
The Androgen Hormone-Induced Increase in Androgen Receptor Protein Expression Is Caused by the Autoinduction of the Androgen Receptor Translational Activity

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. WST-1 Cell Viability Assay
2.3. Total RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR (qPCR)
2.4. Western Blot Analysis
2.5. Immunofluorescence
2.6. eosFP-AR Transfection
2.7. Statistical Analysis
3. Results
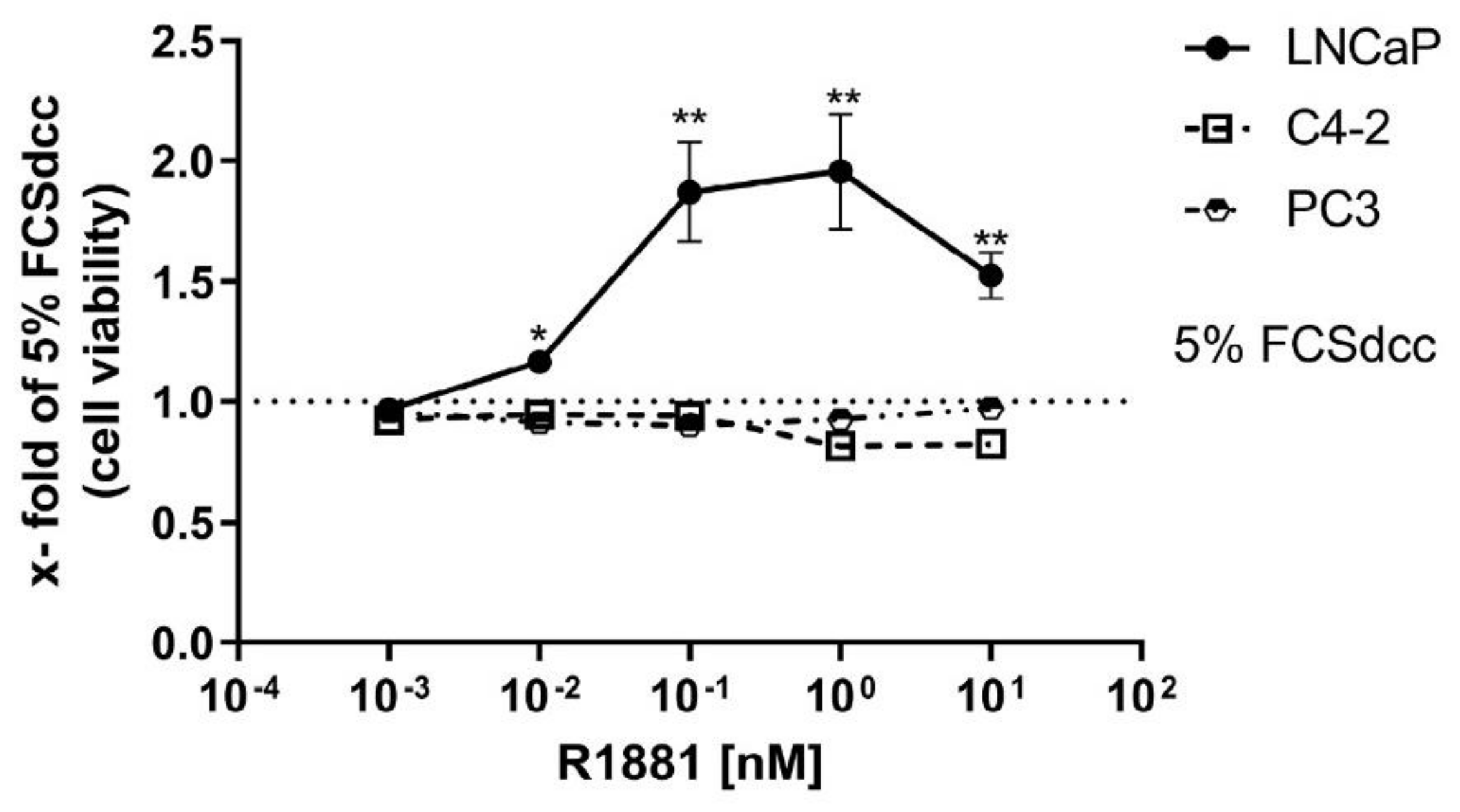
3.1. High R1881 Concentrations Inhibit the Cell Viability in LNCaP Cells
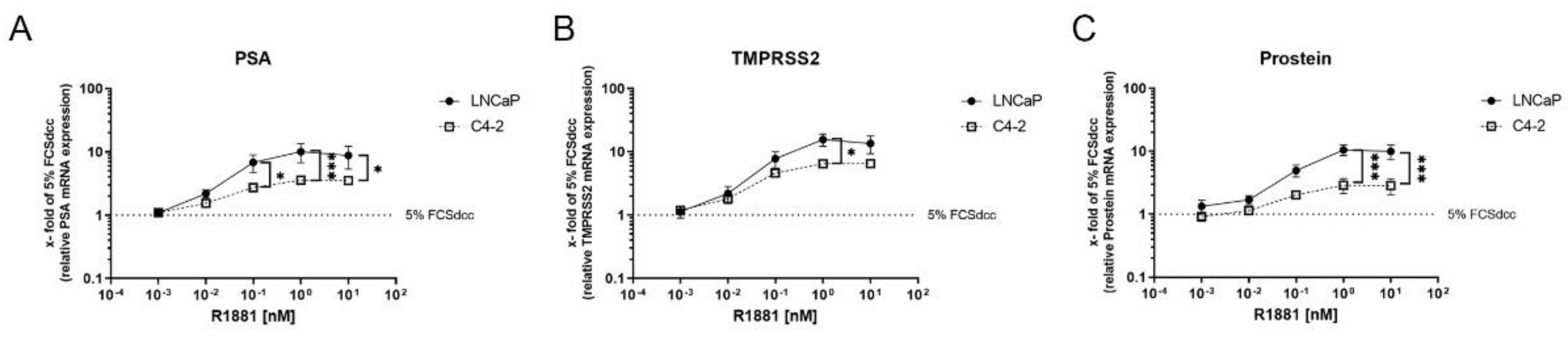
3.2. High R1881 Concentrations Do Not Inhibit AR-Mediated Gene Transcription
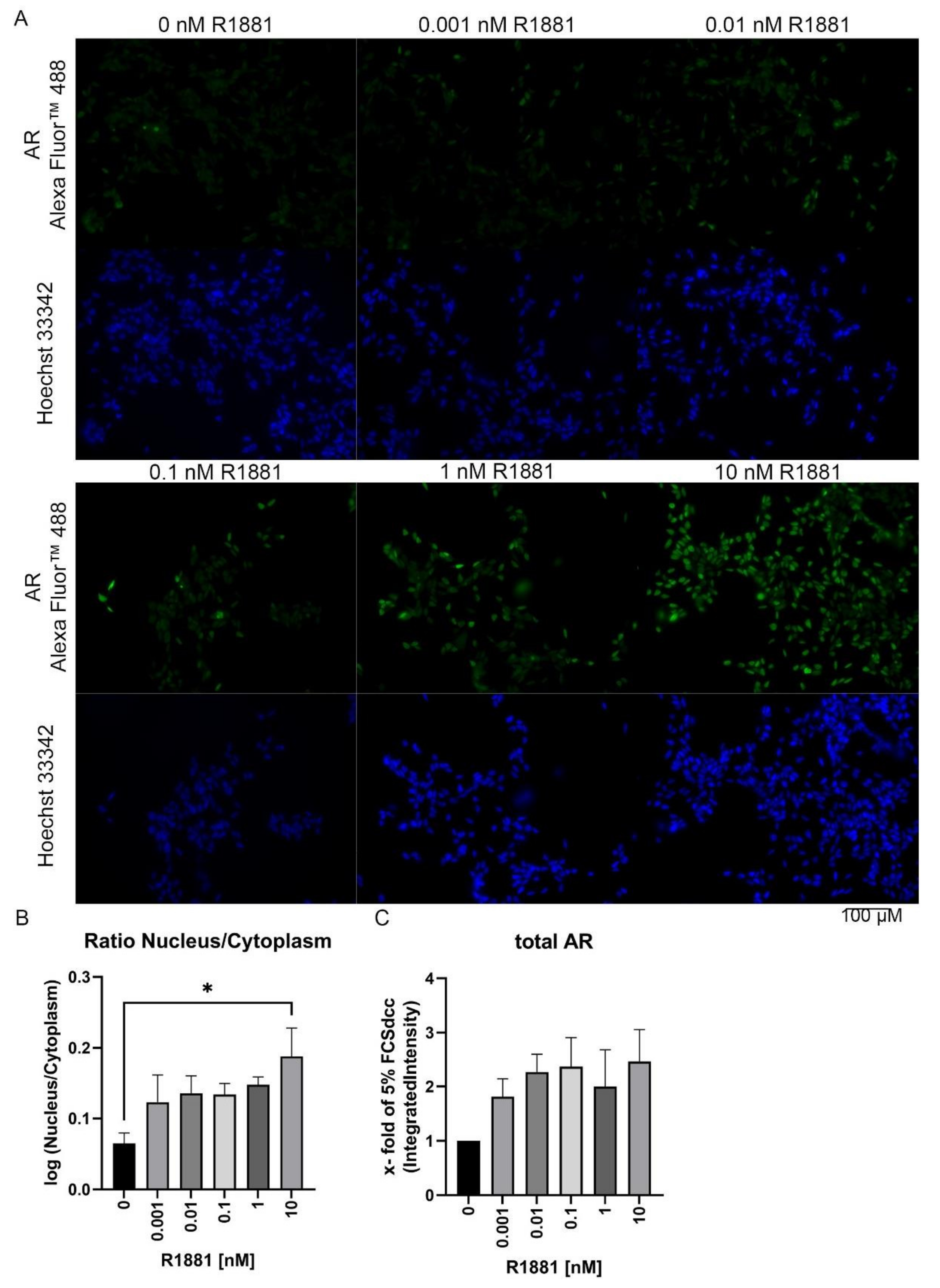
3.3. R1881 Activates Nuclear AR Translocation in a Time and Concentration-Dependent Manner
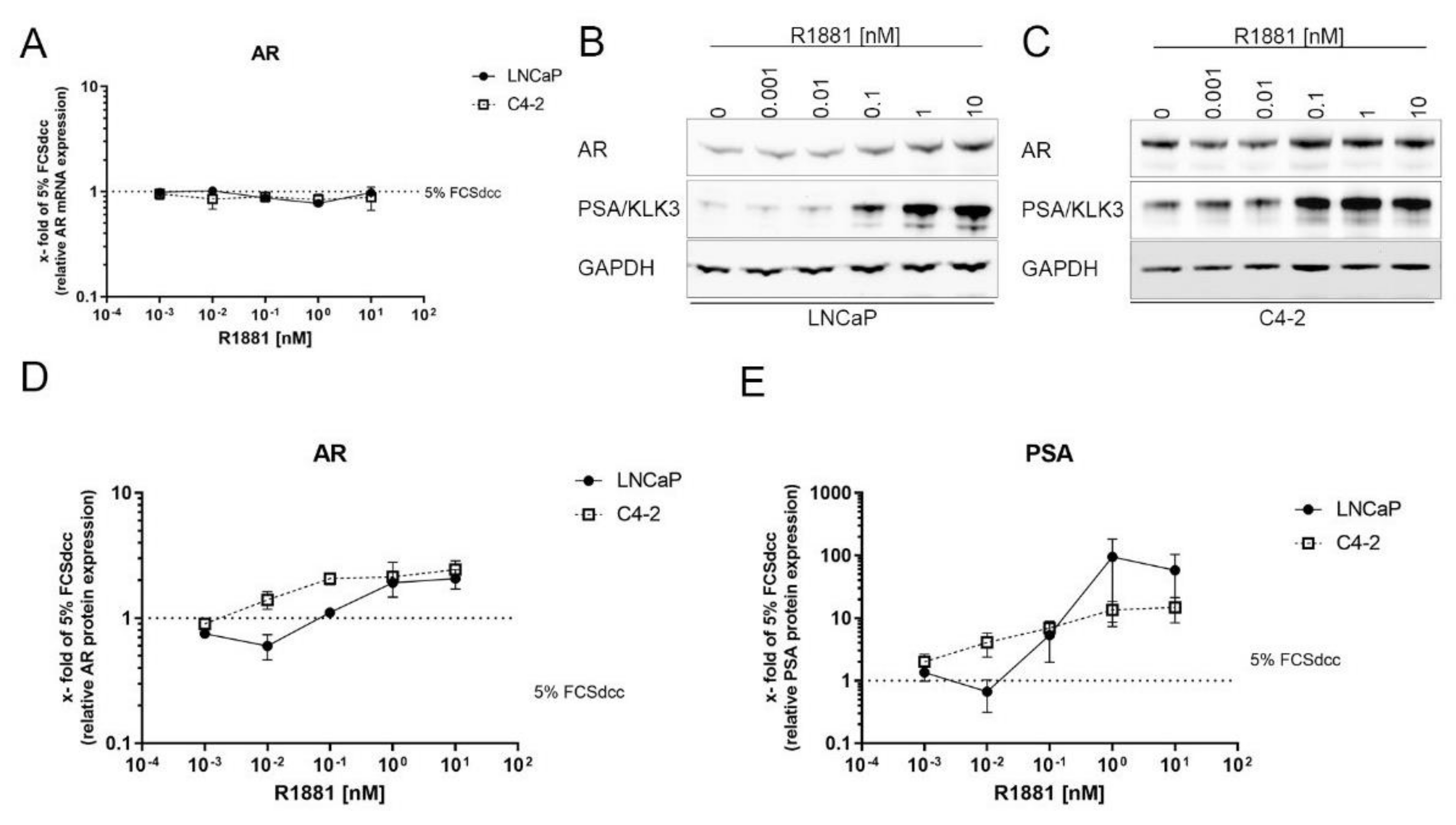
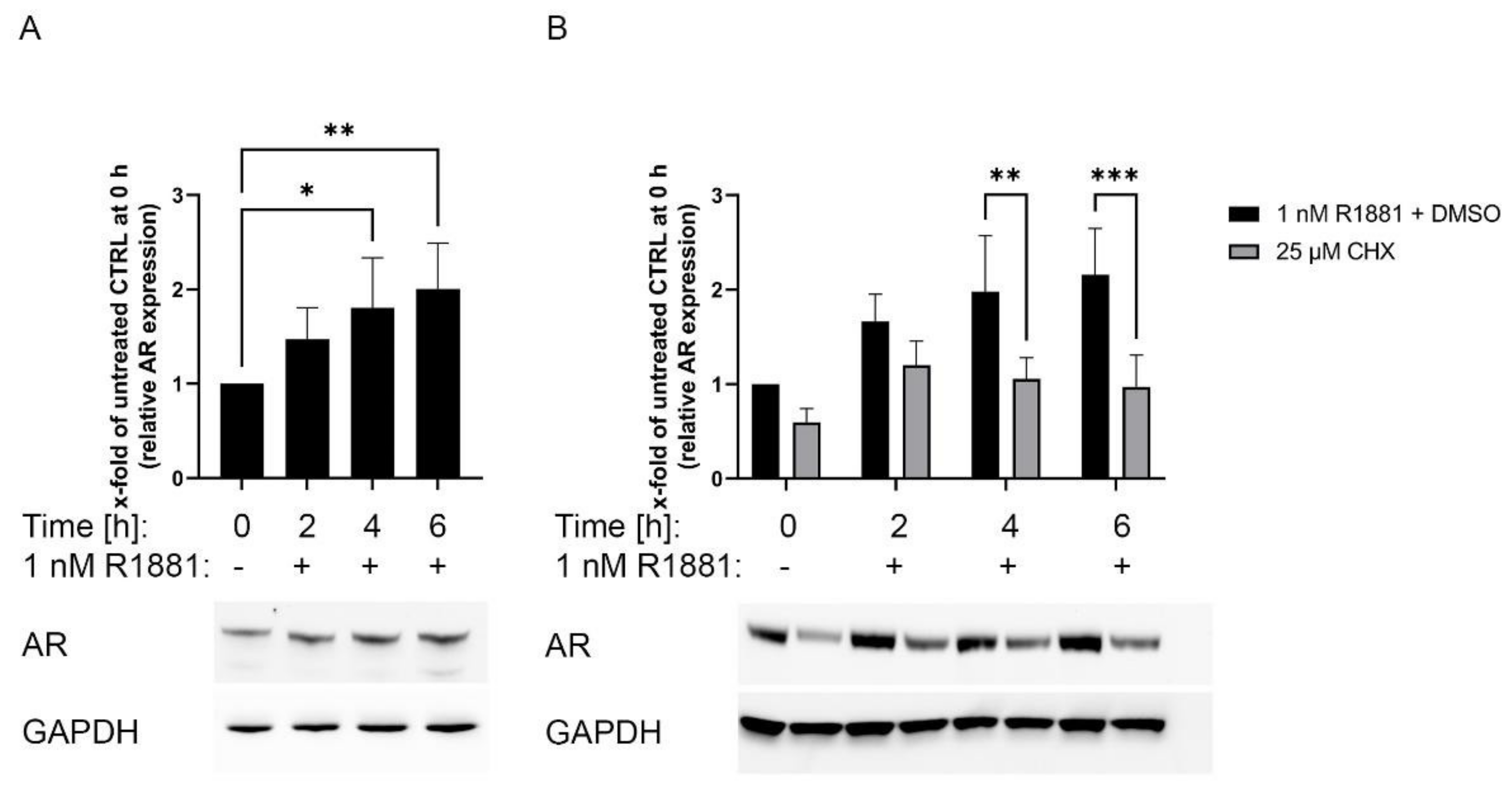
3.4. AR Is Stabilised Transcriptionally Independent by R1881
3.5. Inhibition of the Protein Synthesis by Cycloheximide Prevents R1881 Induced AR Protein Increase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, C.; Kokontis, J.; Liao, S. Molecular cloning of human and rat complementary DNA encoding androgen receptors. Science 1988, 240, 324–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schutz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.K.; Lavrovsky, Y.; Song, C.S.; Chen, S.; Jung, M.H.; Velu, N.K.; Bi, B.Y.; Chatterjee, B. Regulation of androgen action. Vitam. Horm. 1999, 55, 309–352. [Google Scholar]
- Green, S.M.; Mostaghel, E.A.; Nelson, P.S. Androgen action and metabolism in prostate cancer. Mol. Cell. Endocrinol. 2012, 360, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, R.A.; Grossmann, M. Androgen Receptor Structure, Function and Biology: From Bench to Bedside. Clin. Biochem. Rev. 2016, 37, 3–15. [Google Scholar]
