
Doxycycline Alters the Porcine Renal Proteome and Degradome during Hypothermic Machine Perfusion

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
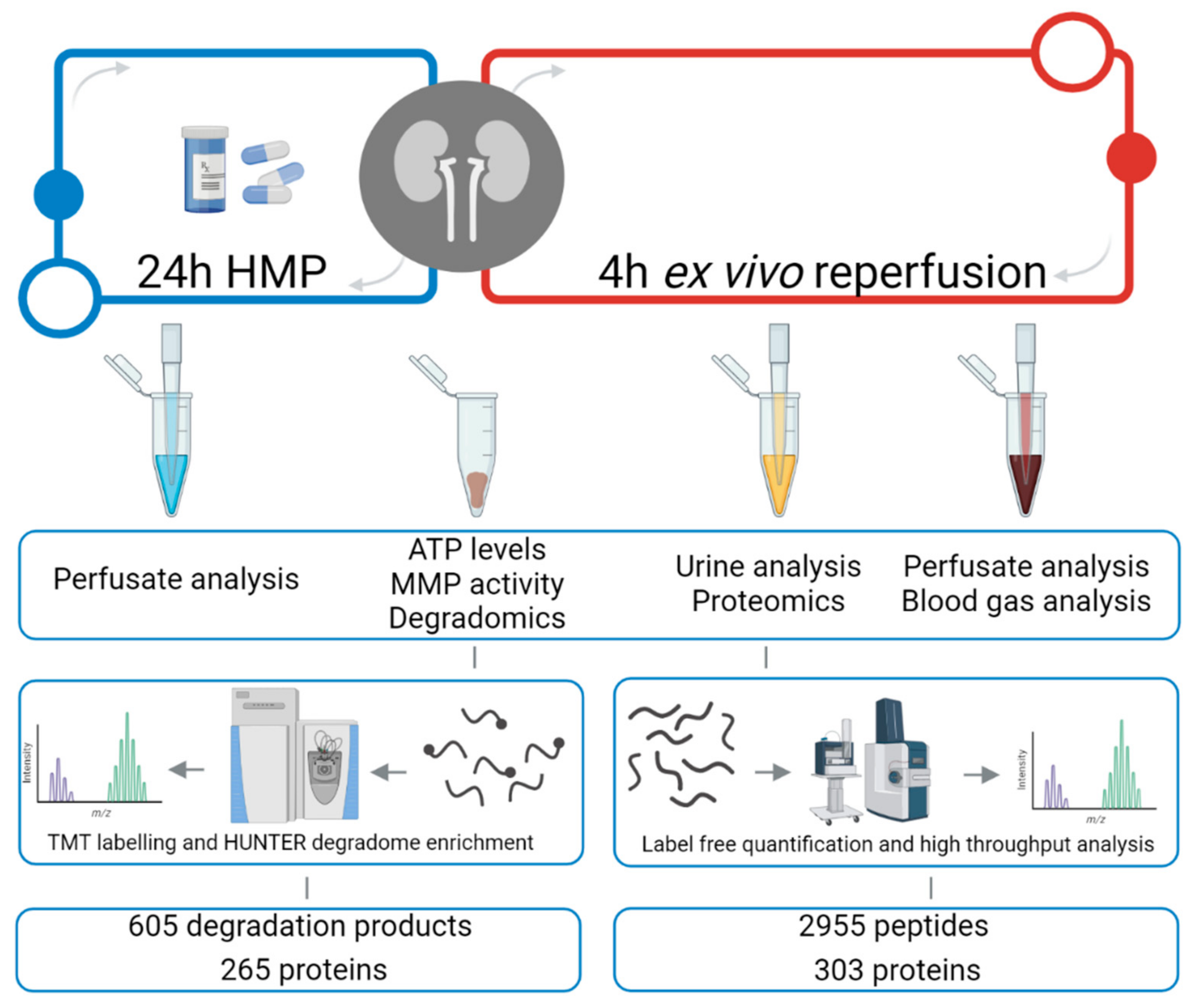
2.1. Experimental Design
2.2. Proteomic Analysis of Urine Using an In-Solution Digestion
2.3. Tissue Degradome Analysis Using a TMT-HUNTER Workflow
2.4. Visualization and Statistics of Proteomics and Degradomics Data
3. Results
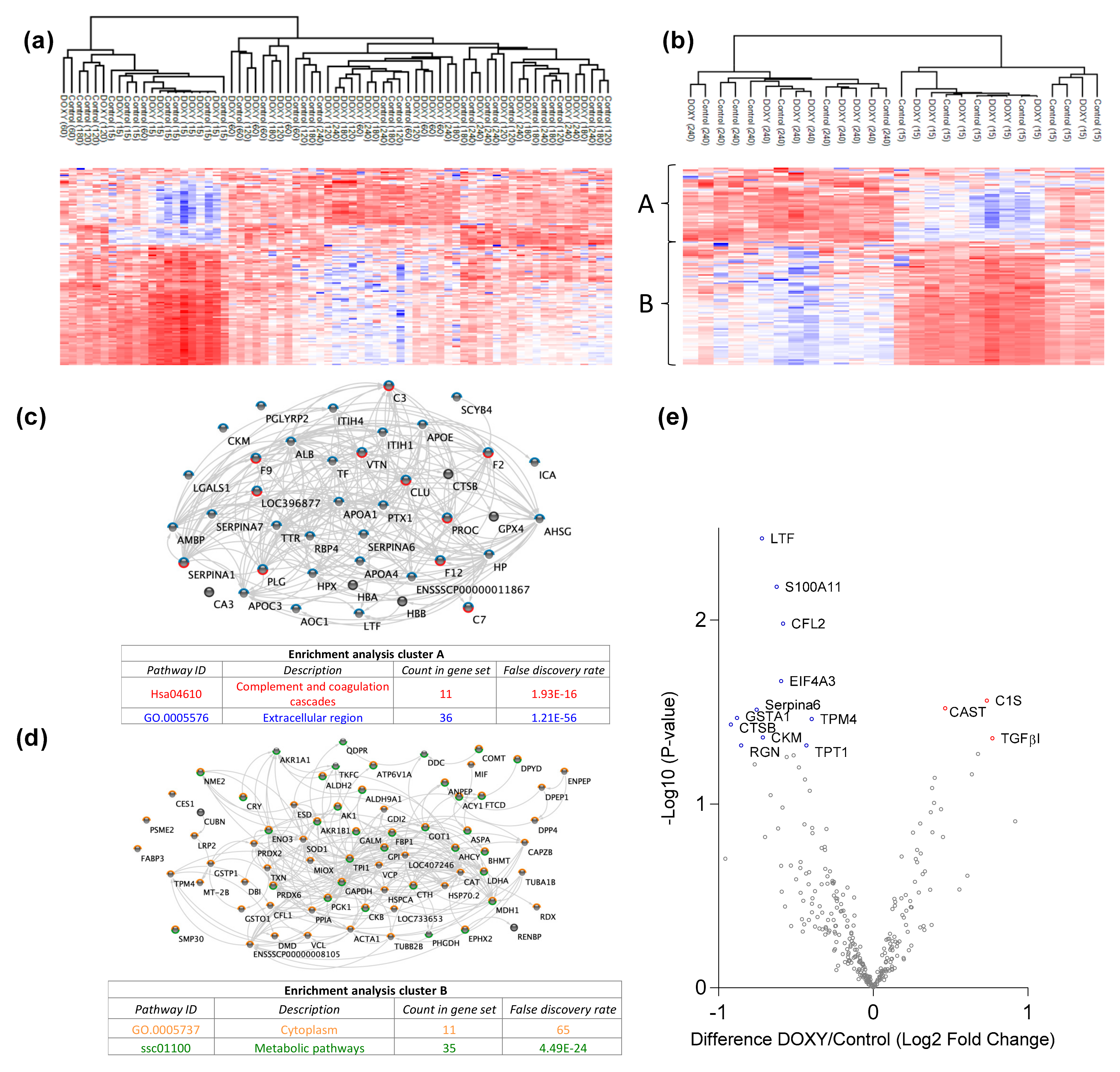
3.1. Kidney Urine Proteomics and Tissue Degradomics Profiles
3.2. Doxycycline Causes Molecular Alterations in Urine during Machine Perfusion
3.3. Doxycycline Alters Renal Degradome
3.4. Protease Activity in Experimental Groups
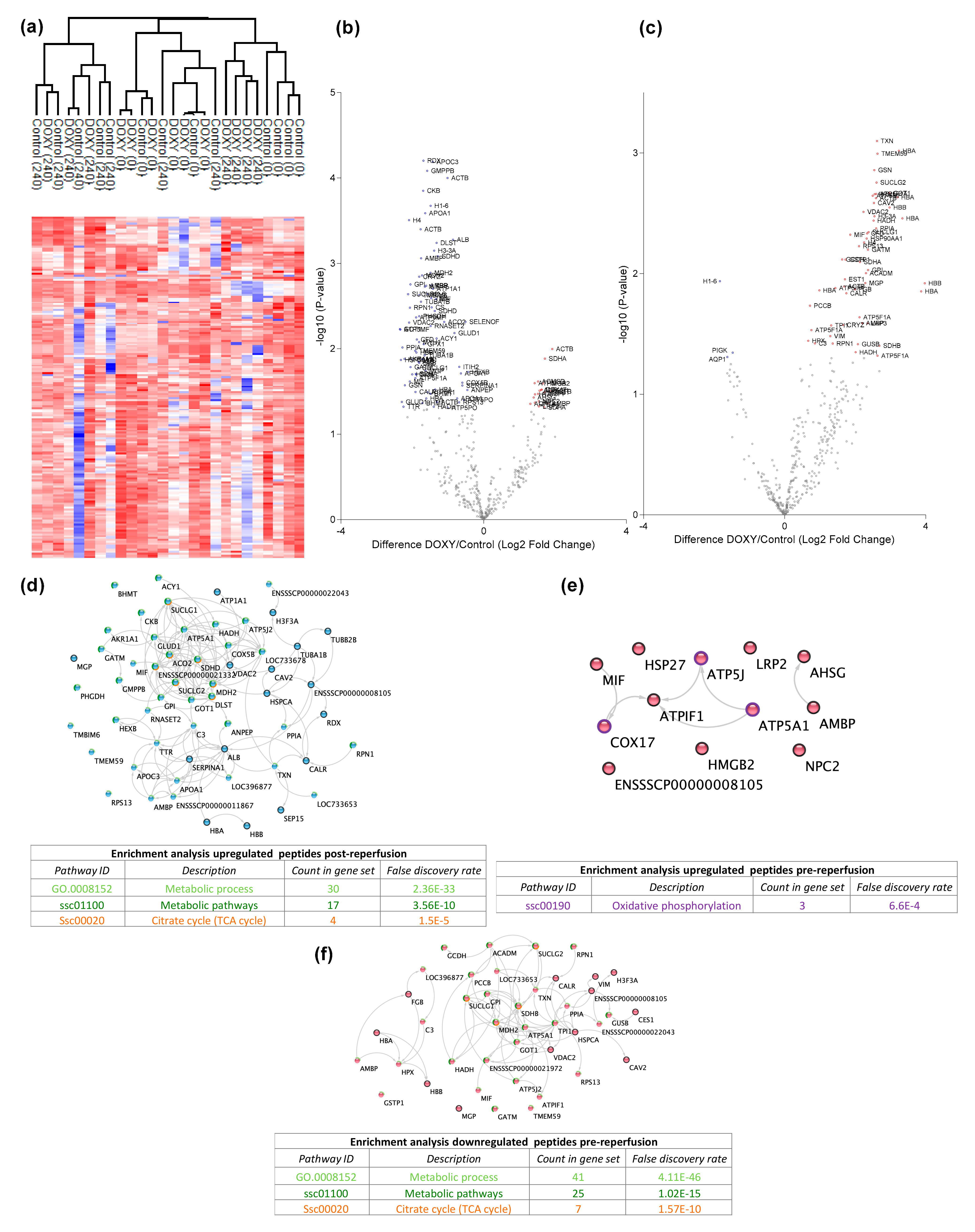
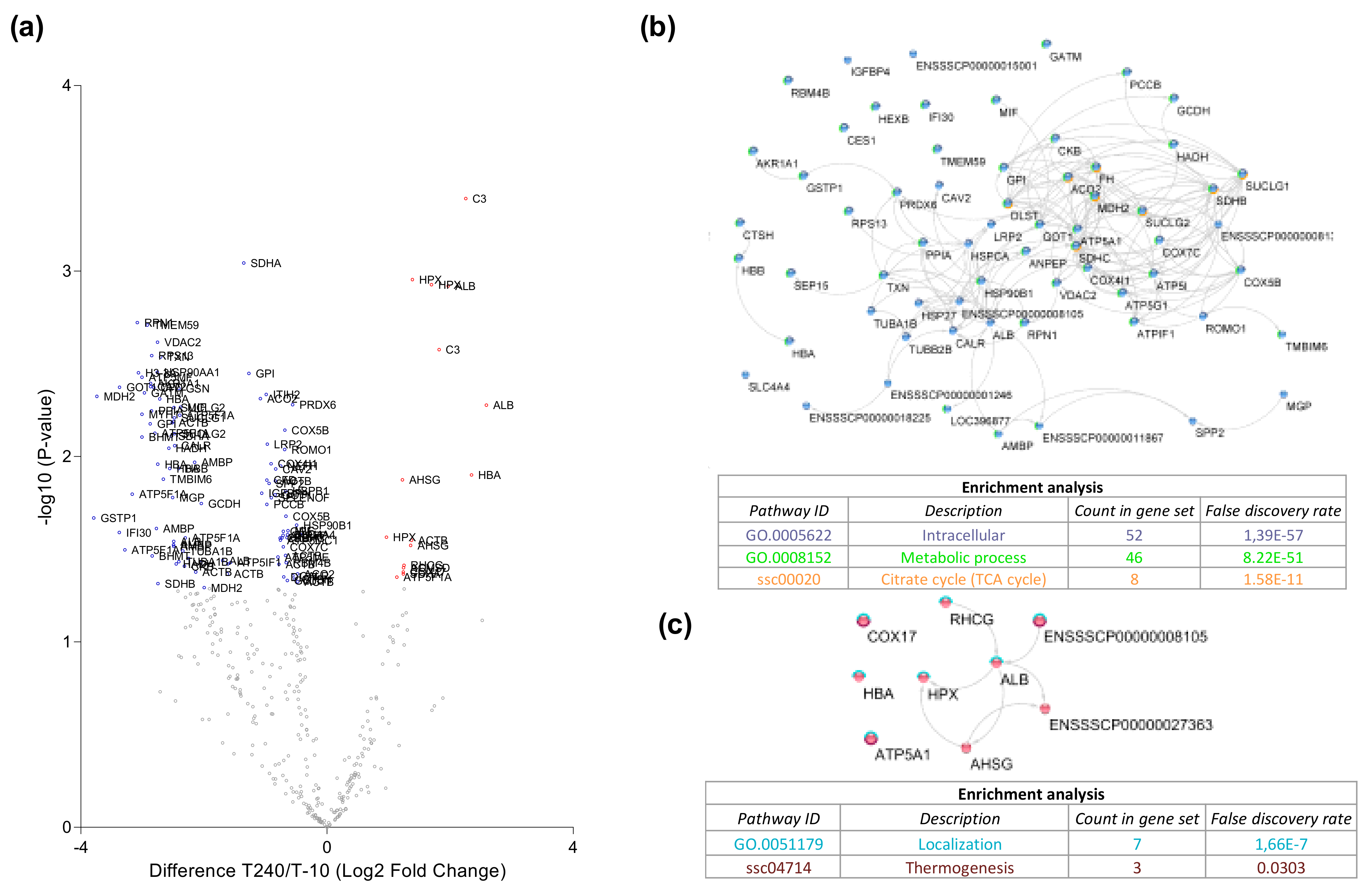
3.5. Protein Degradation during Ex Vivo Reperfusion
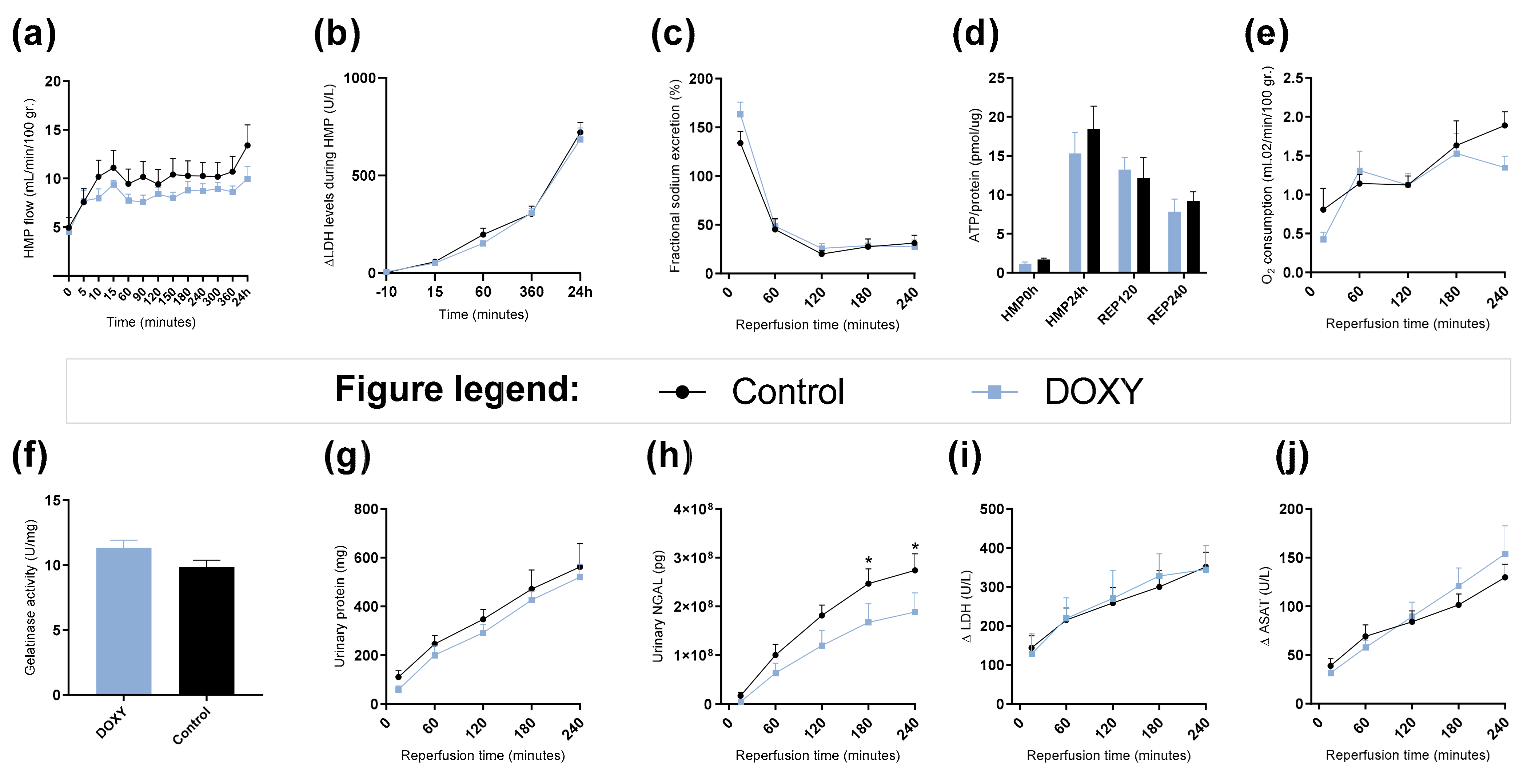
3.6. Renal Function and Injury Markers during HMP and Reperfusion
4. Discussion
4.1. Protein Secretion during Ex Vivo Reperfusion
4.2. Changes in Metabolic Pathways
4.3. Changes in Complement and Coagulation Cascades
4.4. Protein Degradation during Ex Vivo Reperfusion
4.5. Proteases as Possible Molecular Targets
4.6. Effect of Doxycycline
4.7. Clinical Implementation
4.8. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Perfusate Composition: | ||
| Heparinized and leukocyte depleted autologous blood | 500 mL | |
| Ringers lactate solution (Baxter) | 280 mL | |
| Amoxicillin/Clavulanic acid (1000 mg/200 mg) | 1 ampule | |
| 8.4% Sodium Bicarbonate (B. Braun) | 10 mL | |
| 5% glucose (Baxter) | 10 mL | |
| Dexamethasone (20 mg/mL) (Centrafarm) | 333 μL | |
| Mannitol (Merck) | 6 mg | |
| Creatinine (Merck) | 90 mg | |
| Sodium Nitroprusside (Merck) | 2 mg | |
| Infusion solution: 20 mL/h | ||
| Aminoplasmal (B. Braun) | 90 mL | |
| 8.4% Sodium Bicarbonate | 2.75 mL | |
| Insulin (100 IU/mL) (NovoRapid) | 0.186 mL | |
| Corrections based on blood gas values: | ||
| If glucose < 8 mmol/L, 5% glucose administrated to a concentration of 8 mmol/L. | ||
| If pH under 7.3, 8.4% sodium bicarbonate administrated using Henderson–Hasselbalch equation to obtain a pH of 7.35. | ||
| Renal function analysis: | ||
| Fractional sodium excretion | Creatinine and sodium concentrations were determined in perfusate samples and corresponding urine samples, in a routine fashion at the lab of clinical chemistry (UMCG). Calculated as: (urinary sodium levels × urinary flow)/(creatinine clearance × perfusate sodium levels). | |
| Oxygen consumption | Calculated using pO2 differences between arterial and venous sites, measured with a pH-blood gas analyzer ABL90 FLEX (Radiometer, Brønshøj, Denmark) and expressed as mL O2/min/100 r. according to trans-renal flow and kidney mass pre-reperfusion using the equation as described by Venema et al. 2019 [77]. | |
| ATP/Protein | Tissue samples collected after 30 min of warm ischemia, 24 h of HMP, 120 min of reperfusion and 240 min of reperfusion were stored in sonification solution (0.372 g EDTA in 130 mL H2O and NaOH, pH 10.9 + 370 mL 96% ethanol). A bioluminescence kit was used (Roche Diagnostics). Luminescence was measured using a luminometer (Packerd LumiCountä, IL, USA). The obtained ATP value was normalized to the total protein content using PierceTM BCA protein assay kit. The final ATP content was expressed as pmol ATP/μg protein. | |
| LDH & ASAT | Concentrations were measured in HMP and NMP perfusate in a routine fashion at the lab of clinical chemistry (UMCG). | |
| Urinary protein concentrations | Determined using Pierce™ BCA Protein Assay Kit (Thermo Scientific) according to manufacturer’s instructions. | |
| Urinary NGAL | Determined using an ELISA kit (Eurobio, Les Ulis, France) according to manufacturer’s instructions. | |
