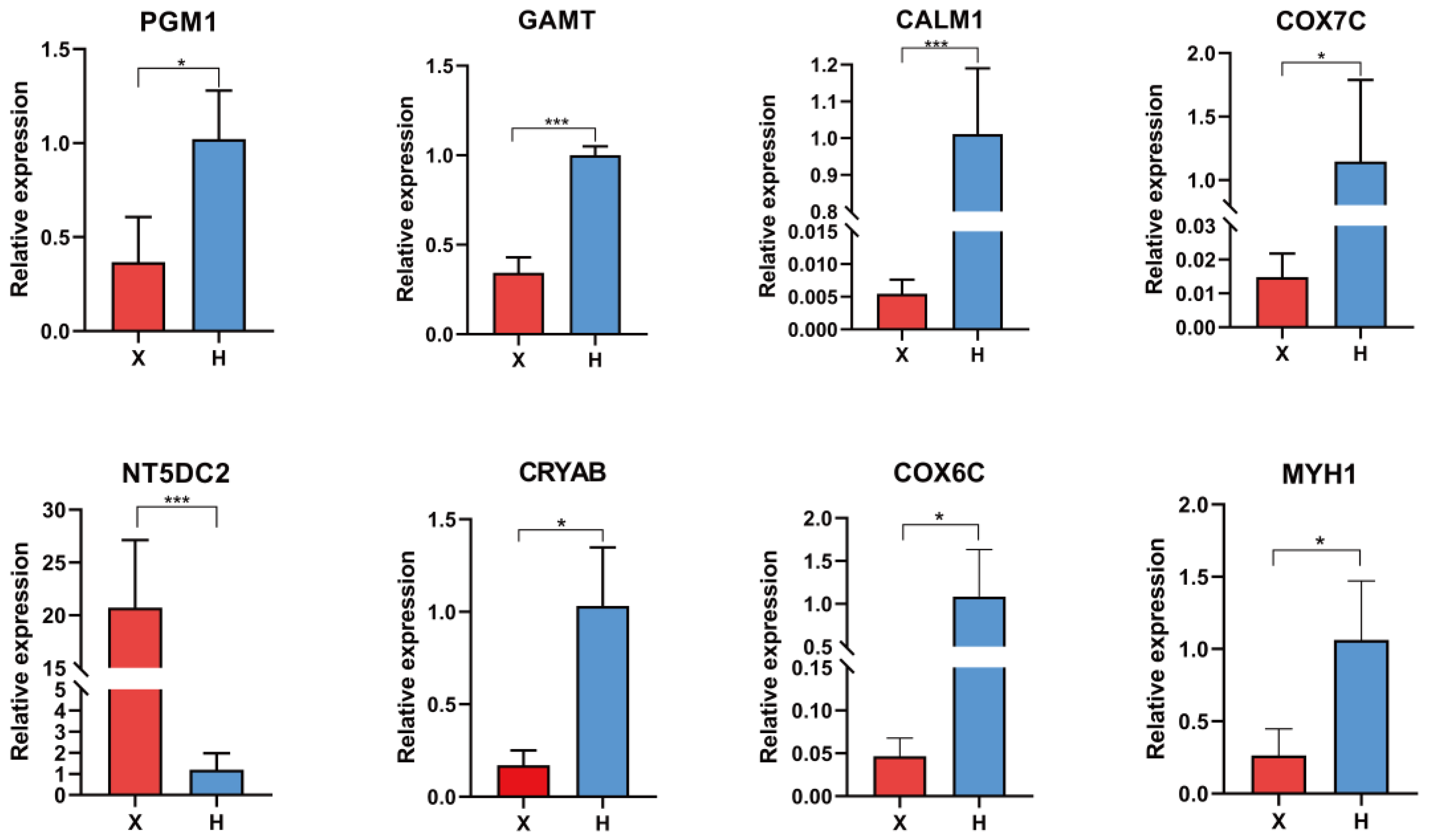
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.5. LC-MS/MS Metabolomics Analysis
2.6. Transcriptome and Metabolome Joint Analysis
2.7. Statistical Analysis
3. Results
3.1. Analysis of Meat Characteristics and Amino Acid Composition
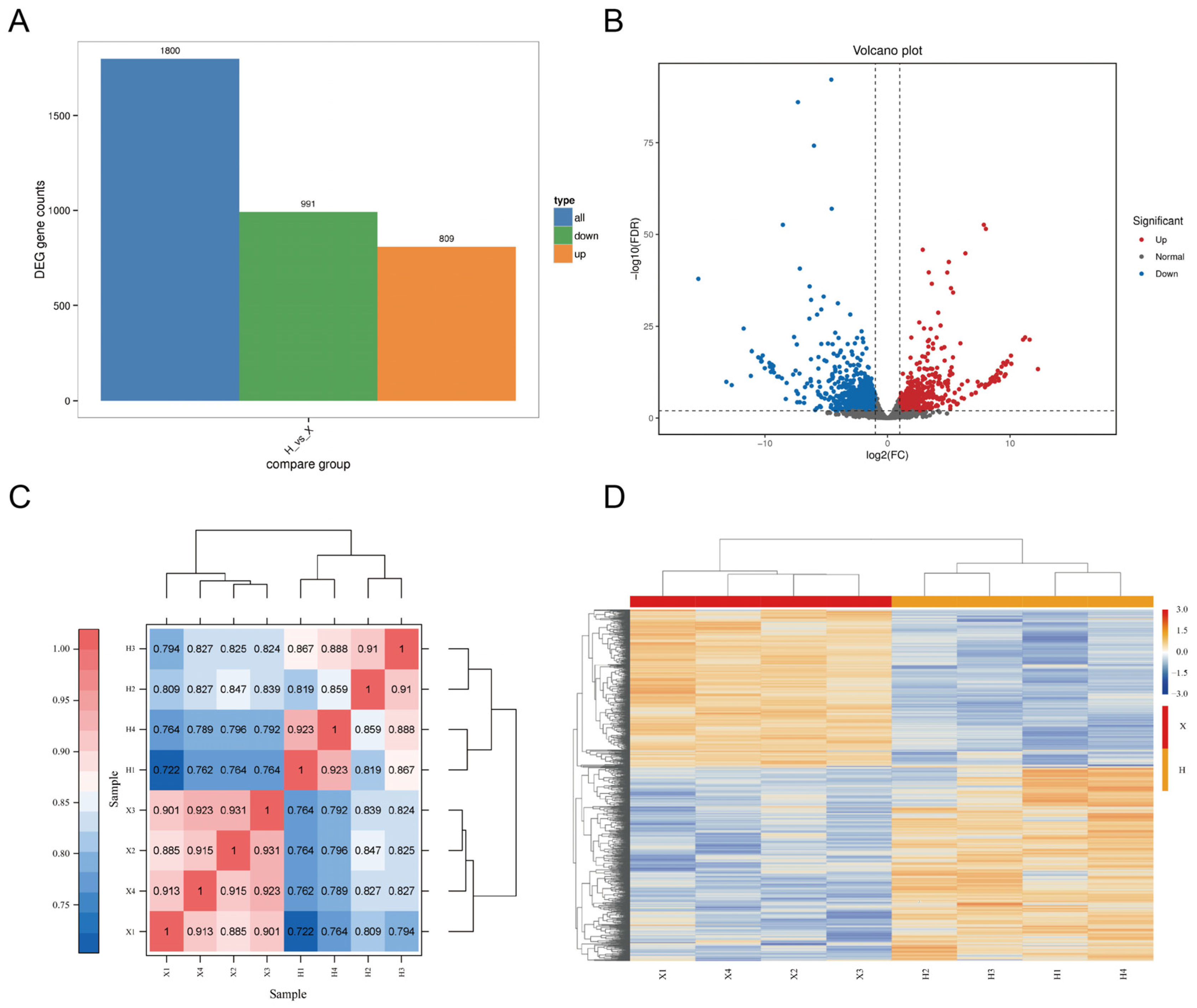
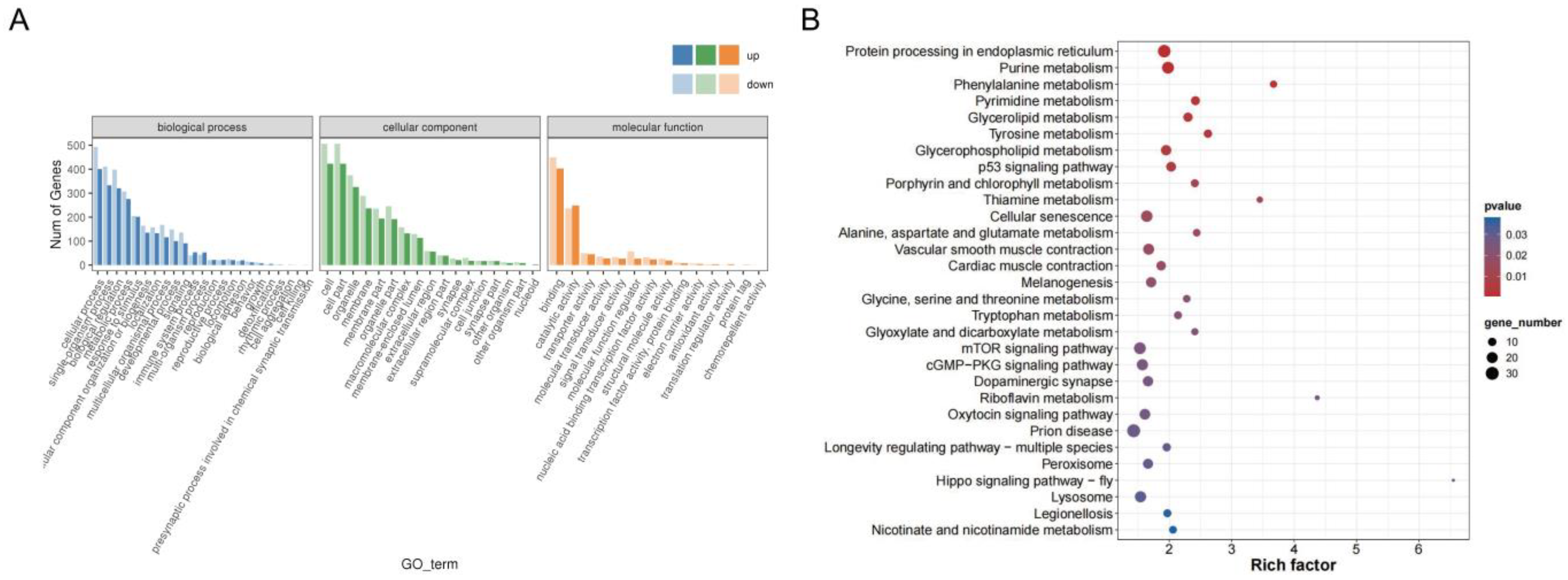
3.2. Analysis of DEGs and Transcriptome Data
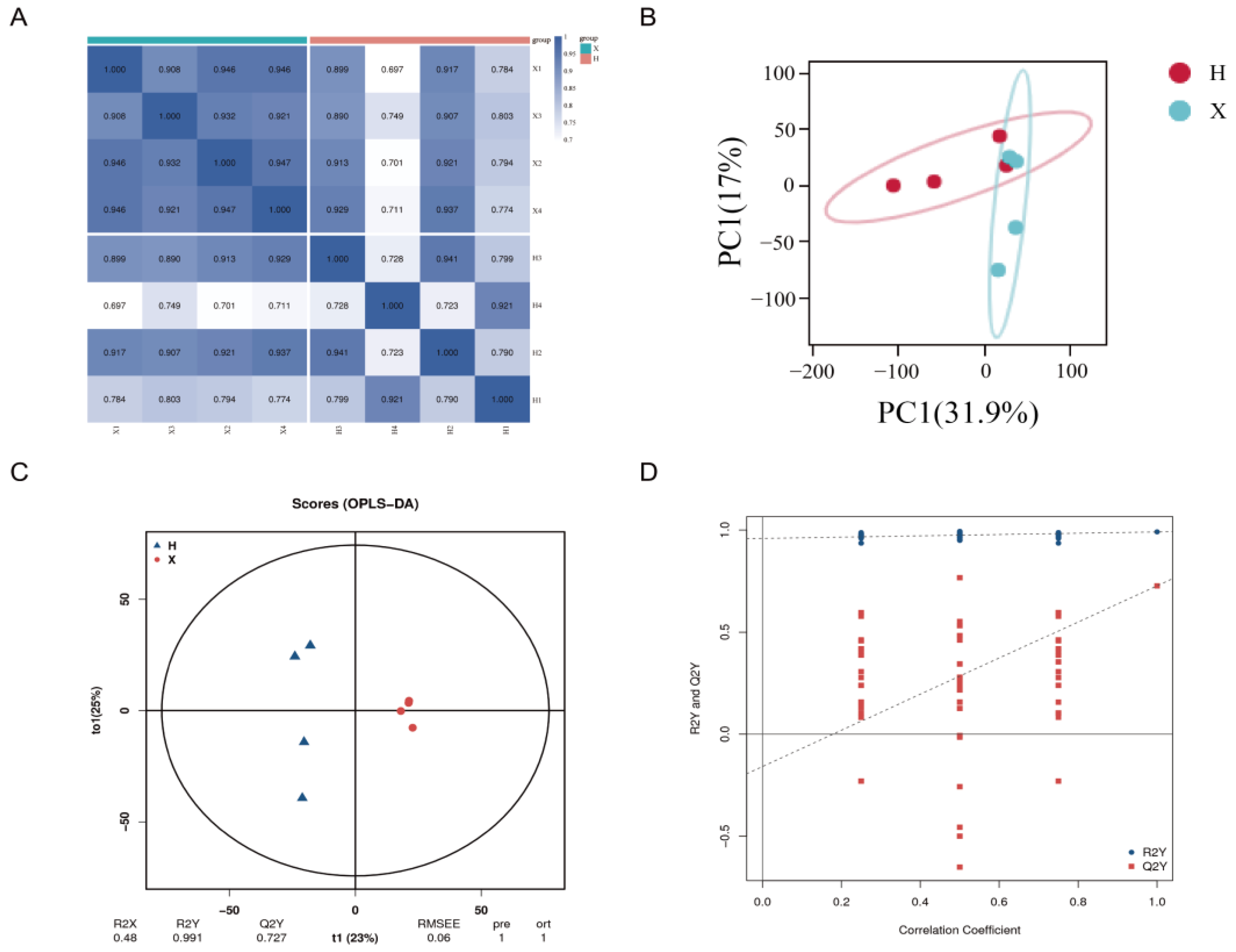
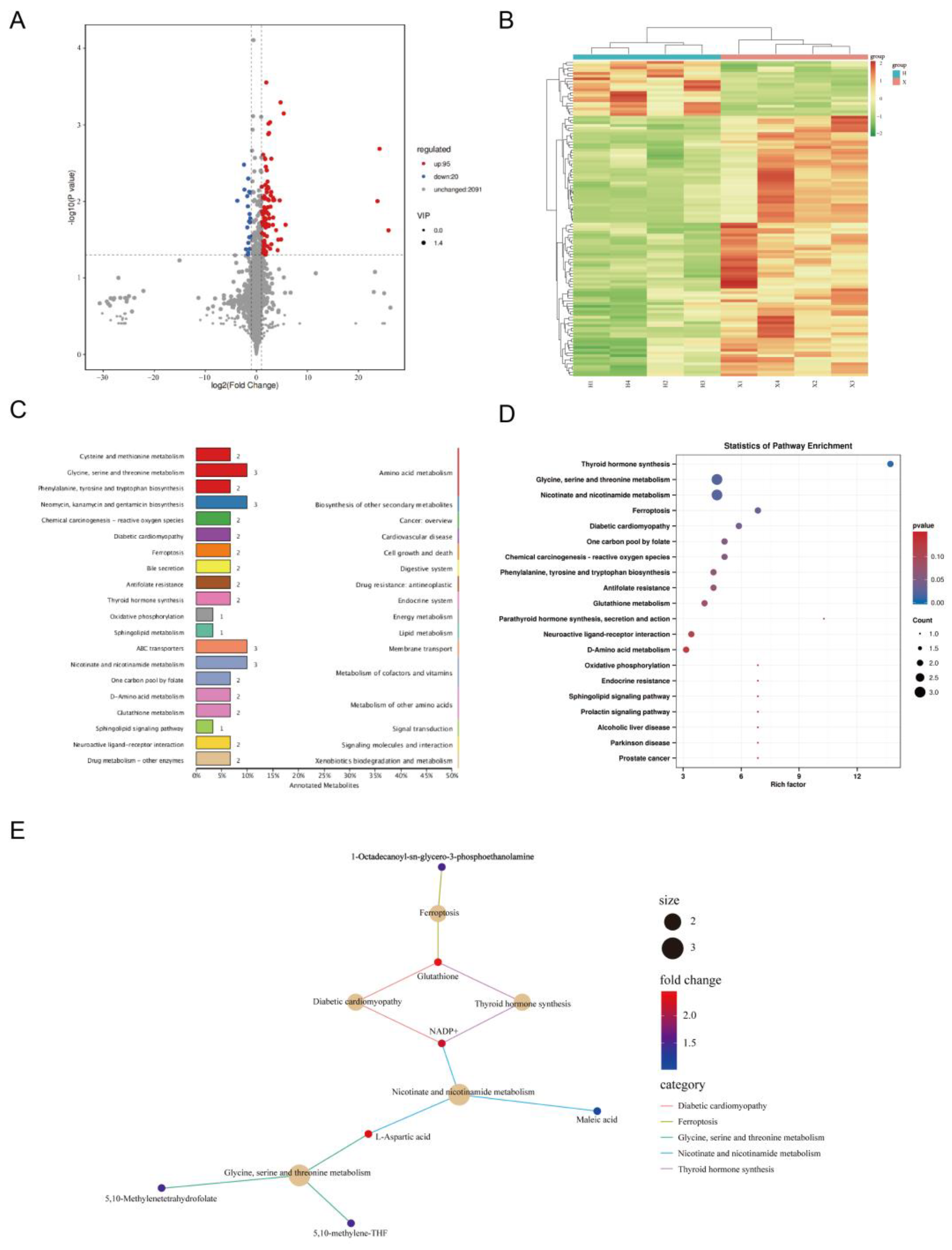
3.3. Analysis of DAMs and Metabolomic Data
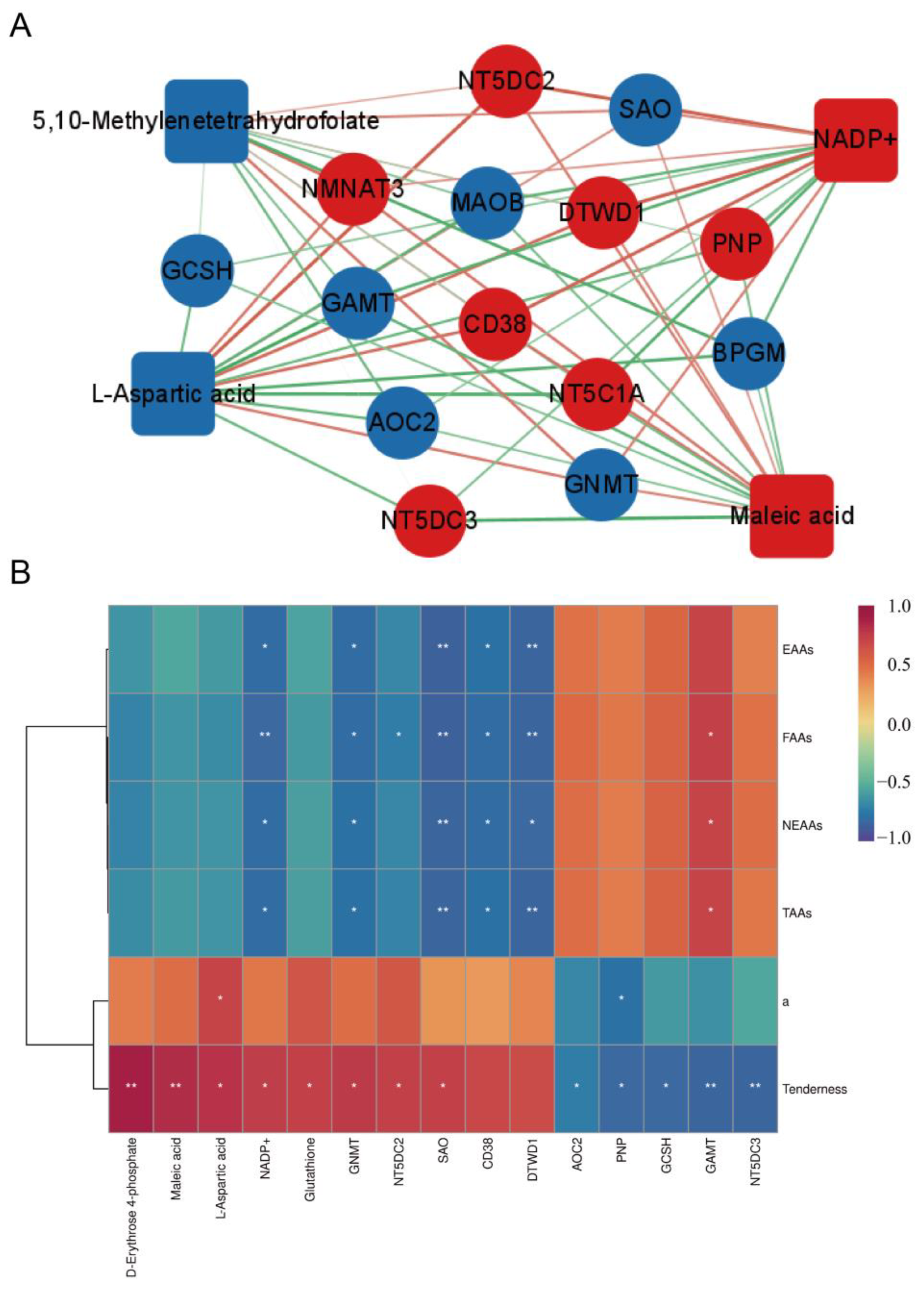
3.4. Joint Analysis of Transcriptome and Metabolome Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karakosta, L.K.; Vatavali, K.A.; Kosma, I.S.; Badeka, A.V.; Kontominas, M.G. Combined Effect of Chitosan Coating and Laurel Essential Oil (Laurus nobilis) on the Microbiological, Chemical, and Sensory Attributes of Water Buffalo Meat. Foods 2022, 11, 1664. [Google Scholar] [CrossRef] [PubMed]
- Di Stasio, L.; Brugiapaglia, A. Current Knowledge on River Buffalo Meat: A Critical Analysis. Animals 2021, 11, 2111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Sun, Z.; Yu, Z.; Li, H.; Luo, H.; Wang, B. Transcriptome and targeted metabolome analysis provide insights into bile acids? new roles and mechanisms on fat deposition and meat quality in lamb. Food Res. Int. 2022, 162, 111941. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, K.; Wang, P.; Feng, T.; Shi, D.; Cui, K.; Luo, C.; Shafique, L.; Qian, Q.; Ruan, J.; et al. Comparison of Long Non-Coding RNA Expression Profiles of Cattle and Buffalo Differing in Muscle Characteristics. Front. Genet. 2020, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Niu, K.; Rushdi, H.E.; Zhang, M.; Fu, T.; Gao, T.; Yang, L.; Liu, S.; Lin, F. Heat Stress Induces Shifts in the Rumen Bacteria and Metabolome of Buffalo. Animals 2022, 12, 1300. [Google Scholar] [CrossRef] [PubMed]
