

Suppressive Effects of 4-(Phenylsulfanyl) Butan-2-One on CCL-1 Production via Histone Acetylation in Monocytes
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Preparation
2.2. Cell Viability
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Western Blotting
2.5. Chromatin Immunoprecipitation (ChIP)
2.6. Statistical Analyses
3. Results
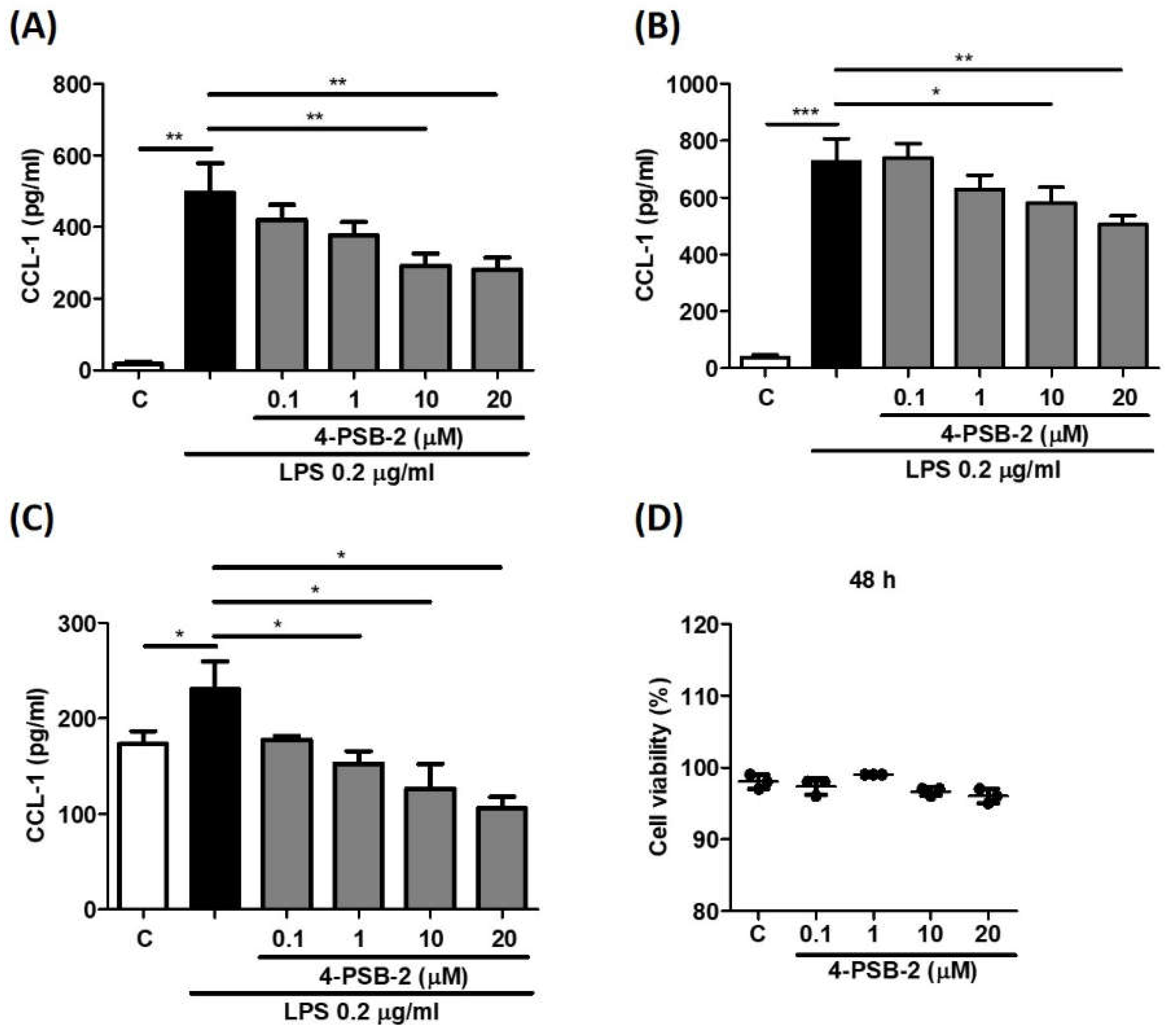
3.1. 4-PSB-2 Suppressed LPS-Induced CCL-1 Production in THP-1 Cells
3.2. The Experimental Concentration of 4-PSB-2 Had No Cytotoxic Effects on THP-1 Cells
3.3. 4-PSB-2 Suppressed CCL-1 Production in THP-1-Cell-Derived Macrophage and Human CD14+ Cells
3.4. 4-PSB-2 Suppressed CCL-1 Expression through MAPK and NF-κB Pathways
3.5. 4-PSB-2 Suppressed LPS-Induced CCL-1 via Histone Acetylation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jean, Y.H.; Chen, W.F.; Sung, C.S.; Duh, C.Y.; Huang, S.Y.; Lin, C.S.; Tai, M.H.; Tzeng, S.F.; Wen, Z.H. Capnellene, a natural marine compound derived from soft coral, attenuates chronic constriction injury-induced neuropathic pain in rats. Br. J. Pharm. 2009, 158, 713–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Huang, S.Y.; Jean, Y.H.; Chen, W.F.; Sung, C.S.; Kao, E.S.; Wang, H.M.; Chakraborty, C.; Duh, C.Y.; Wen, Z.H. Intrathecal lemnalol, a natural marine compound obtained from Formosan soft coral, attenuates nociceptive responses and the activity of spinal glial cells in neuropathic rats. Behav. Pharm. 2011, 22, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Chakraborty, C.; Sung, C.S.; Feng, C.W.; Jean, Y.H.; Lin, Y.Y.; Hung, H.C.; Huang, T.Y.; Huang, S.Y.; Su, T.M.; et al. Neuroprotection by marine-derived compound, 11-dehydrosinulariolide, in an in vitro Parkinson’s model: A promising candidate for the treatment of Parkinson’s disease. Naunyn Schmiedebergs Arch. Pharm. 2012, 385, 265–275. [Google Scholar] [CrossRef]
- Huang, S.Y.; Chen, N.F.; Chen, W.F.; Hung, H.C.; Lee, H.P.; Lin, Y.Y.; Wang, H.M.; Sung, P.J.; Sheu, J.H.; Wen, Z.H. Sinularin from indigenous soft coral attenuates nociceptive responses and spinal neuroinflammation in carrageenan-induced inflammatory rat model. Mar. Drugs 2012, 10, 1899–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.F.; Huang, S.Y.; Lu, C.H.; Chen, C.L.; Feng, C.W.; Chen, C.H.; Hung, H.C.; Lin, Y.Y.; Sung, P.J.; Sung, C.S.; et al. Flexibilide obtained from cultured soft coral has anti-neuroinflammatory and analgesic effects through the upregulation of spinal transforming growth factor-beta1 in neuropathic rats. Mar. Drugs 2014, 12, 3792–3817. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.H.; Chao, C.H.; Wu, M.H.; Sheu, J.H. A neuroprotective sulfone of marine origin and the in vivo anti-inflammatory activity of an analogue. Eur. J. Med. Chem. 2010, 45, 5998–6004. [Google Scholar] [CrossRef]