| MMP activity | Measured using the Gelatinase (Gelatin Degradation/Zymography) Assay Kit (BioVision, San Francisco, CA, USA) according to manufacturer’s instructions. | |
| Statistics | GraphPad Prism 7.02 (GraphPad Software, USA) was used for visualizing data and statistical analyses. Values are shown as means with corresponding standard error of the mean. Continuous variables were plotted over time. Statistical differences between groups for each timepoint were determined using 2 way ANOVA and Fisher’s LSD for multiple comparisons. The cut-off for statistical significance was set at p < 0.5. | |
References
- Schaubel, D.; Desmeules, M.; Mao, Y.; Jeffery, J.; Fenton, S. Survival experience among elderly end-stage renal disease patients. A controlled comparison of transplantation and dialysis. Transplantation 1995, 60, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Filiopoulos, V.; Boletis, J.N. Renal transplantation with expanded criteria donors: Which is the optimal immunosuppression? World J. Transplant. 2016, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- Saidi, R.F.; Elias, N.; Kawai, T.; Hertl, M.; Farrell, M.-L.; Goes, N.; Wong, W.; Hartono, C.; Fishman, J.A.; Kotton, C.N.; et al. Outcome of Kidney Transplantation Using Expanded Criteria Donors and Donation After Cardiac Death Kidneys: Realities and Costs. Am. J. Transplant. 2007, 7, 2769–2774. [Google Scholar] [CrossRef]
- Venkat, K.K.; Eshelman, A.K. The evolving approach to ethical issues in living donor kidney transplantation: A review based on illustrative case vignettes. Transplant. Rev. 2014, 28, 134–139. [Google Scholar] [CrossRef]
- Matas, A.J.; Gillingham, K.J.; Humar, A.; Kandaswamy, R.; Sutherland, D.E.R.; Payne, W.D.; Dunn, T.B.; Najarian, J.S. 2202 kidney transplant recipients with 10 years of graft function: What happens next? Am. J. Transplant 2008, 8, 2410–2419. Available online: http://www.ncbi.nlm.nih.gov/pubmed/18925907 (accessed on 12 October 2017). [CrossRef] [Green Version]
- Pascual, M.; Theruvath, T.; Kawai, T.; Tolkoff-Rubin, N.; Cosimi, A.B. Strategies to Improve Long-Term Outcomes after Renal Transplantation. N. Engl. J. Med. 2002, 346, 580–590. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11856798 (accessed on 12 October 2017). [CrossRef] [PubMed]
- Weight, S.C.; Bell, P.R.F.; Nicholson, M.L. Renal ischaemia-reperfusion injury. Br. J. Surg. 1996, 83, 162–170. [Google Scholar] [CrossRef]
- Nieuwenhuijs-Moeke, G.J.; Pischke, S.E.; Berger, S.P.; Sanders, J.S.F.; Pol, R.A.; Struys, M.M.R.F.; Ploeg, R.J.; Leuvenink, H.G.D. Ischemia and Reperfusion Injury in Kidney Transplantation: Relevant Mechanisms in Injury and Repair. J. Clin. Med. 2020, 9, 253. Available online: www.mdpi.com/journal/jcm (accessed on 5 March 2021). [CrossRef] [PubMed] [Green Version]
- Malek, M.; Nematbakhsh, M. Renal ischemia/reperfusion injury; from pathophysiology to treatment. J. Renal Inj Prev. 2015, 4, 20–27. [Google Scholar] [CrossRef]
- Nelson, K.K.; Melendez, J.A. Mitochondrial redox control of matrix metalloproteinases. Free Radic. Biol. Med. 2004, 37, 768–784. Available online: https://www.sciencedirect.com/science/article/pii/S0891584904004617?via%3Dihub#aep-section-id17 (accessed on 9 October 2018). [CrossRef] [PubMed]
- Dejonckheere, E.; Vandenbroucke, R.E.; Libert, C. Matrix metalloproteinases as drug targets in ischemia/reperfusion injury. Drug Discov. Today 2011, 16, 762–778. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21745586 (accessed on 25 September 2018). [CrossRef] [PubMed]
- Basile, D.P.; Fredrich, K.; Weihrauch, D.; Hattan, N.; Chilian, W.M. Angiostatin and matrix metalloprotease expression following ischemic acute renal failure. Am. J. Physiol. Physiol. 2004, 286, F893–F902. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15075185 (accessed on 25 September 2018). [CrossRef] [PubMed] [Green Version]