- Kwoji, I.D.; Aiyegoro, O.A.; Okpeku, M.; Adeleke, M.A. ‘Multi-omics’ data integration: Applications in probiotics studies. NPJ Sci. Food 2023, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Han, L.; Hou, S.; Raza, S.H.A.; Wang, Z.; Yang, B.; Sun, S.; Ding, B.; Gui, L.; Simal-Gandara, J.; et al. Effects of different feeding regimes on muscle metabolism and its association with meat quality of Tibetan sheep. Food Chem. 2022, 374, 131611. [Google Scholar] [CrossRef]
- Lu, X.; Arbab, A.A.I.; Zhang, Z.; Fan, Y.; Han, Z.; Gao, Q.; Sun, Y.; Yang, Z. Comparative Transcriptomic Analysis of the Pituitary Gland between Cattle Breeds Differing in Growth: Yunling Cattle and Leiqiong Cattle. Animals 2020, 10, 1271. [Google Scholar] [CrossRef]
- Chen, S.; Chang, C.; Cui, K.; Yang, W.; Li, B.; Ni, S.; Zhang, W.; Li, S.; Li, X.; Wu, G.; et al. Whole-genome analyses reveal the genomic diversity and selection signatures of Hainan cattle. Livest. Sci. 2023, 275, 105311. [Google Scholar] [CrossRef]
- Hoa, V.-B.; Song, D.-H.; Seol, K.-H.; Kang, S.-M.; Kim, H.-W.; Bae, I.-S.; Kim, E.-S.; Park, Y.-S.; Cho, S.-H. A Comparative Study on the Meat Quality, Taste and Aroma Related Compounds between Korean Hanwoo and Chikso Cattle. Foods 2023, 12, 805. [Google Scholar] [CrossRef]
- Wang, Z.; An, X.; Yang, Y.; Zhang, L.; Jiao, T.; Zhao, S. Comprehensive Analysis of the Longissimus Dorsi Transcriptome and Metabolome Reveals the Regulatory Mechanism of Different Varieties of Meat Quality. J. Agric. Food Chem. 2023, 71, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Blecker, C.; Chen, L.; Xiang, C.; Zheng, X.; Wang, Z.; Zhang, D. Integrating identification and targeted proteomics to discover the potential indicators of postmortem lamb meat quality. Meat Sci. 2023, 199, 109126. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Zhang, M.; Zhao, Y.; Wang, R.; Zhang, G.; Lambo, M.T.; Zhang, Y.; Li, Y.; Wang, L. Altering the ratio of palmitic, stearic, and oleic acids in dietary fat affects nutrient digestibility, plasma metabolites, growth performance, carcass, meat quality, and lipid metabolism gene expression of Angus bulls. Meat Sci. 2023, 199, 109138. [Google Scholar] [CrossRef] [PubMed]