- Chien, J.Y.; Sheu, J.H.; Wen, Z.H.; Tsai, R.K.; Huang, S.P. Neuroprotective effect of 4-(Phenylsulfanyl)butan-2-one on optic nerve crush model in rats. Exp. Eye Res. 2016, 143, 148–157. [Google Scholar] [CrossRef]
- Wu, S.Y.; Wang, H.M.; Wen, Y.S.; Liu, W.; Li, P.H.; Chiu, C.C.; Chen, P.C.; Huang, C.Y.; Sheu, J.H.; Wen, Z.H. 4-(Phenylsulfanyl)butan-2-One Suppresses Melanin Synthesis and Melanosome Maturation In Vitro and In Vivo. Int. J. Mol. Sci. 2015, 16, 20240–20257. [Google Scholar] [CrossRef] [Green Version]
- Varinthra, P.; Huang, S.P.; Chompoopong, S.; Wen, Z.H.; Liu, I.Y. 4-(Phenylsulfanyl) Butan-2-One Attenuates the Inflammatory Response Induced by Amyloid-beta Oligomers in Retinal Pigment Epithelium Cells. Mar. Drugs 2020, 19, 1. [Google Scholar] [CrossRef]
- Busse, W.W.; Lemanske, R.F., Jr. Asthma. N. Engl. J. Med. 2001, 344, 350–362. [Google Scholar] [CrossRef]
- Smit, J.J.; Lukacs, N.W. A closer look at chemokines and their role in asthmatic responses. Eur. J. Pharm. 2006, 533, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Panina-Bordignon, P.; Papi, A.; Mariani, M.; Di Lucia, P.; Casoni, G.; Bellettato, C.; Buonsanti, C.; Miotto, D.; Mapp, C.; Villa, A.; et al. The C-C chemokine receptors CCR4 and CCR8 identify airway T cells of allergen-challenged atopic asthmatics. J. Clin. Investig. 2001, 107, 1357–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, R.C.; Becker, A.B.; Kozyrskyj, A.L.; Hayglass, K.T. Altered epigenetic regulation and increasing severity of bronchial hyperresponsiveness in atopic asthmatic children. J. Allergy Clin. Immunol. 2009, 124, 1116–1118. [Google Scholar] [CrossRef]
- Harb, H.; Raedler, D.; Ballenberger, N.; Bock, A.; Kesper, D.A.; Renz, H.; Schaub, B. Childhood allergic asthma is associated with increased IL-13 and FOXP3 histone acetylation. J. Allergy Clin. Immunol. 2015, 136, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; von Bulow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. Histone modifications and their role in epigenetics of atopy and allergic diseases. Allergy Asthma Clin. Immunol. 2018, 14, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, C.H.; Ko, Y.C.; Yang, S.N.; Chu, Y.T.; Wang, W.L.; Huang, S.K.; Chen, H.N.; Wei, W.J.; Jong, Y.J.; Hung, C.H. Effects of PGI2 analogues on Th1- and Th2-related chemokines in monocytes via epigenetic regulation. J. Mol. Med. 2011, 89, 29–41. [Google Scholar] [CrossRef] [PubMed]
