
Construction and Characterization of T7 Bacteriophages Harboring Apidaecin-Derived Sequences


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Peptide Synthesis
2.3. Antibacterial Activity
2.4. Phage-Peptide-Synergy Assay
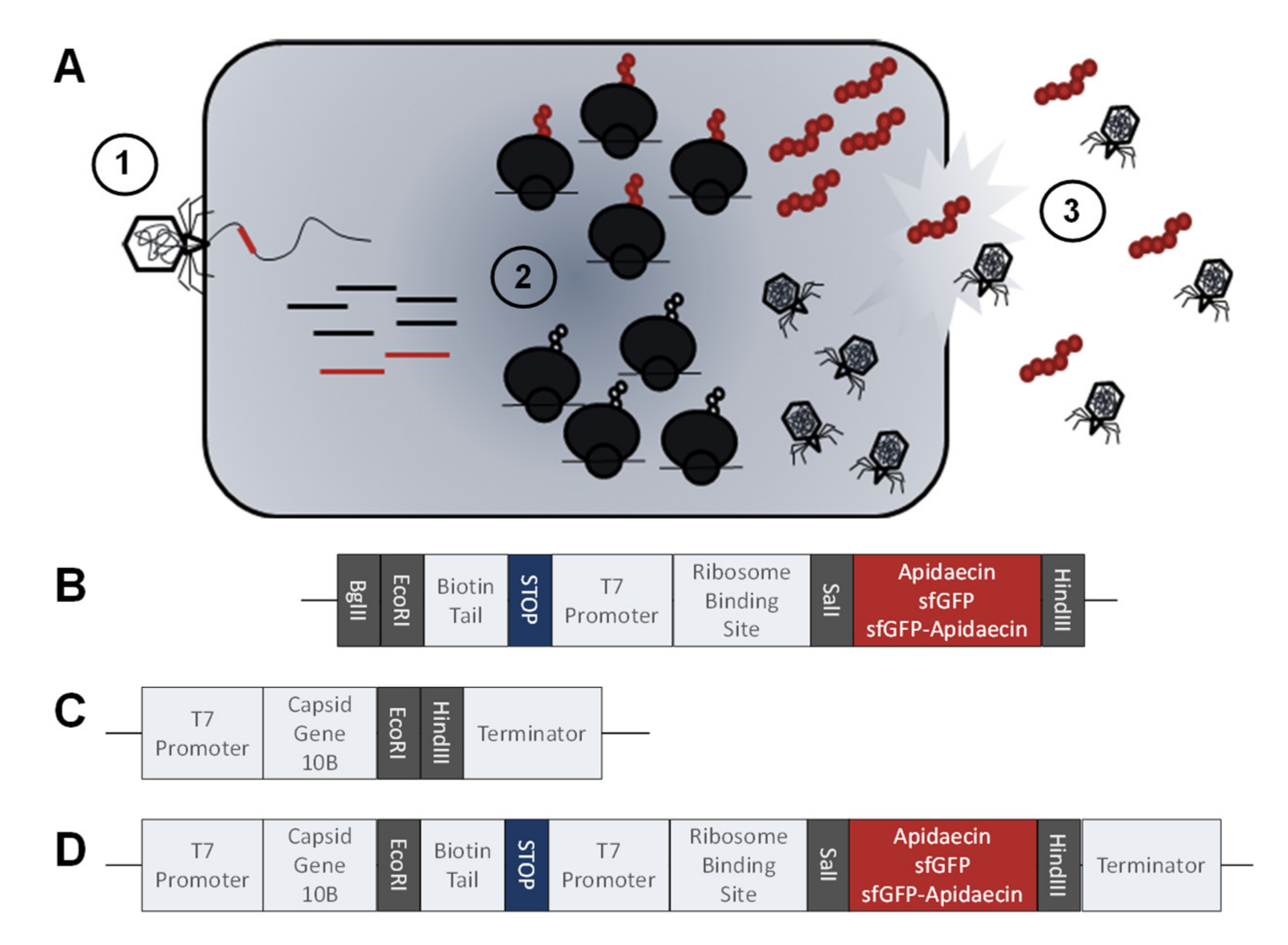
2.5. DNA constructs for Expression and Genetic Modification of Phages
2.6. Protein and Peptide Expression
