
Hemin with Peroxidase Activity Can Inhibit the Oxidative Damage Induced by Ultraviolet A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Measurement of Fluorescence
2.3. Antioxidant Activity Assay
2.3.1. Superoxide Anion Free-Radical-Scavenging Assay
2.3.2. DPPH· Scavenging Assay
2.3.3. Hydroxyl Radical Scavenging
2.4. Cell Culture and UVA Treatment
2.5. Cell Viability Assay
2.6. Measurement of LDH and MDA Release
2.7. Annexin-V–Propidium Iodide Assay
2.8. Cell-Cycle Analysis
2.9. Statistical Analysis
3. Results
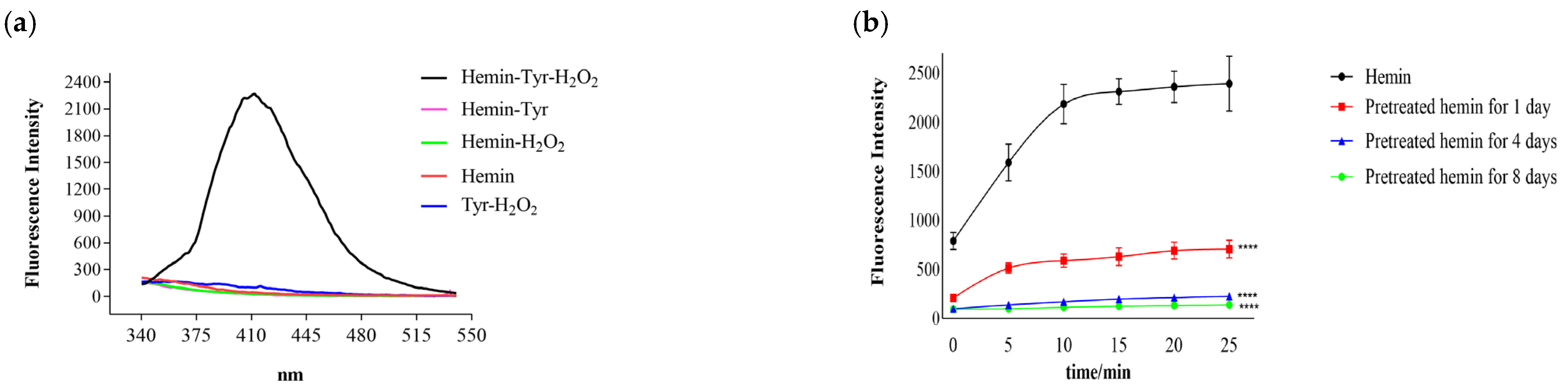
3.1. Catalysis of Hemin
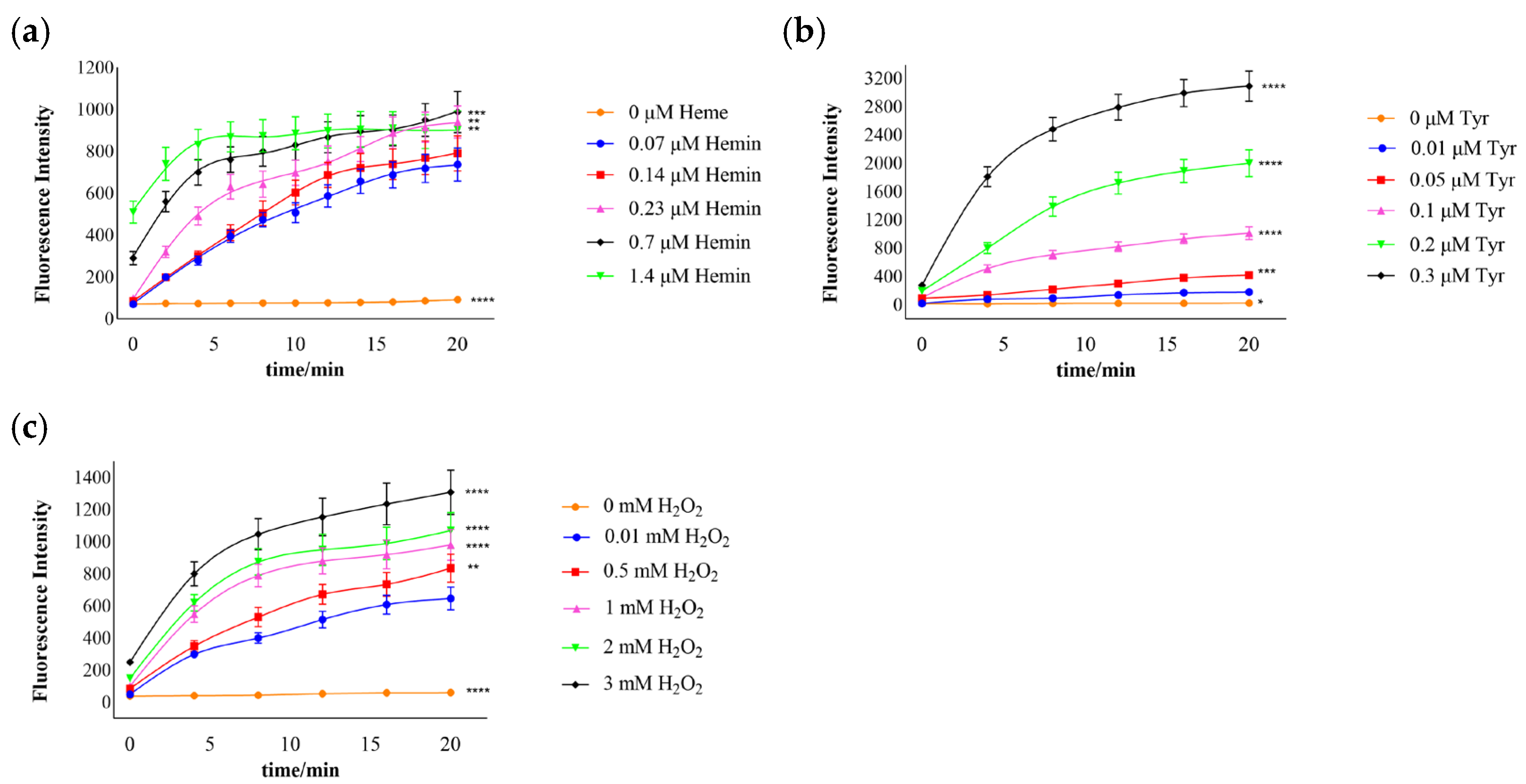
3.2. Effects of Hemin and Substrate Concentration on the Catalytic Reaction
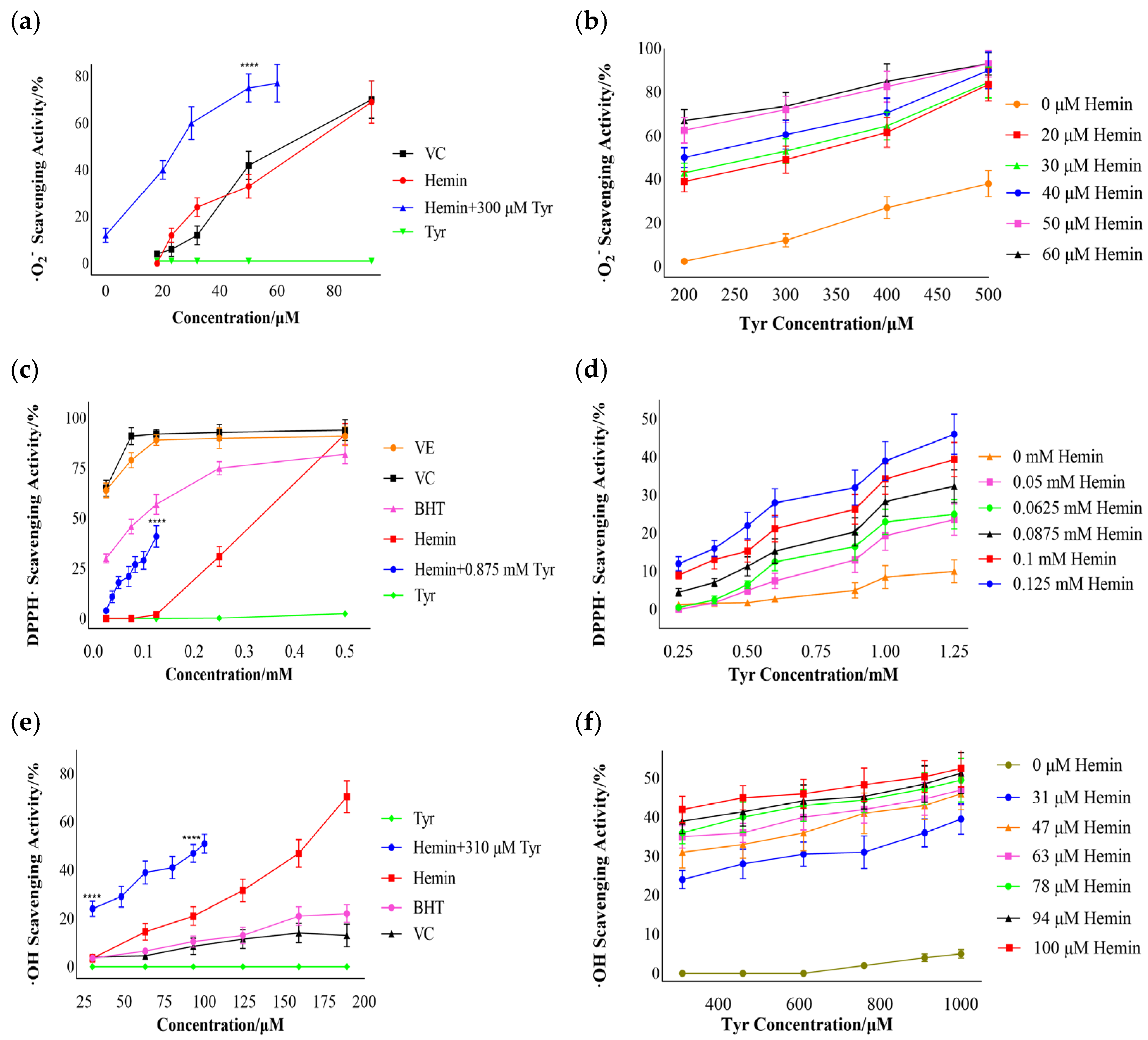
3.3. Free-Radical Scavenging by Hemin
3.3.1. Superoxide Anion Free-Radical Scavenging
3.3.2. DPPH·Scavenging
3.3.3. Hydroxyl Free-Radical Scavenging
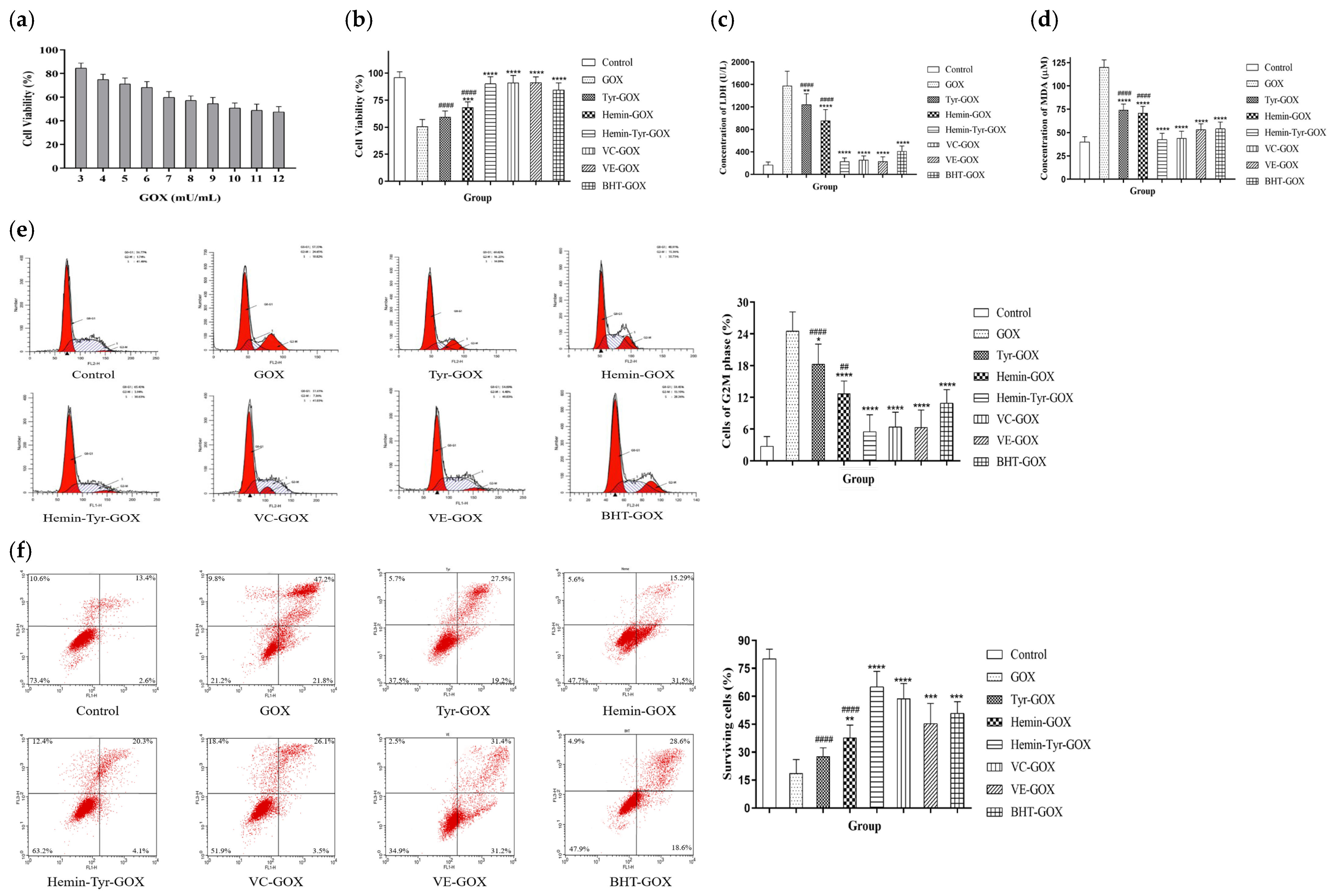
3.4. Protection by Hemin Catalysis against the Oxidative Damage of Cells Induced by GOX
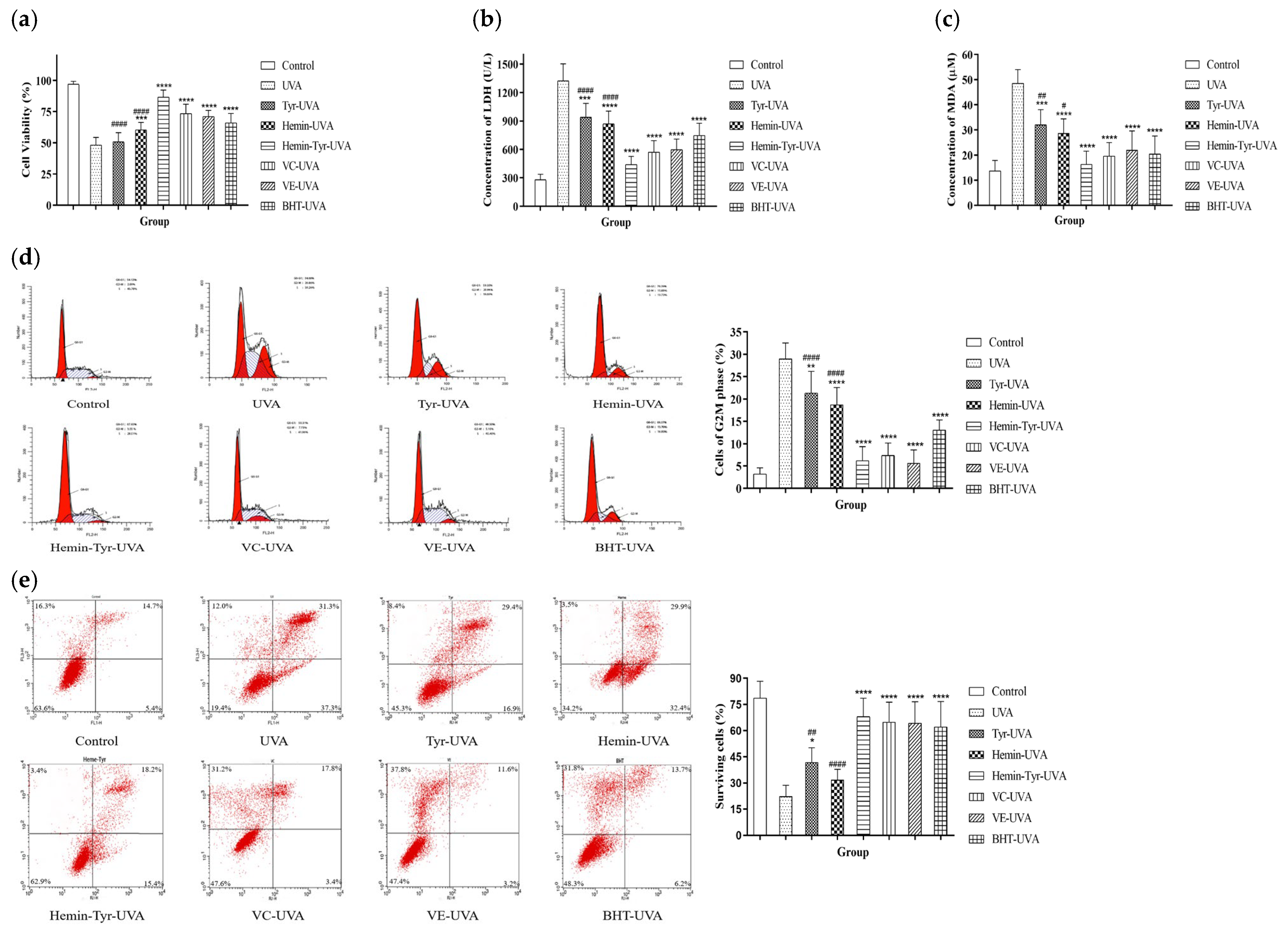
3.5. Effects of Hemin and Tyrosine on UVA-Induced Photodamage in HaCaT Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New insights in photoaging, UVA induced damage and skin types. Exp. Dermatol. 2014, 23 (Suppl. S1), 7–12. [Google Scholar] [CrossRef] [PubMed]
- Souza de Carvalho, V.M.; Covre, J.L.; Correia-Silva, R.D.; Lice, I.; Corrêa, M.P.; Leopoldino, A.M.; Gil, C.D. Bellis perennis extract mitigates UVA-induced keratinocyte damage: Photoprotective and immunomodulatory effects. J. Photochem. Photobiol. B 2021, 221, 112247. [Google Scholar] [CrossRef] [PubMed]