- Gelmann, E.P. Molecular biology of the androgen receptor. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 3001–3015. [Google Scholar] [CrossRef] [PubMed]
- Claessens, F.; Denayer, S.; Van Tilborgh, N.; Kerkhofs, S.; Helsen, C.; Haelens, A. Diverse roles of androgen receptor (AR) domains in AR-mediated signaling. Nucl. Recept. Signal. 2008, 6, e008. [Google Scholar] [CrossRef] [Green Version]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev. 2004, 25, 276–308. [Google Scholar] [CrossRef] [Green Version]
- Eder, I.E.; Culig, Z.; Putz, T.; Nessler-Menardi, C.; Bartsch, G.; Klocker, H. Molecular biology of the androgen receptor: From molecular understanding to the clinic. Eur. Urol. 2001, 40, 241–251. [Google Scholar] [CrossRef]
- Crona, D.J.; Whang, Y.E. Androgen Receptor-Dependent and -Independent Mechanisms Involved in Prostate Cancer Therapy Resistance. Cancers 2017, 9, 67. [Google Scholar] [CrossRef]
- Bevan, C.; Parker, M. The role of coactivators in steroid hormone action. Exp. Cell Res. 1999, 253, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.P. The role of coactivators and corepressors in the biology and mechanism of action of steroid hormone receptors. J. Mammary Gland Biol. Neoplasia 2000, 5, 307–324. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Kim, J.; Yu, J. Androgen receptor genomic regulation. Transl. Androl. Urol. 2013, 2, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.R. Human androgen insensitivity syndrome. J. Androl. 1995, 16, 299–303. [Google Scholar]
- McEwan, I.J. Structural and functional alterations in the androgen receptor in spinal bulbar muscular atrophy. Biochem. Soc. Trans. 2001, 29, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kumar, N.; Thakur, D.S.; Patidar, A. Male hypogonadism: Symptoms and treatment. J. Adv. Pharm. Technol. Res. 2010, 1, 297–301. [Google Scholar] [CrossRef]
- Webber, R. Benign prostatic hyperplasia. Clin. Evid. 2006, 2006, 1213–1226. [Google Scholar]