- N’Diaye, M.; Le Ferrec, E.; Lagadic-Gossmann, D.; Corre, S.; Gilot, D.; Lecureur, V.; Monteiro, P.; Rauch, C.; Galibert, M.D.; Fardel, O. Aryl hydrocarbon receptor- and calcium-dependent induction of the chemokine CCL1 by the environmental contaminant benzo[a]pyrene. J. Biol. Chem. 2006, 281, 19906–19915. [Google Scholar] [CrossRef] [Green Version]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Palmqvist, C.; Wardlaw, A.J.; Bradding, P. Chemokines and their receptors as potential targets for the treatment of asthma. Br. J. Pharm. 2007, 151, 725–736. [Google Scholar] [CrossRef]
- Rojas-Dotor, S.; Segura-Mendez, N.H.; Miyagui-Namikawa, K.; Mondragon-Gonzalez, R. Expression of resistin, CXCR3, IP-10, CCR5 and MIP-1alpha in obese patients with different severity of asthma. Biol. Res. 2013, 46, 13–20. [Google Scholar] [CrossRef]
- Wang, W.L.; Kuo, C.H.; Chu, Y.T.; Huang, C.H.; Lam, K.P.; Huang, S.K.; Jong, Y.J.; Kuo, Y.T.; Hung, C.H. Prostaglandin I(2) analogues suppress TNF-alpha expression in human monocytes via mitogen-activated protein kinase pathway. Inflamm. Res. 2011, 60, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.H.; Suen, J.L.; Hua, Y.M.; Chiang, W.; Chang, H.C.; Chen, C.N.; Jong, Y.J. Suppressive effects of ketotifen on Th1- and Th2-related chemokines of monocytes. Pediatr. Allergy Immunol. 2007, 18, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Lin, C.H.; Yang, S.N.; Huang, M.Y.; Chen, H.L.; Kuo, P.L.; Hsu, Y.L.; Huang, S.K.; Jong, Y.J.; Wei, W.J.; et al. Effect of prostaglandin I2 analogs on cytokine expression in human myeloid dendritic cells via epigenetic regulation. Mol. Med. 2012, 18, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Jean, Y.H.; Chen, W.F.; Duh, C.Y.; Huang, S.Y.; Hsu, C.H.; Lin, C.S.; Sung, C.S.; Chen, I.M.; Wen, Z.H. Inducible nitric oxide synthase and cyclooxygenase-2 participate in anti-inflammatory and analgesic effects of the natural marine compound lemnalol from Formosan soft coral Lemnalia cervicorni. Eur. J. Pharm. 2008, 578, 323–331. [Google Scholar] [CrossRef]
- Chao, C.H.; Wen, Z.H.; Wu, Y.C.; Yeh, H.C.; Sheu, J.H. Cytotoxic and anti-inflammatory cembranoids from the soft coral Lobophytum crassum. J. Nat. Prod. 2008, 71, 1819–1824. [Google Scholar] [CrossRef]
- Tseng, Y.J.; Wen, Z.H.; Dai, C.F.; Chiang, M.Y.; Sheu, J.H. Nanolobatolide, a new C18 metabolite from the Formosan soft coral Sinularia nanolobata. Org. Lett. 2009, 11, 5030–5032. [Google Scholar] [CrossRef]
- Varinthra, P.; Ganesan, K.; Huang, S.P.; Chompoopong, S.; Eurtivong, C.; Suresh, P.; Wen, Z.H.; Liu, I.Y. The 4-(Phenylsulfanyl) butan-2-one Improves Impaired Fear Memory Retrieval and Reduces Excessive Inflammatory Response in Triple Transgenic Alzheimer’s Disease Mice. Front. Aging Neurosci. 2021, 13, 615079. [Google Scholar] [CrossRef]