- Lone, A.N.; Malik, A.T.; Naikoo, H.S.; Raghu, R.S.; Tasduq, S.A. Trigonelline, a naturally occurring alkaloidal agent protects ultraviolet-B (UV-B) irradiation induced apoptotic cell death in human skin fibroblasts via attenuation of oxidative stress, restoration of cellular calcium homeostasis and prevention of endoplasmic reticulum (ER) stress. J. Photochem. Photobiol. B 2020, 202, 111720. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Q.; Wang, W.; Su, L.; Han, Y.; Wang, C. Molecular mechanism of polypeptides from Chlamys farreri (PCF)'s anti-apoptotic effect in UVA-exposed HaCaT cells involves HSF1/HSP70, JNK, XO, iNOS and NO/ROS. J. Photochem. Photobiol. B 2014, 130, 47–56. [Google Scholar] [CrossRef]
- De Gruijl, F.R. Photocarcinogenesis: UVA vs. UVB radiation. Ski. Pharmacol. Physiol. 2002, 15, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, P.; Xue, H.; Li, Q. Cyanidin-3-glucoside prevents hydrogen peroxide (H(2)O(2))-induced oxidative damage in HepG2 cells. Biotechnol. Lett. 2020, 42, 2453–2466. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated damage to cellular DNA by UVB and UVA radiation. Photochem. Photobiol. 2015, 91, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Geng, R.; Lu, Q.; Tan, X.; Rao, R.; Zhou, H.; Yang, X.; Liu, W. Involvement of TGF-β and ROS in G1 Cell Cycle Arrest Induced by Titanium Dioxide Nanoparticles Under UVA Irradiation in a 3D Spheroid Model. Int. J. Nanomed. 