2.7. Genetic Modification of Phages
2.8. Plaque Assay
2.9. Time Kill Assay
2.10. Mixed Culture Liquid Assay
3. Results
3.1. Activity of Apidaecin Analogs against E. coli
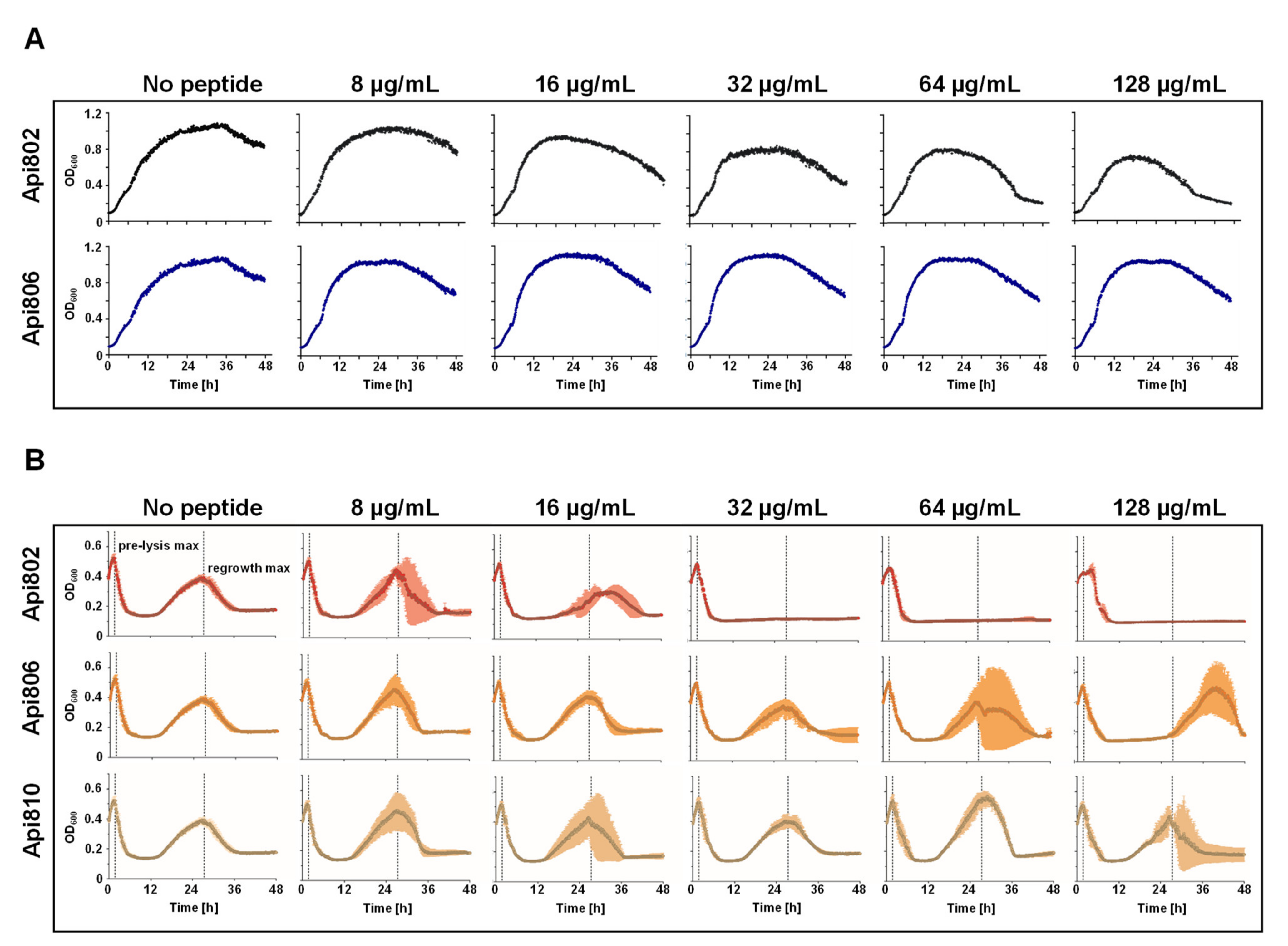
3.2. Api802 Affects Phage Mediated Lysis
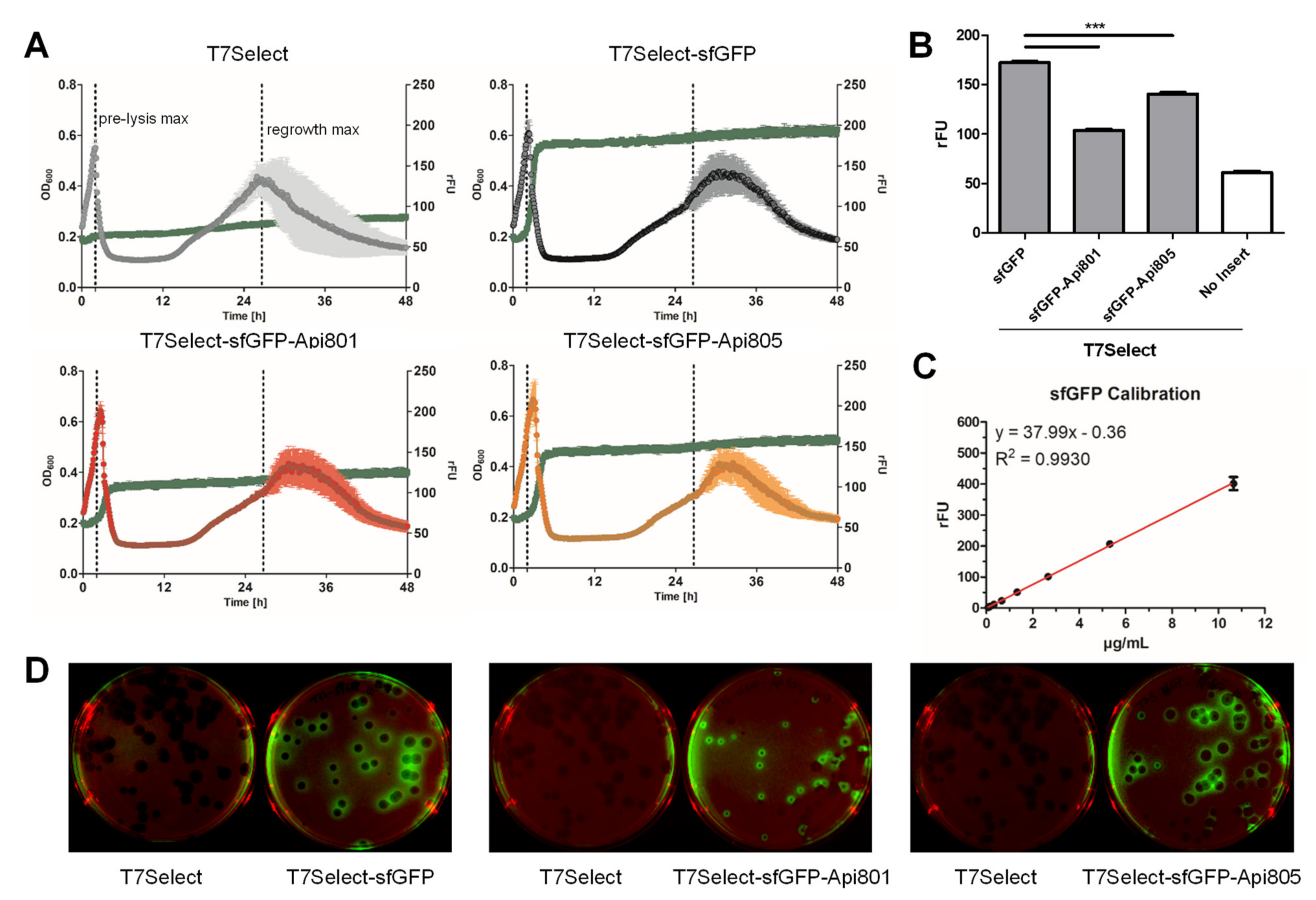
3.3. Generation of sfGFP Reporter Phages
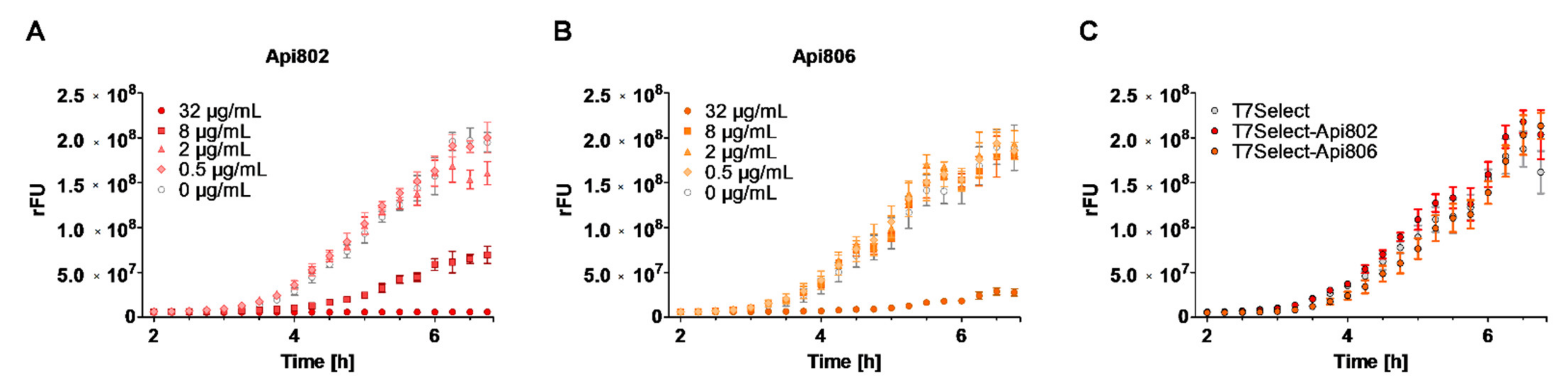
3.4. Api802 Delays Bacterial Protein Expression of Phage Proteins