- Bostami, A.B.M.R.; Mun, H.-S.; Yang, C.-J. Longissimus dorsi Muscle’s Chemical Composition, Fatty Acid Pattern, and Oxidative Stability in Korean Hanwoo Finishing Cattle Following Slaughtering and Stunning with or without Brain Disruption and State of Consciousness. Foods 2023, 12, 928. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-W.; Su, S.-F.; Zhao, J.; He, X.-L.; Fu, S.-Y.; Wang, B.; Wang, Y.-F.; Wang, D.-Q.; Yun, N.-N.; Chen, X.; et al. Effects of dietary oat supplementation on carcass traits, muscle metabolites, amino acid profiles, and its association with meat quality of Small-tail Han sheep. Food Chem. 2023, 411, 135456. [Google Scholar] [CrossRef]
- VanSlyke, J.K.; Musil, L.S. Analysis of connexin intracellular transport and assembly. Methods 2000, 20, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, T.; Shen, X.; Liu, J.; Zhao, D.; Sun, Y.; Wang, L.; Liu, Y.; Gong, X.; Liu, Y.; et al. Serum metabolomics for early diagnosis of esophageal squamous cell carcinoma by UHPLC-QTOF/MS. Metabolomics 2016, 12, 116. [Google Scholar] [CrossRef]
- Chen, B.; Yue, Y.; Li, J.; Liu, J.; Yuan, C.; Guo, T.; Zhang, D.; Yang, B.; Lu, Z. Transcriptome-metabolome analysis reveals how sires affect meat quality in hybrid sheep populations. Front. Nutr. 2022, 9, 799685. [Google Scholar] [CrossRef]
- Yue, J.; Dong, Y.; Liu, S.; Jia, Y.; Li, C.; Wang, Z.; Gong, S. Integrated Proteomic and Metabolomic Analyses Provide Insights Into Acquisition of Embryogenic Ability in Agapanthus praecox. Front. Plant Sci. 2022, 13, 858065. [Google Scholar] [CrossRef]
- Bertoni, A.; Napolitano, F.; Mota-Rojas, D.; Sabia, E.; Álvarez-Macías, A.; Mora-Medina, P.; Morales-Canela, A.; Berdugo-Gutiérrez, J.; Guerrero-Legarreta, I.J.J.B.S. Similarities and differences between river buffaloes and cattle: Health, physiological, behavioral and productivity aspects. J. Buffalo Sci. 2020, 9, 92–109. [Google Scholar] [CrossRef]