2020, 15, 1997–2010. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.Q.; Travers, J.B.; Kemp, M.G. Roles of UVA radiation and DNA damage responses in melanoma pathogenesis. Environ. Mol. Mutagen. 2018, 59, 438–460. [Google Scholar] [CrossRef]
- Wu, S.; Hu, Y.; Li, Z.; Bai, W.; Zhao, J.; Huang, C.; Li, Q.; Fan, C.; Deng, L.; Lu, D. The effect of Cyanidin-3-o-glucoside on UVA-induced damage in human dermal fibroblasts. Photodermatol. Photoimmunol. Photomed. 2018, 34, 224–231. [Google Scholar] [CrossRef]
- Hoppe, M.; Brün, B.; Larsson, M.P.; Moraeus, L.; Hulthén, L. Heme iron-based dietary intervention for improvement of iron status in young women. Nutrition 2013, 29, 89–95. [Google Scholar] [CrossRef]
- Quintero-Gutiérrez, A.G.; González-Rosendo, G.; Pozo, J.P.; Villanueva-Sánchez, J. Heme Iron Concentrate and Iron Sulfate Added to Chocolate Biscuits: Effects on Hematological Indices of Mexican Schoolchildren. J. Am. Coll. Nutr. 2016, 35, 544–551. [Google Scholar] [CrossRef]
- Gamage, S.M.K.; Dissabandara, L.; Lam, A.K.; Gopalan, V. The role of heme iron molecules derived from red and processed meat in the pathogenesis of colorectal carcinoma. Crit. Rev. Oncol. 2018, 126, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Shimzu, K.; Takahashi, T.; Iwasaki, T.; Shimizu, H.; Inoue, K.; Morimatsu, H.; Omori, E.; Matsumi, M.; Akagi, R.; Morita, K. Hemin treatment abrogates monocrotaline-induced pulmonary hypertension. Med. Chem. 