- Watson, P.A.; Arora, V.K.; Sawyers, C.L. Emerging mechanisms of resistance to androgen receptor inhibitors in prostate cancer. Nat. Rev. Cancer 2015, 15, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Cornford, P.; van den Bergh, R.C.N.; Briers, E.; Van den Broeck, T.; Cumberbatch, M.G.; De Santis, M.; Fanti, S.; Fossati, N.; Gandaglia, G.; Gillessen, S.; et al. EAU-EANM-ESTRO-ESUR-SIOG Guidelines on Prostate Cancer. Part II-2020 Update: Treatment of Relapsing and Metastatic Prostate Cancer. Eur. Urol. 2021, 79, 263–282. [Google Scholar] [CrossRef]
- Nordeen, S.K.; Su, L.J.; Osborne, G.A.; Hayman, P.M.; Orlicky, D.J.; Wessells, V.M.; van Bokhoven, A.; Flaig, T.W. Titration of Androgen Signaling: How Basic Studies Have Informed Clinical Trials Using High-Dose Testosterone Therapy in Castrate-Resistant Prostate Cancer. Life 2021, 11, 884. [Google Scholar] [CrossRef] [PubMed]
- Horoszewicz, J.S.; Leong, S.S.; Chu, T.M.; Wajsman, Z.L.; Friedman, M.; Papsidero, L.; Kim, U.; Chai, L.S.; Kakati, S.; Arya, S.K.; et al. The LNCaP cell line—A new model for studies on human prostatic carcinoma. Prog. Clin. Biol. Res. 1980, 37, 115–132. [Google Scholar] [PubMed]
- Thalmann, G.N.; Sikes, R.A.; Wu, T.T.; Degeorges, A.; Chang, S.M.; Ozen, M.; Pathak, S.; Chung, L.W. LNCaP progression model of human prostate cancer: Androgen-independence and osseous metastasis. Prostate 2000, 44, 91–103. [Google Scholar] [CrossRef]
- Tai, S.; Sun, Y.; Squires, J.M.; Zhang, H.; Oh, W.K.; Liang, C.Z.; Huang, J. PC3 is a cell line characteristic of prostatic small cell carcinoma. Prostate 2011, 71, 1668–1679. [Google Scholar] [CrossRef] [Green Version]
- Kaighn, M.E.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.F.; Jones, L.W. Establishment and characterization of a human prostatic carcinoma cell line (PC-3). Investig. Urol. 1979, 17, 16–23. [Google Scholar]