- Caron, A.; Desrosiers, R.R.; Langlois, S.; Béliveau, R. Ischemia–reperfusion injury stimulates gelatinase expression and activity in kidney glomeruli. Can. J. Physiol. Pharmacol. 2005, 83, 287–300. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15870843 (accessed on 25 September 2018). [CrossRef]
- Cavdar, Z.; Ozbal, S.; Celik, A.; Ergur, B.; Guneli, E.; Ural, C.; Camsari, T.; Guner, G. The effects of alpha-lipoic acid on MMP-2 and MMP-9 activities in a rat renal ischemia and re-perfusion model. Biotech. Histochem. 2014, 89, 304–314. Available online: http://www.tandfonline.com/doi/full/10.3109/10520295.2013.847498 (accessed on 8 June 2018). [CrossRef]
- Tan Kui, T.; Zheng, G.; Hsu, T.-T.; Wang, Y.; Lee, V.W.S.; Tian, X.; Wang, Y.; Cao, Q.; Wang, Y.; Harris, D.C.H. Macrophage Matrix Metalloproteinase-9 Mediates Epithelial-Mesenchymal Transition in Vitro in Murine Renal Tubular Cells. Am. J. Pathol. 2010, 176, 1256–1270. Available online: http://www.ncbi.nlm.nih.gov/pubmed/20075196 (accessed on 4 June 2018). [CrossRef] [PubMed] [Green Version]
- Kui Tan, T.; Zheng, G.; Hsu, T.-T.; Ra Lee, S.; Zhang, J.; Zhao, Y.; Tian, X.; Wang, Y.; Min Wang, Y.; Cao, Q.; et al. Matrix metalloproteinase-9 of tubular and macrophage origin contributes to the pathogenesis of renal fibrosis via macrophage recruitment through osteopontin cleavage. Lab. Investig. 2013, 93, 434–449. Available online: http://www.ncbi.nlm.nih.gov/pubmed/23358111 (accessed on 4 June 2018). [CrossRef] [Green Version]
- Novak, K.B.; Le, H.D.; Christison-Lagay, E.R.; Nose, V.; Doiron, R.J.; Moses, M.A.; Puder, M. Effects of metalloproteinase inhibition in a murine model of renal ischemia-reperfusion injury. Pediatric Res. 2010, 67, 257–262. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19915515 (accessed on 8 June 2018). [CrossRef] [PubMed] [Green Version]
- Roach, D.M.; Fitridge, R.A.; Laws, P.E.; Millard, S.H.; Varelias, A.; Cowled, P.A. Up-regulation of MMP-2 and MMP-9 Leads to Degradation of Type IV Collagen During Skeletal Muscle Reperfusion Injury; Protection by the MMP Inhibitor, Doxycycline. Eur. J. Vasc. Endovasc. Surg 2002, 23, 260–269. Available online: http://www.idealibrary.com (accessed on 16 June 2018). [CrossRef] [Green Version]
- Saglam, F.; Celik, A.; Tayfur, D.; Cavdar, Z.; Yilmaz, O.; Sargioglu, S.; Kolatan, E.; Oktay, G.; Camsari, T. Decrease in cell proliferation by an matrix metalloproteinase inhibitor, doxycycline, in a model of immune-complex nephritis. Nephrology 2010, 15, 560–567. Available online: http://www.ncbi.nlm.nih.gov/pubmed/20649877 (accessed on 16 June 2018). [CrossRef]
- Cortes, A.L.; Gonsalez, S.R.; Rioja, L.S.; Oliveira, S.S.C.; Santos, A.L.S.; Prieto, M.C.; Melo, P.A.; Lara, L.S. Protective outcomes of low-dose doxycycline on renal function of Wistar rats subjected to acute ischemia/reperfusion injury. Biochim. Biophys. Acta -Mol. Basis Dis. 2018, 1864, 102–114. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28987762 (accessed on 16 June 2018). [CrossRef]
- Moers, C.; Smits, J.M.; Maathuis, M.-H.J.; Treckmann, J.; Van Gelder, F.; Napieralski, B.P.; Van Kasterop-Kutz, M.; Homan Van Der Heide, J.J.; Squifflet, J.-P.; Van Heurn, E.; et al. Machine Perfusion or Cold Storage in Decreased-Donor Kidney Transplantation. N. Engl. J. Med. 2009, 360, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochmans, I.; Moers, C.; Smits, J.M.; Leuvenink, H.G.D.; Treckmann, J.; Paul, A.; Rahmel, A.; Squifflet, J.-P.; van Heurn, E.; Monbaliu, D.; et al. Machine Perfusion Versus Cold Storage for the Preservation of Kidneys Donated After Cardiac Death. Ann. Surg. 2010, 252, 756–764. [Google Scholar] [CrossRef]