2008, 4, 572–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Jena, G.R.; Ram, M.; Lingaraju, M.C.; Singh, V.; Prasad, R.; Kumawat, S.; Kant, V.; Gupta, P.; Tandan, S.K.; et al. Hemin attenuated oxidative stress and inflammation to improve wound healing in diabetic rats. Naunyn Schmiedebergs Arch. Pharm. 2019, 392, 1435–1445. [Google Scholar] [CrossRef]
- Huo, S.; Lei, X.; He, D.; Zhang, H.; Yang, Z.; Mu, W.; Fang, K.; Xue, D.; Li, H.; Li, X.; et al. Ferrous hemoglobin and hemoglobin-based oxygen carriers acting as a peroxidase can inhibit oxidative damage to endothelial cells caused by hydrogen peroxide. Artif. Organs 2021, 45, 1229–1239. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Sharma, S.K.; Singh, A.P. In vitro antioxidant and free radical scavenging activity of Nardostachys jatamansi DC. J. Acupunct. Meridian Stud. 2012, 5, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.E.; Lin, L.H. DPPH scavenging capacity of extracts from Camellia seed dregs using polyol compounds as solvents. Heliyon 2019, 5, e02315. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Lin, Y.; Li, Q.; Gu, Y. The contribution ratio of various characteristic tea compounds in antioxidant capacity by DPPH assay. J. Food Biochem. 2020, 44, e13270. [Google Scholar] [CrossRef]