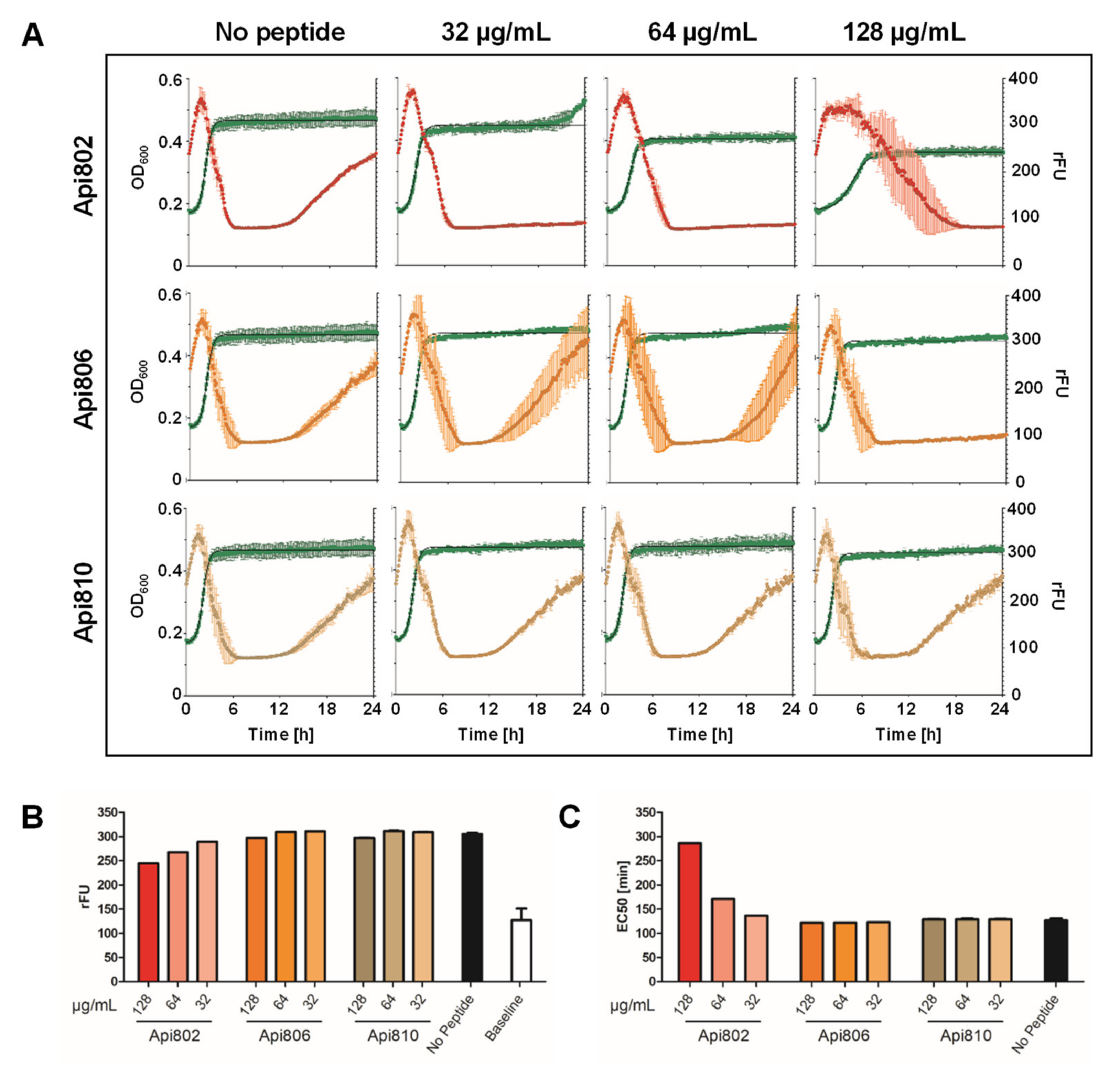
3.5. Generation and Activity of PrAMP-Phages
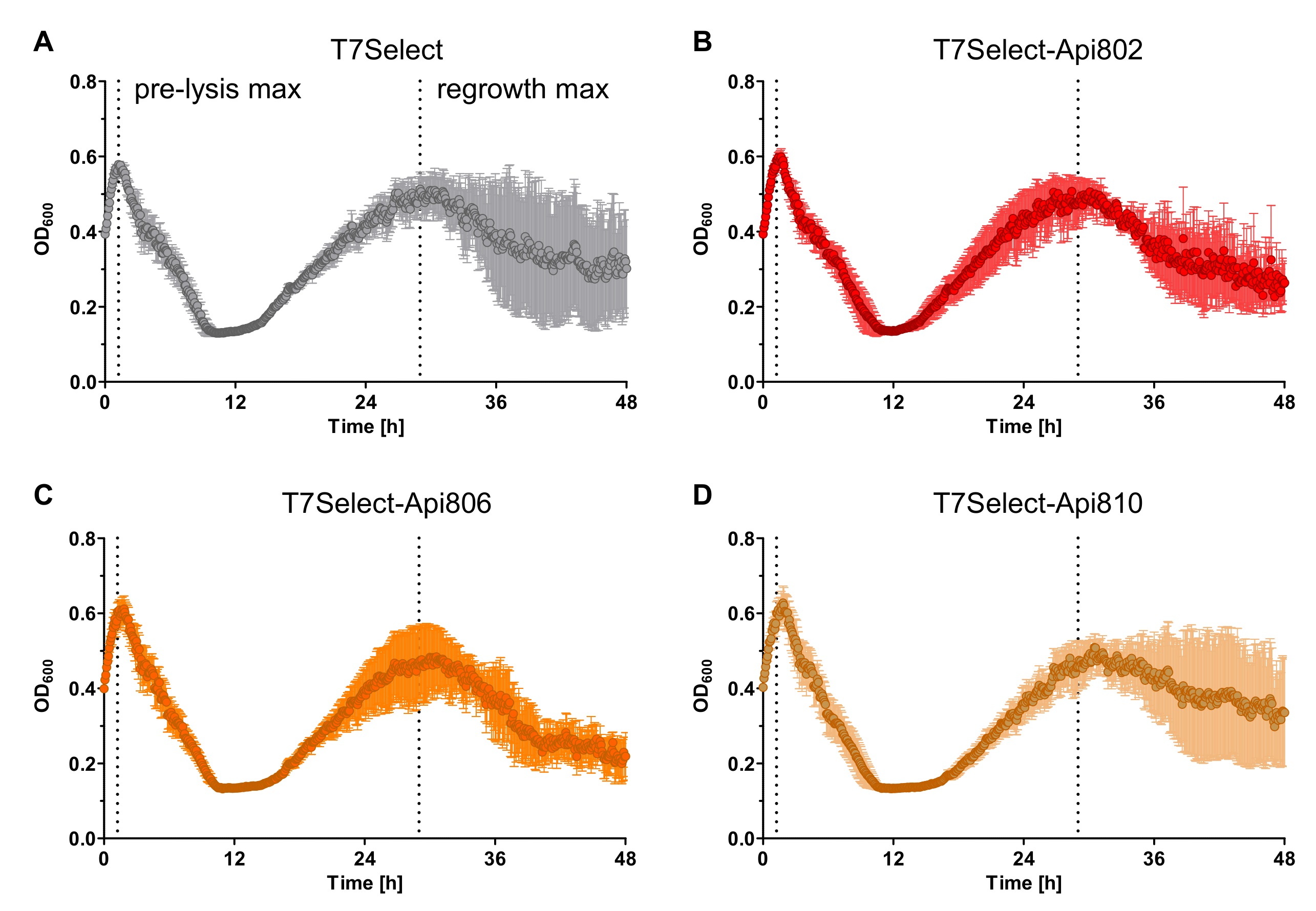
3.6. Mixed Culture Liquid Assay with PrAMP-Phages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Herelle, F. The Bacteriophage and Its Behavior; The Williams & Wilkins Company: Philadelphia, PA, USA, 1926. [Google Scholar]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G. Bacteriophage Therapy; Azeredo, J., Sillankorva, S., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; Volume 1693, ISBN 978-1-4939-7394-1. [Google Scholar]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; Le Floch, R.; et al. Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (PhagoBurn): A randomised, controlled, double-blind phase 1/2 trial. Lancet Infect. Dis. 2019, 19, 35–45. [Google Scholar] [CrossRef]
- Ascending Dose Study of the Safety of AB-SA01 When Topically Applied to Intact Skin of Healthy Adults. Available online: https://clinicaltrials.gov/ct2/show/NCT02757755?term=armata&rank=6 (accessed on 19 September 2019).
- Individual Patient Expanded Access for AB-PA01, an Investigational Anti-Pseudomonas Aeruginosa Bacteriophage Therapeutic. Available online: https://clinicaltrials.gov/ct2/show/NCT03395743?term=armata&rank=2 (accessed on 19 September 2019).
- Individual Patient Expanded Access for AB-SA01, an Investigational Anti-Staphylococcus Aureus Bacteriophage Therapeutic. Available online: https://clinicaltrials.gov/ct2/show/NCT03395769?term=armata&rank=1 (accessed on 19 September 2019).