- Leung, T.F.; Wong, G.W.; Ko, F.W.; Lam, C.W.; Fok, T.F. Increased macrophage-derived chemokine in exhaled breath condensate and plasma from children with asthma. Clin. Exp. Allergy 2004, 34, 786–791. [Google Scholar] [CrossRef]
- Sironi, M.; Martinez, F.O.; D’Ambrosio, D.; Gattorno, M.; Polentarutti, N.; Locati, M.; Gregorio, A.; Iellem, A.; Cassatella, M.A.; Van Damme, J.; et al. Differential regulation of chemokine production by Fcgamma receptor engagement in human monocytes: Association of CCL1 with a distinct form of M2 monocyte activation (M2b, Type 2). J. Leukoc. Biol. 2006, 80, 342–349. [Google Scholar] [CrossRef]
- Denis, C.; Deiteren, K.; Mortier, A.; Tounsi, A.; Fransen, E.; Proost, P.; Renauld, J.C.; Lambeir, A.M. C-terminal clipping of chemokine CCL1/I-309 enhances CCR8-mediated intracellular calcium release and anti-apoptotic activity. PLoS ONE 2012, 7, e34199. [Google Scholar] [CrossRef]
- Hoelzinger, D.B.; Smith, S.E.; Mirza, N.; Dominguez, A.L.; Manrique, S.Z.; Lustgarten, J. Blockade of CCL1 inhibits T regulatory cell suppressive function enhancing tumor immunity without affecting T effector responses. J. Immunol. 2010, 184, 6833–6842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.K.; Jan, R.L.; Lin, C.H.; Kuo, C.H.; Yang, S.N.; Chen, H.N.; Huang, M.Y.; Hung, C.H. Suppressive effects of imidapril on Th1- and Th2-related chemokines in monocytes. J. Investig. Med. 2011, 59, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.T.; Lu, J.H.; Wang, W.T.; Hung, C.H.; Sheu, C.C.; Huang, S.K. Epigenetic synergism between interleukin-4 and aryl-hydrocarbon receptor in human macrophages. J. Mol. Med. 2017, 95, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Glucocorticosteroids: Current and future directions. Br. J. Pharm. 2011, 163, 29–43. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, M.-K.; Tsai, M.-L.; Wen, Z.-H.; Liao, W.-T.; Lin, Y.-C.; Chiou, H.-Y.C.; Lin, M.-H.; Hung, C.-H. Suppressive Effects of 4-(Phenylsulfanyl) Butan-2-One on CCL-1 Production via Histone Acetylation in Monocytes. Curr. Issues Mol. Biol. 2022, 44, 4616-4625. https://doi.org/10.3390/cimb44100315
Tsai M-K, Tsai M-L, Wen Z-H, Liao W-T, Lin Y-C, Chiou H-YC, Lin M-H, Hung C-H. Suppressive Effects of 4-(Phenylsulfanyl) Butan-2-One on CCL-1 Production via Histone Acetylation in Monocytes. Current Issues in Molecular Biology. 2022; 44(10):4616-4625. https://doi.org/10.3390/cimb44100315
Chicago/Turabian StyleTsai, Ming-Kai, Mei-Lan Tsai, Zhi-Hong Wen, Wei-Ting Liao, Yi-Ching Lin, Hsin-Ying Clair Chiou, Ming-Hong Lin, and Chih-Hsing Hung. 2022. "Suppressive Effects of 4-(Phenylsulfanyl) Butan-2-One on CCL-1 Production via Histone Acetylation in Monocytes" Current Issues in Molecular Biology 44, no. 10: 4616-4625. https://doi.org/10.3390/cimb44100315