- Hoang, A.; Xuan, T.D.; Dieu Thuy, N.T.; Quan, N.V.; Trang, L.T. Antioxidant and a-amylase Inhibitory Activities and Phytocompounds of Clausena indica Fruits. Medicines 2020, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Annibal, A.; Colombo, G.; Milzani, A.; Dalle-Donne, I.; Fedorova, M.; Hoffmann, R. Identification of dityrosine cross-linked sites in oxidized human serum albumin. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1019, 147–155. [Google Scholar] [CrossRef]
- Nagababu, E.; Rifkind, J.M. Formation of fluorescent heme degradation products during the oxidation of hemoglobin by hydrogen peroxide. Biochem. Biophys. Res. Commun. 1998, 247, 592–596. [Google Scholar] [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Role of mitochondrial ROS in the brain: From physiology to neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef]
- Tang, B.; Wang, Y.; Liang, H.; Chen, Z.; He, X.; Shen, H. Studies on the oxidation reaction of tyrosine (Tyr) with H2O2 catalyzed by horseradish peroxidase (HRP) in alcohol-water medium by spectrofluorimetry and differential spectrophotometry. Spectrochim. Acta A Mol. Biomol. 2006, 63, 609–613. [Google Scholar] [CrossRef]
- Atoji, T.; Yatami, H.; Aihara, M.; Takeoka, S. Enzymatic elimination of hydrogen peroxide by a methemoglobin/L-Tyrosine system. Artif. Cells Blood Substit. Immobil. Biotechnol. 2007, 35, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Bonagura, C.A.; Bhaskar, B.; Shimizu, H.; Li, H.; Sundaramoorthy, M.; McRee, D.E.; Goodin, D.B.; Poulos, T.L. High-resolution crystal structures and spectroscopy of native and compound I cytochrome c peroxidase. Biochemistry 2003, 42, 5600–5608. [Google Scholar] [CrossRef]