- Huff, W.E.; Huff, G.R.; Rath, N.C.; Balog, J.M.; Donoghue, A.M. Therapeutic efficacy of bacteriophage and Baytril (enrofloxacin) individually and in combination to treat colibacillosis in broilers. Poult. Sci. 2004, 83, 1944–1947. [Google Scholar] [CrossRef] [PubMed]
- Oechslin, F.; Piccardi, P.; Mancini, S.; Gabard, J.; Moreillon, P.; Entenza, J.M.; Resch, G.; Que, Y.A. Synergistic interaction between phage therapy and antibiotics clears Pseudomonas Aeruginosa infection in endocarditis and reduces virulence. J. Infect. Dis. 2017, 215, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliaferri, T.L.; Jansen, M.; Horz, H.P. Fighting Pathogenic Bacteria on Two Fronts: Phages and Antibiotics as Combined Strategy. Front. Cell. Infect. Microbiol. 2019, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Martínez, B.; Obeso, J.M.; Rodríguez, A.; García, P. Nisin-bacteriophage crossresistance in Staphylococcus aureus. Int. J. Food Microbiol. 2008, 122, 253–258. [Google Scholar] [CrossRef]
- White, R.L.; Burgess, D.S.; Manduru, M.; Bosso, J.A. Comparison of three different in vitro methods of detecting synergy: Time-kill, checkerboard, and E test. Antimicrob. Agents Chemother. 1996, 40, 1914–1918. [Google Scholar] [CrossRef] [Green Version]
- Górski, A.; Miedzybrodzki, R.; Weber-Dabrowska, B.; Fortuna, W.; Letkiewicz, S.; Rogóz, P.; Jończyk-Matysiak, E.; Dabrowska, K.; Majewska, J.; Borysowski, J. Phage therapy: Combating infections with potential for evolving from merely a treatment for complications to targeting diseases. Front. Microbiol. 2016, 7, 1515. [Google Scholar] [CrossRef] [Green Version]
- Yoichi, M.; Abe, M.; Miyanaga, K.; Unno, H.; Tanji, Y. Alteration of tail fiber protein gp38 enables T2 phage to infect Escherichia coli O157:H7. J. Biotechnol. 2005, 115, 101–107. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lo, Y.H.; Tseng, P.W.; Chang, S.F.; Lin, Y.T.; Chen, T.S. A T3 and T7 recombinant phage acquires efficient adsorption and a broader host range. PLoS ONE 2012, 7, e30954. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Lemire, S.; Pires, D.P.; Lu, T.K. Engineering Modular Viral Scaffolds for Targeted Bacterial Population Editing. Cell Syst. 2015, 1, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Lam, C.N.; Mehta-kolte, M.G.; Martins-sorenson, N.; Eckert, B.; Lin, P.H.; Chu, K.; Moghaddasi, A.; Goldman, D.; Nguyen, H.; Chan, R.; et al. A Tail Fiber Engineering Platform for Improved Bacterial Transduction-Based Diagnostic Reagents. ACS Synth Biol. 2021, 10, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.K.; Collins, J.J. Dispersing biofilms with engineered enzymatic bacteriophage. Proc. Natl. Acad. Sci. USA 2007, 104, 11197–11202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemon, D.J.; Kay, M.K.; Titus, J.K.; Ford, A.A.; Chen, W.; Hamlin, N.J.; Hwang, Y.Y. Construction of a genetically modified T7Select phage system to express the antimicrobial peptide 1018. J. Microbiol. 2019, 57, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Kawabata, S.; Uedoi, T.; Honjoh, K.; Miyamoto, T. Construction of Leaderless-Bacteriocin-Producing Bacteriophage Targeting E. coli and Neighboring Gram-Positive Pathogens. Microbiol. Spectr. 2021, 9, e00141-21. [Google Scholar] [CrossRef] [PubMed]
- Short, F.L.; Murdoch, S.L.; Ryan, R.P. Polybacterial human disease: The ills of social networking. Trends Microbiol. 2014, 22, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casteels, P.; Ampe, C.; Jacobs, F.; Vaeck, M.; Tempst, P. Apidaecins: Antibacterial peptides from honeybees. EMBO J. 1989, 8, 2387–2391. [Google Scholar] [CrossRef]
- Ludwig, T.; Krizsan, A.; Mohammed, G.K.; Hoffmann, R. Antimicrobial Activity and 70S Ribosome Binding of Apidaecin-Derived Api805 with Increased Bacterial Uptake Rate. Antibiotics 2022, 11, 430. [Google Scholar] [CrossRef]