- Erb, H.H.; Bodenbender, J.; Handle, F.; Diehl, T.; Donix, L.; Tsaur, I.; Gleave, M.; Haferkamp, A.; Huber, J.; Fuessel, S. Assessment of STAT5 as a potential therapy target in enzalutamide-resistant prostate cancer. PLoS ONE 2020, 15, e0237248. [Google Scholar] [CrossRef]
- McQuin, C.; Goodman, A.; Chernyshev, V.; Kamentsky, L.; Cimini, B.A.; Karhohs, K.W.; Doan, M.; Ding, L.; Rafelski, S.M.; Thirstrup, D.; et al. CellProfiler 3.0: Next-generation image processing for biology. PLoS Biol. 2018, 16, e2005970. [Google Scholar] [CrossRef] [Green Version]
- Rinnab, L.; Schütz, S.V.; Diesch, J.; Schmid, E.; Küfer, R.; Hautmann, R.E.; Spindler, K.-D.; Cronauer, M.V. Inhibition of glycogen synthase kinase-3 in androgen-responsive prostate cancer cell lines: Are GSK inhibitors therapeutically useful? Neoplasia 2008, 10, 624–634. [Google Scholar] [CrossRef] [Green Version]
- Nienhaus, G.U.; Karin, N.; Angela, H.; Sergey, I.; Fabiana, R.; Franz, O.; Michael, W.; Florian, S.; Carlheinz, R.; Beatrice, V.; et al. Photoconvertible Fluorescent Protein EosFP: Biophysical Properties and Cell Biology Applications. Photochem. Photobiol. 2006, 82, 351–358. [Google Scholar] [CrossRef]
- Ivanchenko, S.; Röcker, C.; Oswald, F.; Wiedenmann, J.; Nienhaus, G.U. Targeted Green-Red Photoconversion of EosFP, a Fluorescent Marker Protein. J. Biol. Phys. 2005, 31, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Vaux, D.L. Research methods: Know when your numbers are significant. Nature 2012, 492, 180–181. [Google Scholar] [CrossRef] [PubMed]
- Massie, C.E.; Lynch, A.; Ramos-Montoya, A.; Boren, J.; Stark, R.; Fazli, L.; Warren, A.; Scott, H.; Madhu, B.; Sharma, N.; et al. The androgen receptor fuels prostate cancer by regulating central metabolism and biosynthesis. EMBO J. 2011, 30, 2719–2733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.; Wang, Y.; Yue, H.H.; Zhang, Y.T.; Shi, C.H.; Liu, F.; Bao, T.Y.; Yang, Z.Y.; Yuan, J.L.; Shao, G.X. Biphasic effect of androgens on prostate cancer cells and its correlation with androgen receptor coactivator dopa decarboxylase. J. Androl. 