- Jochmans, I.; Brat, A.; Davies, L.; Hofker, H.S.; van de Leemkolk, F.E.M.; Leuvenink, H.G.D.; Knight, S.R.; Pirenne, J.; Ploeg, R.J.; Abramowicz, D.; et al. Oxygenated versus standard cold perfusion preservation in kidney transplantation (COMPARE): A randomised, double-blind, paired, phase 3 trial. Lancet 2020, 396, 1653–1662. Available online: www.eurotransplant.org (accessed on 11 March 2021). [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. Available online: https://thermo.flexnetoperations.com/control/ (accessed on 7 May 2021). [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. Available online: http://www.nature.com/articles/nmeth.3901 (accessed on 17 March 2020). [CrossRef]
- Weng, S.S.H.; Demir, F.; Ergin, E.K.; Dirnberger, S.; Uzozie, A.; Tuscher, D.; Nierves, L.; Tsui, J.; Huesgen, P.F.; Lange, P.F. Sensitive Determination of Proteolytic Proteoforms in Limited Microscale Proteome Samples. Mol. Cell. Proteom. 2019, 18, 2335–2347. Available online: http://www.ncbi.nlm.nih.gov/pubmed/31471496 (accessed on 17 March 2020). [CrossRef] [PubMed] [Green Version]
- Fye, H.K.S.; Mrosso, P.; Bruce, L.; Thézénas, M.L.; Davis, S.; Fischer, R.; Rwegasira, G.L.; Makani, J.; Kessler, B.M. A robust mass spectrometry method for rapid profiling of erythrocyte ghost membrane proteomes. Clin. Proteom. 2018, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. Available online: http://www.ncbi.nlm.nih.gov/pubmed/27924014 (accessed on 20 March 2018). [CrossRef]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. Available online: https://academic.oup.com/nar/article-lookup/doi/10.1093/nar/gkw1138 (accessed on 5 April 2018). [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. Available online: https://pubmed.ncbi.nlm.nih.gov/14597658/ (accessed on 6 May 2021). [CrossRef]
- Fortelny, N.; Yang, S.; Pavlidis, P.; Lange, P.F.; Overall, C.M. Proteome TopFIND 3.0 with TopFINDer and PathFINDer: Database and analysis tools for the association of protein termini to pre- and post-translational events. Nucleic Acids Res. 2015, 43, D290–D297. Available online: http://academic.oup.com/nar/article/43/D1/D290/2435434/Proteome-TopFIND-30-with-TopFINDer-and-PathFINDer (accessed on 6 May 2018). [CrossRef] [Green Version]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. Available online: http://academic.oup.com/nar/article/46/D1/D624/4626772 (accessed on 6 May 2018). [CrossRef] [PubMed]
- Dinis, P.; Nunes, P.; Marconi, L.; Furriel, F.; Parada, B.; Moreira, P.; Figueiredo, A.; Bastos, C.; Roseiro, A.; Dias, V.; et al. Kidney Retransplantation: Removal or Persistence of the Previous Failed Allograft? Transplant. Proc. 2014, 46, 1730–1734. Available online: http://www.ncbi.nlm.nih.gov/pubmed/25131023 (accessed on 6 October 2017). [CrossRef] [PubMed]
- Anderson, N.G.; Anderson, N.L. Twenty years of two-dimensional electrophoresis: Past, present and future. Electrophoresis 1996, 17, 443–453. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8740157 (accessed on 24 October 2017). [CrossRef] [PubMed]
- Graves, P.R.; Haystead, T.A.J. Molecular biologist’s guide to proteomics. Microbiol. Mol. Biol. Rev. 2002, 66, 39–63. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11875127 (accessed on 24 October 2017). [CrossRef] [PubMed] [Green Version]
- López-Otín, C.; Overall, C.M. Protease degradomics: A new challenge for proteomics. Nat. Rev. Mol. Cell Biol. 2002, 3, 509–519. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12094217 (accessed on 22 October 2017). [CrossRef]
- Coppo, R. Proteasome inhibitors in progressive renal diseases. Nephrol. Dial. Transpl. 2014, 29, 25–30. Available online: https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/ndt/29/suppl_1/10.1093_ndt_gft271/3/gft271.pdf?Expires=1509553937&Signature=bRec1EGIsaSFJxmA3Q9n7Y-sTHt2qe4VAn80-YBucmq5bd~OzjsbR36R~aPHCsWisFxJn-k0LqbFBVTyGbBqZaYcuv4QRfMogZ-61sCYHrDk60Re9yPW4WotXPji3St1AqtvA4jIjkLlq1rIbtP4yDHk26RNbac5UQipA1JYK7IU0sTw~ozGnvlqZLPcCcdzBrc9QpN-yQP16chUB2yNC-R-pBjnQEkIWoi6M84gZqsBG886G1eb075se0jtjuEVy5v41sk9-P7CJHtD94t7O3BGuzaVa9Hng7w~v0zit6JxBXWBpRtNx80vAcckbYI7vT5Lyjcf7HsVetl32I9UYw__&Key-Pair-Id=APKAIUCZBIA4LVPAVW3Q (accessed on 31 October 2017). [CrossRef] [Green Version]