- Chen, J.; Alcaine, S.D.; Jackson, A.A.; Rotello, V.M.; Nugen, S.R. Development of Engineered Bacteriophages for Escherichia coli Detection and High-Throughput Antibiotic Resistance Determination. ACS Sens. 2017, 2, 484–489. [Google Scholar] [CrossRef]
- Berthold, N.; Czihal, P.; Fritsche, S.; Sauer, U.; Schiffer, G.; Knappe, D.; Alber, G.; Hoffmann, R. Novel apidaecin 1b analogs with superior serum stabilities for treatment of infections by Gram-negative pathogens. Antimicrob. Agents Chemother. 2013, 57, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Krizsan, A.; Volke, D.; Weinert, S.; Sträter, N.; Knappe, D.; Hoffmann, R. Insect-Derived Proline-Rich Antimicrobial Peptides Kill Bacteria by Inhibiting Bacterial Protein Translation at the 70 S Ribosome. Angew. Chem. Int. Ed. 2014, 53, 12236–12239. [Google Scholar] [CrossRef]
- Florin, T.; Maracci, C.; Graf, M.; Karki, P.; Klepacki, D.; Berninghausen, O.; Beckmann, R.; Vázquez-Laslop, N.; Wilson, D.N.; Rodnina, M.V.; et al. An antimicrobial peptide that inhibits translation by trapping release factors on the ribosome. Nat. Struct. Mol. Biol. 2017, 24, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.H.; Furuta, M.; Nasu, T.; Hirono, M.; Pruet, J.; Duc, H.M.; Zhang, Y.; Masuda, Y.; Honjoh, K.; Miyamoto, T. Inhibition of phage-resistant bacterial pathogen re-growth with the combined use of bacteriophages and EDTA. Food Microbiol. 2021, 100, 103853. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Feng, Y.; McNally, A.; Zong, Z. Characterization of phage resistance and phages capable of intestinal decolonization of carbapenem-resistant Klebsiella pneumoniae in mice. Commun. Biol. 2022, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Meile, S.; Baggenstos, J.; Jäggi, T.; Piffaretti, P.; Hunold, L.; Matter, C.I.; Leitner, L.; Kessler, T.M.; Loessner, M.J.; et al. Enhancing bacteriophage therapeutics through in situ production and release of heterologous antimicrobial effectors. bioRxiv 2022. [Google Scholar] [CrossRef]
- Xu, H.; Bao, X.; Hong, W.; Wang, A.; Wang, K.; Dong, H.; Hou, J.; Govinden, R.; Deng, B.; Chenia, H.Y. Biological Characterization and Evolution of Bacteriophage T7-△holin During the Serial Passage Process. Front. Microbiol. 2021, 12, 705310. [Google Scholar] [CrossRef]
- Novagen T7Select System Manual; TB178 Rev; Merck KGaA: Darmstadt, Germany, 2011.
- Volkmer, B.; Heinemann, M. Condition-Dependent cell volume and concentration of Escherichia coli to facilitate data conversion for systems biology modeling. PLoS ONE 2011, 6, e23126. [Google Scholar] [CrossRef] [Green Version]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G. Bacteriophage Therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Graf, M.; Wilson, D.N. Intracellular Antimicrobial Peptides Targeting the Protein Synthesis Machinery. In Antimicrobial Peptides: Basics for Clinical Application; Matsuzaki, K., Ed.; Springer: Singapore, 2019; pp. 73–89. ISBN 978-981-13-3588-4. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PrAMP | Sequence | MIC [µg/mL] | |
|---|---|---|---|
| 25% MHBII | LB Medium | ||
| Api801 | GNNRPVYIPRPRPPHPRL-OH | 2 | n.d. |
| Api802 | MNNRPIYIPRPRPPHPRL-OH | 2–4 | 64 |
| Api805 | GNNRPIYIPRPRPPHPRPIRV-OH | 4 | n.d. |
| Api806 | MNNRPIYIPRPRPPHPRPIRV-OH | 4–8 | 128 |
| Api810 | MNNRPIYIPRPRPPHPAL-OH | 128 | >128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ludwig, T.; Hoffmann, R.; Krizsan, A. Construction and Characterization of T7 Bacteriophages Harboring Apidaecin-Derived Sequences. Curr. Issues Mol. Biol. 2022, 44, 2554-2568. https://doi.org/10.3390/cimb44060174
Ludwig T, Hoffmann R, Krizsan A. Construction and Characterization of T7 Bacteriophages Harboring Apidaecin-Derived Sequences. Current Issues in Molecular Biology. 2022; 44(6):2554-2568. https://doi.org/10.3390/cimb44060174
Chicago/Turabian StyleLudwig, Tobias, Ralf Hoffmann, and Andor Krizsan. 2022. "Construction and Characterization of T7 Bacteriophages Harboring Apidaecin-Derived Sequences" Current Issues in Molecular Biology 44, no. 6: 2554-2568. https://doi.org/10.3390/cimb44060174