- Cai, Y.Z.; Mei, S.; Jie, X.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationships of phenolic compounds from traditional Chinese medicinal plants. Life Sci. 2006, 78, 2872–2888. [Google Scholar] [CrossRef]
- Di Lisa, F.; Carpi, A.; Giorgio, V.; Bernardi, P. The mitochondrial permeability transition pore and cyclophilin D in cardioprotection. Biochim. Biophys. Acta 2011, 1813, 1316–1322. [Google Scholar] [CrossRef] [Green Version]
- Bayo Jimenez, M.T.; Frenis, K.; Kröller-Schön, S.; Kuntic, M.; Stamm, P.; Kvandová, M.; Oelze, M.; Li, H.; Steven, S.; Münzel, T.; et al. Noise-Induced Vascular Dysfunction, Oxidative Stress, and Inflammation Are Improved by Pharmacological Modulation of the NRF2/HO-1 Axis. Antioxidants 2021, 10, 625. [Google Scholar] [CrossRef]
- Ahn, Y.; Kim, M.G.; Jo, K.; Hong, K.B.; Suh, H.J. Effects of Sphingomyelin-Containing Milk Phospholipids on Skin Hydration in UVB-Exposed Hairless Mice. Molecules 2022, 27, 2545. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Hseu, Y.C.; Gowrisankar, Y.V.; Chung, Y.T.; Zhang, Y.Z.; Way, T.D.; Yang, H.L. The anti-melanogenic effects of 3-O-ethyl ascorbic acid via Nrf2-mediated α-MSH in-hibition in UVA-irradiated keratinocytes and autophagy induction in melanocytes. Free Radic. Biol. Med. 2021, 173, 151–169. [Google Scholar] [CrossRef]
- Lan, C.E.; Hung, Y.T.; Fang, A.H.; Ching-Shuang, W. Effects of irradiance on UVA-induced skin aging. J. Dermatol. Sci. 2019, 94, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, S.; Braunberger, T.; Mohammad, T.F.; Kohli, I.; Hamzavi, I.H. The Role of Sunscreen in Melasma and Postinflammatory Hyperpigmentation. Indian J. Derm. 2020, 65, 5–10. [Google Scholar] [CrossRef]