- Abbenante, G.; Fairlie, D.P. Protease inhibitors in the clinic. Med. Chem. 2005, 1, 71–104. Available online: http://www.ncbi.nlm.nih.gov/pubmed/16789888 (accessed on 24 October 2017). [CrossRef]
- Sigdel, T.K.; Lee, S.; Sarwal, M.M. Profiling the proteome in renal transplantation. Proteomics. Clin. Appl. 2011, 5, 269–280. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21520424 (accessed on 24 October 2017). [CrossRef]
- Späth, M.R.; Bartram, M.P.; Palacio-Escat, N.; Hoyer, K.J.R.; Debes, C.; Demir, F.; Schroeter, C.B.; Mandel, A.M.; Grundmann, F.; Ciarimboli, G.; et al. The proteome microenvironment determines the protective effect of preconditioning in cisplatin-induced acute kidney injury. Kidney Int. 2019, 95, 333–349. [Google Scholar] [CrossRef]
- Amer, H.; Fidler, M.E.; Myslak, M.; Morales, P.; Kremers, W.K.; Larson, T.S.; Stegall, M.D.; Cosio, F.G. Proteinuria after kidney transplantation, relationship to allograft histology and survival. Am. J. Transplant. 2007, 7, 2748–2756. Available online: https://pubmed.ncbi.nlm.nih.gov/17941956/ (accessed on 26 November 2020). [CrossRef]
- Coskun, A.; Baykal, A.T.; Kazan, D.; Akgoz, M.; Senal, M.O.; Berber, I.; Titiz, I.; Bilsel, G.; Kilercik, H.; Karaosmanoglu, K.; et al. Proteomic Analysis of Kidney Preservation Solutions Prior to Renal Transplantation. PLoS ONE 2016, 11, e0168755. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28036361 (accessed on 25 April 2018). [CrossRef] [PubMed] [Green Version]
- Devarajan, P. Update on Mechanisms of Ischemic Acute Kidney Injury. J. Am. Soc. Nephrol. 2006, 17, 1503–1520. Available online: http://www.ncbi.nlm.nih.gov/pubmed/16707563 (accessed on 21 March 2018). [CrossRef] [Green Version]
- Diepenhorst, G.M.P.; Van Gulik, T.M.; Hack, C.E. Complement-mediated ischemia-reperfusion injury: Lessons learned from animal and clinical studies. Ann. Surg. 2009, 249, 889–899. [Google Scholar] [CrossRef]
- McCaughan, J.A.; O’Rourke, D.M.; Courtney, A.E. The complement cascade in kidney disease: From sideline to center stage. Am. J. Kidney Dis. 2013, 62, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Gasque, P. Complement: A unique innate immune sensor for danger signals. Mol. Immunol. 2004, 41, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, L.L.; Spraakman, N.A.; Brat, A.; Huang, H.; Thorne, A.M.; Bonham, S.; van Balkom, B.W.M.; Ploeg, R.J.; Kessler, B.M.; Leuvenink, H.G.D. Proteomic analysis of machine perfusion solution from brain dead donor kidneys reveals that elevated complement, cytoskeleton and lipid metabolism proteins are associated with 1-year outcome. Transpl. Int. 2021, 34, 1618–1629. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/tri.13984 (accessed on 9 September 2021). [CrossRef] [PubMed]
- Stowell, S.R.; Winkler, A.M.; Maier, C.L.; Arthur, C.M.; Smith, N.H.; Girard-Pierce, K.R.; Cummings, R.D.; Zimring, J.C.; Hendrickson, J.E. Initiation and Regulation of Complement during Hemolytic Transfusion Reactions. Clin. Dev. Immunol. 2012, 2012, 12. [Google Scholar] [CrossRef] [Green Version]
- Danobeitia, J.S.; Djamali, A.; Fernandez, L.A. The role of complement in the pathogenesis of renal ischemia-reperfusion injury and fibrosis. Fibrogenes. Tissue Repair 2014, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Debigaré, R.; Price, S.R. Proteolysis, the ubiquitin-proteasome system, and renal diseases. Am. J. Physiol. Renal Physiol. 2003, 285, F1–F8. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12788783 (accessed on 29 October 2017). [CrossRef] [Green Version]