- Guerrero-Legarreta, I.; Napolitano, F.; Cruz-Monterrosa, R.; Mota-Rojas, D.; Mora-Medina, P.; Ramírez-Bribiesca, E.; Bertoni, A.; Berdugo-Gutiérrez, J.; Braghieri, A. River buffalo meat production and quality: Sustainability, productivity, nutritional and sensory properties. J. Buffalo Sci. 2020, 9, 159–169. [Google Scholar] [CrossRef]
- Huang, K.; Chen, M.; Zhong, D.; Luo, X.; Feng, T.; Song, M.; Chen, Y.; Wei, X.; Shi, D.; Liu, Q.; et al. Circular RNA Profiling Reveals an Abundant circEch1 That Promotes Myogenesis and Differentiation of Bovine Skeletal Muscle. J. Agric. Food Chem. 2021, 69, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Coleman, L.W.; Schreurs, N.M.; Kenyon, P.R.; Morris, S.T.; Hickson, R.E. Growth, carcass and meat quality characteristics of Charolais-sired steers and heifers born to Angus-cross-dairy and Angus breeding cows. Meat Sci. 2023, 201, 109178. [Google Scholar] [CrossRef] [PubMed]
- Spanghero, M.; Gracco, L.; Valusso, R.; Piasentier, E.J.L.P.S. In vivo performance, slaughtering traits and meat quality of bovine (Italian Simmental) and buffalo (Italian Mediterranean) bulls. Livest. Prod. Sci. 2004, 91, 129–141. [Google Scholar] [CrossRef]
- Lapitan, R.M.; Del Barrio, A.N.; Katsube, O.; Ban-Tokuda, T.; Orden, E.A.; Robles, A.Y.; Fujihara, T.; Cruz, L.C.; Homma, H.; Kanai, Y. Comparison of carcass and meat characteristics of Brahman grade cattle (Bos indicus) and crossbred water buffalo (Bubalus bubalis). Anim. Sci. J. 2007, 78, 596–604. [Google Scholar] [CrossRef]
- Zhan, H.; Xiong, Y.; Wang, Z.; Dong, W.; Zhou, Q.; Xie, S.; Li, X.; Zhao, S.; Ma, Y. Integrative analysis of transcriptomic and metabolomic profiles reveal the complex molecular regulatory network of meat quality in Enshi black pigs. Meat Sci. 2022, 183, 108642. [Google Scholar] [CrossRef]
- Kwak, J.Y.; Ham, H.J.; Kim, C.M.; Hwang, E.S. Nicotinamide Exerts Antioxidative Effects on Senescent Cells. Mol. Cells 2015, 38, 229–235. [Google Scholar] [CrossRef]