2007, 28, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Androgens: Biphasic dose responses. Crit. Rev. Toxicol. 2001, 31, 517–522. [Google Scholar] [CrossRef] [PubMed]
- De Launoit, Y.; Veilleux, R.; Dufour, M.; Simard, J.; Labrie, F. Characteristics of the biphasic action of androgens and of the potent antiproliferative effects of the new pure antiestrogen EM-139 on cell cycle kinetic parameters in LNCaP human prostatic cancer cells. Cancer Res. 1991, 51, 5165–5170. [Google Scholar] [PubMed]
- Watson, P.A.; Chen, Y.F.; Balbas, M.D.; Wongvipat, J.; Socci, N.D.; Viale, A.; Kim, K.; Sawyers, C.L. Constitutively active androgen receptor splice variants expressed in castration-resistant prostate cancer require full-length androgen receptor. Proc. Natl. Acad. Sci. USA 2010, 107, 16759–16765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Y.; Dai, B.; Ye, D.; Kong, Y.; Chang, K.; Jia, Z.; Yang, X.; Zhang, H.; Zhu, Y.; Shi, G. Constitutively active AR-V7 plays an essential role in the development and progression of castration-resistant prostate cancer. Sci. Rep. 2015, 5, 7654. [Google Scholar] [CrossRef] [Green Version]
- Wadosky, K.M.; Koochekpour, S. Androgen receptor splice variants and prostate cancer: From bench to bedside. Oncotarget 2017, 8, 18550–18576. [Google Scholar] [CrossRef] [Green Version]
- Teply, B.A.; Kachhap, S.; Eisenberger, M.A.; Denmeade, S.R. Extreme Response to High-dose Testosterone in BRCA2- and ATM-mutated Prostate Cancer. Eur. Urol. 2017, 71, 499. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Niu, Y.; Lee, S.O.; Chang, C. Androgen receptor (AR) positive vs negative roles in prostate cancer cell deaths including apoptosis, anoikis, entosis, necrosis and autophagic cell death. Cancer Treat. Rev. 2014, 40, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Van der Griend, D.J.; Litvinov, I.V.; Isaacs, J.T. Conversion of androgen receptor signaling from a growth suppressor in normal prostate epithelial cells to an oncogene in prostate cancer cells involves a gain of function in c-Myc regulation. Int. J. Biol. Sci. 2014, 10, 627–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinthus, J.H.; Bryskin, I.; Trachtenberg, J.; Lu, J.-P.; Singh, G.; Fridman, E.; Wilson, B.C. Androgen induces adaptation to oxidative stress in prostate cancer: Implications for treatment with radiation therapy. Neoplasia 2007, 9, 68–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyquist, M.D.; Corella, A.; Mohamad, O.; Coleman, I.; Kaipainen, A.; Kuppers, D.A.; Lucas, J.M.; Paddison, P.J.; Plymate, S.R.; Nelson, P.S.; et al. Molecular determinants of response to high-dose androgen therapy in prostate cancer. JCI Insight 2019, 4, e129715. [Google Scholar] [CrossRef] [PubMed]
- Daniels, G.; Jha, R.; Shen, Y.; Logan, S.K.; Lee, P. Androgen receptor coactivators that inhibit prostate cancer growth. Am. J. Clin. Exp. Urol. 2014, 2, 62–70. [Google Scholar]
- Dai, J.L.; Burnstein, K.L. Two androgen response elements in the androgen receptor coding region are required for cell-specific up-regulation of receptor messenger RNA. Mol. Endocrinol. 1996, 10, 1582–1594. [Google Scholar] [CrossRef]
- Mora, G.R.; Prins, G.S.; Mahesh, V.B. Autoregulation of androgen receptor protein and messenger RNA in rat ventral prostate is protein synthesis dependent. J. Steroid Biochem. Mol. Biol. 1996, 58, 539–549. [Google Scholar] [CrossRef]
- Siciliano, T.; Simons, I.H.; Beier, A.-M.K.; Ebersbach, C.; Aksoy, C.; Seed, R.I.; Stope, M.B.; Thomas, C.; Erb, H.H.H. A Systematic Comparison of Antiandrogens Identifies Androgen Receptor Protein Stability as an Indicator for Treatment Response. Life 2021, 11, 874. [Google Scholar] [CrossRef]