- Rajan, V.; Mitch, W.E. Ubiquitin, proteasomes and proteolytic mechanisms activated by kidney disease. BBA -Mol. Basis Dis. 2008, 1782, 795–799. Available online: http://nobelprize.org/chemistry/laureates/2004/ (accessed on 7 May 2021). [CrossRef] [Green Version]
- Sorimachi, H.; Ishiura, S.; Suzuki, K. Structure and physiological function of calpains. Biochem. J. 1997, 328, 721–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saido, T.C.; Sorimachi, H.; Suzuki, K. Calpain: New perspectives in molecular diversity and physiological-pathological involvement. FASEB J. 1994, 8, 814–822. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1096/fasebj.8.11.8070630 (accessed on 26 June 2020). [CrossRef] [Green Version]
- Ma, H.; Yang, H.Q.; Takano, E.; Lee, W.J.; Hatanaka, M.; Maki, M. Requirement of different subdomains of calpastatin for calpain inhibition and for binding to calmodulin-like domains. J. Biochem. 1993, 113, 591–599. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8340353 (accessed on 6 May 2018). [CrossRef] [PubMed]
- Saido, T.C.; Kawashima, S.; Tani, E.; Yokota, M. Up- and down-regulation of calpain inhibitor polypeptide, calpastatin, in postischemic hippocampus. Neurosci. Lett. 1997, 227, 75–78. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9180207 (accessed on 6 May 2018). [CrossRef]
- Sorimachi, Y.; Harada, K.; Saido, T.C.; Ono, T.; Kawashima, S.-I.; Yoshida, K.-I. Downregulation of Calpastatin in Rat Heart after Brief Ischemia and Reperfusion1. J. Biochem 1997, 122, 743–748. Available online: https://www.jstage.jst.go.jp/article/biochemistry1922/122/4/122_4_743/_pdf (accessed on 6 May 2018). [CrossRef]
- Chatterjee, P.K.; Brown, P.A.J.; Cuzzocrea, S.; Zacharowski, K.; Stewart, K.N.; Mota-Filipe, H.; Mcdonald, M.C.; Thiemermann, C. Calpain inhibitor-1 reduces renal ischemia/reperfusion injury in the rat. Kidney Int. 2001, 59, 2073–2083. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, R.H.; Kresse, J.-C.; Farmer, L.K.; Thézénas, M.L.; Kessler, B.M.; Lindeman, J.H.N.; Sharples, E.J.; Welsh, G.I.; Nørregaard, R.; Ploeg, R.J.; et al. Cytoskeletal protein degradation in brain death donor kidneys associates with adverse post-transplant outcomes. Am. J. Transplant. 2021. Published ahead of print. [Google Scholar] [CrossRef]
- Shi, Y.; Melnikov, V.Y.; Schrier, R.W.; Edelstein, C.L. Downregulation of the calpain inhibitor protein calpastatin by caspases during renal ischemia-reperfusion. Am. J. Physiol. Renal Physiol. 2000, 279, F509–F517. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10966930 (accessed on 9 June 2018). [CrossRef]
- Arnold, P.; Otte, A.; Becker-Pauly, C. Meprin metalloproteases: Molecular regulation and function in inflammation and fibrosis. Biochim. Biophys. Acta -Mol. Cell Res. 2017, 1864, 2096–2104. [Google Scholar] [CrossRef]
- Kaushal, G.P.; Haun, R.S.; Herzog, C.; Shah, S.V. Meprin A metalloproteinase and its role in acute kidney injury. Am. J. Physiol. -Ren. Physiol. 2013, 304, F1150. [Google Scholar] [CrossRef] [Green Version]
- Mishra, J.; Dent, C.; Tarabishi, R.; Mitsnefes, M.M.; Ma, Q.; Kelly, C.; Ruff, S.M.; Zahedi, K.; Shao, M.; Bean, J.; et al. Neutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renal injury after cardiac surgery. Lancet 2005, 365, 1231–1238. Available online: https://www.sciencedirect.com/science/article/pii/S014067360574811X?via%3Dihub (accessed on 4 March 2019). [CrossRef]
- Parikh, C.R.; Hall, I.E.; Bhangoo, R.S.; Ficek, J.; Abt, P.L.; Thiessen-Philbrook, H.; Lin, H.; Bimali, M.; Murray, P.T.; Rao, V.; et al. Associations of Perfusate Biomarkers and Pump Parameters With Delayed Graft Function and Deceased Donor Kidney Allograft Function. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2016, 16, 1526–1539. [Google Scholar] [CrossRef]