- Hughes, J.; Clarke, F.; Purslow, P.; Warner, R. High pH in beef longissimus thoracis reduces muscle fibre transverse shrinkage and light scattering which contributes to the dark colour. Food Res. Int. 2017, 101, 228–238. [Google Scholar] [CrossRef]
- Khatri, Y.; Huff-Lonergan, E. Effect of postmortem pH on the physical, biochemical, and sensory characteristics of bovine longissimus thoracis et lumborum muscle. Food Sci. Nutr. 2023, 11, 2002–2011. [Google Scholar] [CrossRef]
- Marrone, R.; Salzano, A.; Di Francia, A.; Vollano, L.; Di Matteo, R.; Balestrieri, A.; Anastasio, A.; Barone, C.M.A. Effects of Feeding and Maturation System on Qualitative Characteristics of Buffalo Meat (Bubalus bubalis). Animals 2020, 10, 899. [Google Scholar] [CrossRef]
- Lonergan, E.H.; Zhang, W.; Lonergan, S.M. Biochemistry of postmortem muscle—Lessons on mechanisms of meat tenderization. Meat Sci. 2010, 86, 184–195. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Posey, E.A.; Steele, C.C.; Savell, J.W.; Bazer, F.W.; Wu, G. Dietary glycine supplementation enhances postweaning growth and meat quality of pigs with intrauterine growth restriction. J. Anim. Sci. 2023, 101, 354. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, W.; Zhang, C.; Li, F.; Yang, H.; Wang, Z. Metabolomic comparison of meat quality and metabolites of geese breast muscle at different ages. Food Chem. X 2023, 19, 100775. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Han, L.; Gui, L.; Raza, S.H.A.; Hou, S.; Yang, B.; Wang, Z.; Ma, Y.; Makhlof, R.T.M.; Alhuwaymil, Z.; et al. Metabolome and microbiome analysis revealed the effect mechanism of different feeding modes on the meat quality of Black Tibetan sheep. Front. Microbiol. 2023, 13, 1076675. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, Y.; Chen, L.; Zhang, Y.; Wang, J.; Li, H.; Yan, X.; Xia, L.; Yao, G. Differences in meat quality between Angus cattle and Xinjiang brown cattle in association with gut microbiota and its lipid metabolism. Front. Microbiol. 2022, 13, 988984. [Google Scholar] [CrossRef]