- Yeap, B.B.; Krueger, R.G.; Leedman, P.J. Differential posttranscriptional regulation of androgen receptor gene expression by androgen in prostate and breast cancer cells. Endocrinology 1999, 140, 3282–3291. [Google Scholar] [CrossRef]
- Erb, H.H.H.; Oster, M.A.; Gelbrich, N.; Cammann, C.; Thomas, C.; Mustea, A.; Stope, M.B. Enzalutamide-induced Proteolytic Degradation of the Androgen Receptor in Prostate Cancer Cells Is Mediated Only to a Limited Extent by the Proteasome System. Anticancer Res. 2021, 41, 3271–3279. [Google Scholar] [CrossRef]
- Lakshmana, G.; Baniahmad, A. Interference with the androgen receptor protein stability in therapy-resistant prostate cancer. Int. J. Cancer 2019, 144, 1775–1779. [Google Scholar] [CrossRef]
- Liu, C.; Lou, W.; Yang, J.C.; Liu, L.; Armstrong, C.M.; Lombard, A.P.; Zhao, R.; Noel, O.D.V.; Tepper, C.G.; Chen, H.-W.; et al. Proteostasis by STUB1/HSP70 complex controls sensitivity to androgen receptor targeted therapy in advanced prostate cancer. Nat. Commun. 2018, 9, 4700. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Horn, J.L.; Banda, K.; Goodman, A.Z.; Lim, Y.; Jana, S.; Arora, S.; Germanos, A.A.; Wen, L.; Hardin, W.R.; et al. The androgen receptor regulates a druggable translational regulon in advanced prostate cancer. Sci. Transl. Med. 2019, 11, 503. [Google Scholar] [CrossRef] [PubMed]
- Stone, L. AR—The link between transcription and translation. Nat. Rev. Urol. 2019, 16, 565. [Google Scholar] [CrossRef] [PubMed]
- Overcash, R.F.; Chappell, V.A.; Green, T.; Geyer, C.B.; Asch, A.S.; Ruiz-Echevarria, M.J. Androgen signaling promotes translation of TMEFF2 in prostate cancer cells via phosphorylation of the alpha subunit of the translation initiation factor 2. PLoS ONE 2013, 8, e55257. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Characteristics | AR Status | Origin | References |