- Mishra, J.; Ma, Q.; Prada, A.; Mitsnefes, M.; Zahedi, K.; Yang, J.; Barasch, J.; Devarajan, P. Identification of neutrophil gelatinase-associated lipocalin as a novel early urinary biomarker for ischemic renal injury. J. Am. Soc. Nephrol. 2003, 14, 2534–2543. Available online: http://www.ncbi.nlm.nih.gov/pubmed/14514731 (accessed on 4 March 2019). [CrossRef] [Green Version]
- Saito, H.; Tanaka, T.; Tanaka, S.; Higashijima, Y.; Yamaguchi, J.; Sugahara, M.; Ito, M.; Uchida, L.; Hasegawa, S.; Wakashima, T.; et al. Persistent expression of neutrophil gelatinase-associated lipocalin and M2 macrophage markers and chronic fibrosis after acute kidney injury. Physiol. Rep. 2018, 6, e13707. Available online: http://www.ncbi.nlm.nih.gov/pubmed/29845768 (accessed on 4 March 2019). [CrossRef] [PubMed]
- Nakagawa, S.; Nishihara, K.; Miyata, H.; Shinke, H.; Tomita, E.; Kajiwara, M.; Matsubara, T.; Iehara, N.; Igarashi, Y.; Yamada, H.; et al. Molecular Markers of Tubulointerstitial Fibrosis and Tubular Cell Damage in Patients with Chronic Kidney Disease. PLoS ONE 2015, 10, e0136994. [Google Scholar] [CrossRef] [Green Version]
- Moser, M.A.J.; Arcand, S.; Lin, H.-B.; Wojnarowicz, C.; Sawicka, J.; Banerjee, T.; Luo, Y.; Beck, G.R.; Luke, P.P.; Sawicki, G. Protection of the Transplant Kidney from Preservation Injury by Inhibition of Matrix Metalloproteinases. PLoS ONE 2016, 11, e0157508. [Google Scholar] [CrossRef] [Green Version]
- Decaneto, E.; Suladze, S.; Rosin, C.; Havenith, M.; Lubitz, W.; Winter, R. Pressure and Temperature Effects on the Activity and Structure of the Catalytic Domain of Human MT1-MMP. Biophys. J. 2015, 109, 2371–2381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Leeuwen, L.L.; Leuvenink, H.G.D.; Olinga, P.; Ruigrok, M.J.R. Shifting paradigms for suppressing fibrosis in kidney transplants: Supplementing perfusion solutions with antifibrotic drugs. Front. Med. 2021, 8, 2917. [Google Scholar] [CrossRef]
- Maassen, H.; Hendriks, K.D.W.; Venema, L.H.; Henning, R.H.; Hofker, S.H.; van Goor, H.; Leuvenink, H.G.D.; Coester, A.M. Hydrogen sulphide-induced hypometabolism in human-sized porcine kidneys. PLoS ONE 2019, 14, e0225152. [Google Scholar] [CrossRef] [PubMed]
- Pool, M.; Eertman, T.; Parraga, J.S.; ’t Hart, N.; van Rhijn, M.R.; Eijken, M.; Jespersen, B.; Reinders, M.; Hoogduijn, M.; Ploeg, R.; et al. Infusing mesenchymal stromal cells into porcine kidneys during normothermic machine perfusion: Intact MSCs can be traced and localised to Glomeruli. Int. J. Mol. Sci. 2019, 20, 3607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijink, T.M.; Venema, L.H.; Posma, R.A.; de Vries, N.J.; Westerkamp, A.C.; Ottens, P.J.; Touw, D.J.; Nijsten, M.W.; Leuvenink, H.G.D. Metformin Preconditioning and Postconditioning to Reduce Ischemia Reperfusion Injury in an Isolated Ex Vivo Rat and Porcine Kidney Normothermic Machine Perfusion Model. Clin. Transl. Sci. 2020, 14, 222–230. [Google Scholar] [CrossRef]
- Posma, R.A.; Venema, L.H.; Huijink, T.M.; Westerkamp, A.C.; Wessels, A.M.A.; de Vries, N.J.; Doesburg, F.; Roggeveld, J.; Ottens, P.J.; Touw, D.J.; et al. Increasing metformin concentrations and its excretion in both rat and porcine ex vivo normothermic kidney perfusion model. BMJ Open Diabetes Res. Care 2020, 8, e000816. [Google Scholar] [CrossRef]
- Hendriks, K.D.W.; Brüggenwirth, I.M.A.; Maassen, H.; Gerding, A.; Bakker, B.; Porte, R.J.; Henning, R.H.; Leuvenink, H.G.D. Renal temperature reduction progressively favors mitochondrial ROS production over respiration in hypothermic kidney preservation. J. Transl. Med. 2019, 17, 265. [Google Scholar] [CrossRef]