- Saguie, B.O.; Martins, R.L.; Fonseca, A.S.D.; Romana-Souza, B.; Monte-Alto-Costa, A. An ex vivo model of human skin photoaging induced by UVA radiation compatible with summer exposure in Brazil. J. Photochem. Photobiol. B 2021, 221, 112255. [Google Scholar] [CrossRef] [PubMed]
- Bugger, H.; Pfeil, K. Mitochondrial ROS in myocardial ischemia reperfusion and remodeling. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165768. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hui, W.; Yang, Z.; Fang, K.; Wu, M.; Mu, W.; Zhao, C.; Xue, D.; Zhu, T.; Li, X.; Gao, M.; et al. Hemin with Peroxidase Activity Can Inhibit the Oxidative Damage Induced by Ultraviolet A. Curr. Issues Mol. Biol. 2022, 44, 2683-2694. https://doi.org/10.3390/cimb44060183
Hui W, Yang Z, Fang K, Wu M, Mu W, Zhao C, Xue D, Zhu T, Li X, Gao M, et al. Hemin with Peroxidase Activity Can Inhibit the Oxidative Damage Induced by Ultraviolet A. Current Issues in Molecular Biology. 2022; 44(6):2683-2694. https://doi.org/10.3390/cimb44060183
Chicago/Turabian StyleHui, Wenli, Zhipeng Yang, Ke Fang, Mengdi Wu, Wenhua Mu, Cong Zhao, Dan Xue, Tengteng Zhu, Xiao Li, Ming Gao, and et al. 2022. "Hemin with Peroxidase Activity Can Inhibit the Oxidative Damage Induced by Ultraviolet A" Current Issues in Molecular Biology 44, no. 6: 2683-2694. https://doi.org/10.3390/cimb44060183