|---|---|---|---|---|
| LNCaP | Androgen dependent | AR T877A | Lymph node | [22] |
| C4-2 | Androgen independent | AR T877A | Lymph node | [23] |
| PC3 | small cell neuroendocrine carcinoma | AR negativ | Bone metastasis | [24,25] |
| Name | Company | Lot | Dilution |
|---|---|---|---|
| Androgen Receptor (D6F11) XP Rabbit mAb | Cell Signaling Technology | 9 | 1:5000 |
| Mouse Monoclonal anti-GAPDH (6C5) | Novus Biologicals | 19/05-G4cc-C5cc | 1:10,000 |
| Polyclonal Rabbit Anti-Mouse Immunoglobulins/HRP | Agilent Technologies | 20066043 | 1:10,000 |
| Polyclonal Swine Anti-Rabbit Immunoglobulins/HRP | Agilent Technologies | 41289300 | 1:10,000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siciliano, T.; Sommer, U.; Beier, A.-M.K.; Stope, M.B.; Borkowetz, A.; Thomas, C.; Erb, H.H.H. The Androgen Hormone-Induced Increase in Androgen Receptor Protein Expression Is Caused by the Autoinduction of the Androgen Receptor Translational Activity. Curr. Issues Mol. Biol. 2022, 44, 597-608. https://doi.org/10.3390/cimb44020041
Siciliano T, Sommer U, Beier A-MK, Stope MB, Borkowetz A, Thomas C, Erb HHH. The Androgen Hormone-Induced Increase in Androgen Receptor Protein Expression Is Caused by the Autoinduction of the Androgen Receptor Translational Activity. Current Issues in Molecular Biology. 2022; 44(2):597-608. https://doi.org/10.3390/cimb44020041
Chicago/Turabian StyleSiciliano, Tiziana, Ulrich Sommer, Alicia-Marie K. Beier, Matthias B. Stope, Angelika Borkowetz, Christian Thomas, and Holger H. H. Erb. 2022. "The Androgen Hormone-Induced Increase in Androgen Receptor Protein Expression Is Caused by the Autoinduction of the Androgen Receptor Translational Activity" Current Issues in Molecular Biology 44, no. 2: 597-608. https://doi.org/10.3390/cimb44020041