- Barcelos, R.P.; Stefanello, S.T.; Mauriz, J.L.; Gonzalez-Gallego, J.; Soares, F.A.A. Creatine and the Liver: Metabolism and Possible Interactions. Mini-Rev. Med. Chem. 2016, 16, 12–18. [Google Scholar] [CrossRef]
- Duan, B.B.; Xu, J.W.; Xing, T.; Li, J.L.; Zhang, L.; Gao, F. Creatine nitrate supplementation strengthens energy status and delays glycolysis of broiler muscle via inhibition of LKB1/AMPK pathway. Poult. Sci. 2022, 101, 101653. [Google Scholar] [CrossRef]
- Su, Y.; Sun, X.; Zhao, S.; Hu, M.; Li, D.; Qi, S.; Jiao, X.; Sun, Y.; Wang, C.; Zhu, X.; et al. Dietary alfalfa powder supplementation improves growth and development, body health, and meat quality of Tibetan sheep. Food Chem. 2022, 396, 133709. [Google Scholar] [CrossRef]
- Huang, Q.; Dong, K.; Wang, Q.; Huang, X.; Wang, G.; An, F.; Luo, Z.; Luo, P. Changes in volatile flavor of yak meat during oxidation based on multi-omics. Food Chem. 2022, 371, 131103. [Google Scholar] [CrossRef]
- Watkins, P.J.; Frank, D.; Singh, T.K.; Young, O.A.; Warner, R.D. Sheepmeat Flavor and the Effect of Different Feeding Systems: A Review. J. Agric. Food Chem. 2013, 61, 3561–3579. [Google Scholar] [CrossRef]
- Madeira, M.S.; Alfaia, C.M.; Costa, P.; Lopes, P.A.; Martins, S.V.; Lemos, J.P.C.; Moreira, O.; Santos-Silva, J.; Bessa, R.J.B.; Prates, J.A.M. Effect of betaine and arginine in lysine-deficient diets on growth, carcass traits, and pork quality. J. Anim. Sci. 2015, 93, 4721–4733. [Google Scholar] [CrossRef] [PubMed]
- Chanalia, P.; Gandhi, D.; Attri, P.; Dhanda, S. Extraction, purification and characterization of low molecular weight Proline iminopeptidase from probiotic L. plantarum for meat tenderization. Int. J. Biol. Macromol. 2018, 109, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Vidal, V.A.S.; Santana, J.B.; Paglarini, C.S.; da Silva, M.A.A.P.; Freitas, M.Q.; Esmerino, E.A.; Cruz, A.G.; Pollonio, M.A.R. Adding lysine and yeast extract improves sensory properties of low sodium salted meat. Meat Sci. 2020, 159, 107911. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, I.H. The effects of betaine supplementation in diets containing different levels of crude protein and methionine on the growth performance, blood components, total tract nutrient digestibility, excreta noxious gas emission, and meat quality of the broiler chickens. Poult. Sci. 2019, 98, 6808–6815. [Google Scholar]
- Roobab, U.; Zeng, X.-A.; Ahmed, W.; Madni, G.M.; Manzoor, M.F.; Aadil, R.M. Effect of Pulsed Electric Field on the Chicken Meat Quality and Taste-Related Amino Acid Stability: Flavor Simulation. Foods 2023, 12, 710. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | H | X | p-Value | ||
|---|---|---|---|---|---|
| Meat quality characteristics | pH | 45 min | 6.22 ± 0.13 | 6.21 ± 0.18 | 0.918 |
| 24 h | 5.29 ± 0.31 | 5.14 ± 0.08 | 0.229 | ||
| Peat color | L | 30.59 ± 1.21 | 30.11 ± 1.60 | 0.536 | |
| a | 12.77 ± 1.12 a | 14.14 ± 1.05 b | 0.034 | ||
| b | 5.55 ± 1.13 | 5.23 ± 0.95 | 0.569 | ||
| Shear force (N) | 58.14 ± 16.14 a | 76.77 ± 7.47 b | 0.015 | ||
| Drip loss (%) | 3.77 ± 1.06 | 4.77 ± 1.20 | 0.121 | ||
| Moisture (%) | 70.59 ± 2.68 | 73.69 ± 0.91 | 0.108 | ||
| Ash (%) | 4.33 ± 0.03 | 4.52 ± 0.09 | 0.206 | ||
| Crude fat (%) | 3.31 ± 1.44 | 2.90 ± 0.66 | 0.667 | ||
| Crude protein (%) | 25.89 ± 2.32 | 23.00 ± 0.68 | 0.083 | ||
| Amino acids | TAA (g/100 g) | 16.09 ± 0.74 a | 13.63 ± 1.39 b | 0.035 | |
| EAA (g/100 g) | 6.50 ± 0.29 a | 5.44 ± 0.62 b | 0.036 | ||
| NEAA (g/100 g) | 9.59 ± 0.46 a | 8.19 ± 0.77 b | 0.036 | ||
| FAA (g/100 g) | 6.76 ± 0.32 a | 5.71 ± 0.52 b | 0.026 |
| Amino Acids (mg/100 g) | H | X | p-Value |
|---|---|---|---|
| Histidine | 703.28 ± 26.78 | 618.66 ± 85.59 | 0.153 |
| 4-Hydroxy-L-Proline | 62.96 ± 13.48 | 53.77 ± 4.68 | 0.307 |
| Arginine | 1035.82 ± 46.33 a | 863.49 ± 91.08 b | 0.027 |
| Asparagine | 0.18 ± 0.09 | 0.12 ± 0.03 | 0.327 |
| Glutamine | 0.97 ± 0.45 | 0.53 ± 0.06 | 0.145 |
| Serine | 652.55 ± 41.72 | 560.99 ± 56.45 | 0.065 |