- Venema, L.H.; van Leeuwen, L.L.; Posma, R.A.; van Goor, H.; Ploeg, R.J.; Hannaert, P.; Hauet, T.; Minor, T.; Leuvenink, H.G.D. Impact of Red Blood Cells on Function and Metabolism of Porcine Deceased Donor Kidneys during Normothermic Machine Perfusion. Transplantation 2021. Published ahead of print. Available online: https://journals.lww.com/transplantjournal/Fulltext/9000/Impact_of_Red_Blood_Cells_on_Function_and.95170.aspx (accessed on 9 September 2021).
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. Available online: https://pubmed.ncbi.nlm.nih.gov/30395289/ (accessed on 18 June 2021). [CrossRef] [PubMed]
- Venema, L.H.; Brat, A.; Moers, C.; ’t Hart, N.A.; Ploeg, R.J.; Hannaert, P.; Minor, T.; Leuvenink, A.H.G.D. Effects of Oxygen During Long-term Hypothermic Machine Perfusion in a Porcine Model of Kidney Donation After Circulatory Death. Transplantation 2019, 103, 2057–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]





| Cysteine Proteases | Metallo Proteases | Serine Proteases |
|---|---|---|
| CTSB | FOLH1 | PLG |
| USP37 | ANPEP | APEH |
| CTSH | MMP1 | DPP4 |
| CAPN1 | THOP1 | CFD |
| CAPN2 | NLN | HP |
| UCHL3 | ENPEP | ESD |
| CTSL | ||
| UCHL1 |
| Doxycycline T-10 | Control T-10 | ||
|---|---|---|---|
| Accession Number Gene Name | Meprin α/β Metalloproteinase Cleavage Sites | Accession Number Gene Name | Meprin α/β Metalloproteinase Cleavage Sites |
| P10809 HSPD1 |  | P10809 HSPD1 |  |
| Q99497 PARK7 |  | P37802 TAGLN2 |  |
| Q15651 HMGN |  | P11142 HSPA8 |  |
| P22626 HNRNPA2B1 |  | P08670 VIM |  |
| Accession number Gene name | Serine protease HTRA2 Cleavage sites | P06396 GSN |  |
| P68371 TUBB4B |  | P14866 HNRNPL |  |
| P63267 ACTG2 |  | Accession number Gene name | Serine protease HTRA2 cleavage sites |
| P68366 TUBA4A |  | P68363 TUBA1B |  |
| Accession number Gene name | Cathepsin S cleavage sites | P68371 TUBB4B |  |
| P60709 ACTB |  | Accession number Gene name | Caspase 3 cleavage sites |
| O60749 |  | P63267 ACTG2 |  |
| Accession number Gene name | Granzyme M cleavage sites | Accession number Gene name | Granzyme M cleavage sites |
| O75367 H2AFY |  | P16402 HIST1H1D |  |
| P16402 HIST1H1D |  | P16402 HIST1H1D |  |
| P60709 ACTB |  | P16402 |  |
| Accession number Gene name | Cathepsin B cleavage sites | Accession number Gene name | Granzyme B cleavage sites |
| P52272 HNRNPM |  | P68104 EEF1A1 |  |
| Accession number Gene name | Mitochondrial-processing peptidase β cleavage sites | Accession number Gene name | Cathepsin D cleavage sites |
| Q99643 SDHC |  | P68871 HBB |  |
| Accession number Gene name | Tripeptidyl-peptidase 1 cleavage sites | ||
| O14773 TPP1 |  | Doxycycline T240 | |
| Accession number Gene name | Cathepsin L1 cleavage sites | Accession number Gene name | Meprin α/β metalloproteinase cleavage sites |
| P53634 CTSC |  | P14866 HNRNPL |  |
| Accession number Gene name | Matrix metalloprotease 11 cleavage sites | Accession number Gene name | Cathepsin S cleavage sites |
| P62937 |  | O00193 SMAP |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Leeuwen, L.; Venema, L.H.; Heilig, R.; Leuvenink, H.G.D.; Kessler, B.M. Doxycycline Alters the Porcine Renal Proteome and Degradome during Hypothermic Machine Perfusion. Curr. Issues Mol. Biol. 2022, 44, 559-577. https://doi.org/10.3390/cimb44020039
van Leeuwen L, Venema LH, Heilig R, Leuvenink HGD, Kessler BM. Doxycycline Alters the Porcine Renal Proteome and Degradome during Hypothermic Machine Perfusion. Current Issues in Molecular Biology. 2022; 44(2):559-577. https://doi.org/10.3390/cimb44020039
Chicago/Turabian Stylevan Leeuwen, Leonie, Leonie H. Venema, Raphael Heilig, Henri G. D. Leuvenink, and Benedikt M. Kessler. 2022. "Doxycycline Alters the Porcine Renal Proteome and Degradome during Hypothermic Machine Perfusion" Current Issues in Molecular Biology 44, no. 2: 559-577. https://doi.org/10.3390/cimb44020039