| Glycine | 935.51 ± 50.60 a | 813.65 ± 54.36 b | 0.029 |
| Aspartic acid | 1612.44 ± 78.86 a | 1346.59 ± 129.67 b | 0.023 |
| Glutamic acid | 1913.71 ± 94.63 a | 1600.13 ± 146.62 b | 0.021 |
| Threonine | 845.14 ± 39.25 a | 722.31 ± 71.07 b | 0.040 |
| Alanine | 1258.15 ± 73.11 | 1084.39 ± 104.57 | 0.056 |
| Gamma-Aminobutyric acid | 1.44 ± 0.35 | 1.07 ± 0.15 | 0.148 |
| Proline | 892.72 ± 55.23 a | 763.97 ± 71.79 b | 0.049 |
| D-2-Aminobutyric acid | 0.24 ± 0.15 | 0.25 ± 0.09 | 0.932 |
| Lysine | 1409.18 ± 103.03 a | 1114.90 ± 178.08 b | 0.048 |
| Cystine | 21.16 ± 2.89 | 16.33 ± 2.30 | 0.064 |
| Methionine | 391.74 ± 24.23 a | 325.51 ± 30.01 b | 0.025 |
| Tyrosine | 667.48 ± 32.20 a | 545.68 ± 57.27 b | 0.018 |
| Valine | 711.07 ± 23.14 | 614.74 ± 66.46 | 0.055 |
| Isoleucine | 732.46 ± 17.87 | 639.48 ± 69.92 | 0.067 |
| Leucine | 1503.17 ± 61.82 | 1309.57 ± 139.75 | 0.071 |
| Phenylalanine | 737.14 ± 30.88 a | 631.02 ± 68.46 b | 0.050 |
| Tryptophan | 6.40 ± 5.11 | 0.90 ± 0.09 | 0.112 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, G.; Qiu, X.; Jiao, Z.; Yang, W.; Pan, H.; Li, H.; Bian, Z.; Geng, Q.; Wu, H.; Jiang, J.; et al. Integrated Analysis of Transcriptome and Metabolome Profiles in the Longissimus Dorsi Muscle of Buffalo and Cattle. Curr. Issues Mol. Biol. 2023, 45, 9723-9736. https://doi.org/10.3390/cimb45120607
Wu G, Qiu X, Jiao Z, Yang W, Pan H, Li H, Bian Z, Geng Q, Wu H, Jiang J, et al. Integrated Analysis of Transcriptome and Metabolome Profiles in the Longissimus Dorsi Muscle of Buffalo and Cattle. Current Issues in Molecular Biology. 2023; 45(12):9723-9736. https://doi.org/10.3390/cimb45120607
Chicago/Turabian StyleWu, Guansheng, Xinjun Qiu, Zizhuo Jiao, Weijie Yang, Haoju Pan, Hong Li, Zhengyu Bian, Qiang Geng, Hui Wu, Junming Jiang, and et al. 2023. "Integrated Analysis of Transcriptome and Metabolome Profiles in the Longissimus Dorsi Muscle of Buffalo and Cattle" Current Issues in Molecular Biology 45, no. 12: 9723-9736. https://doi.org/10.3390/cimb45120607