

Anti-Inflammatory Activity of Bilberry (Vaccinium myrtillus L.)

Abstract
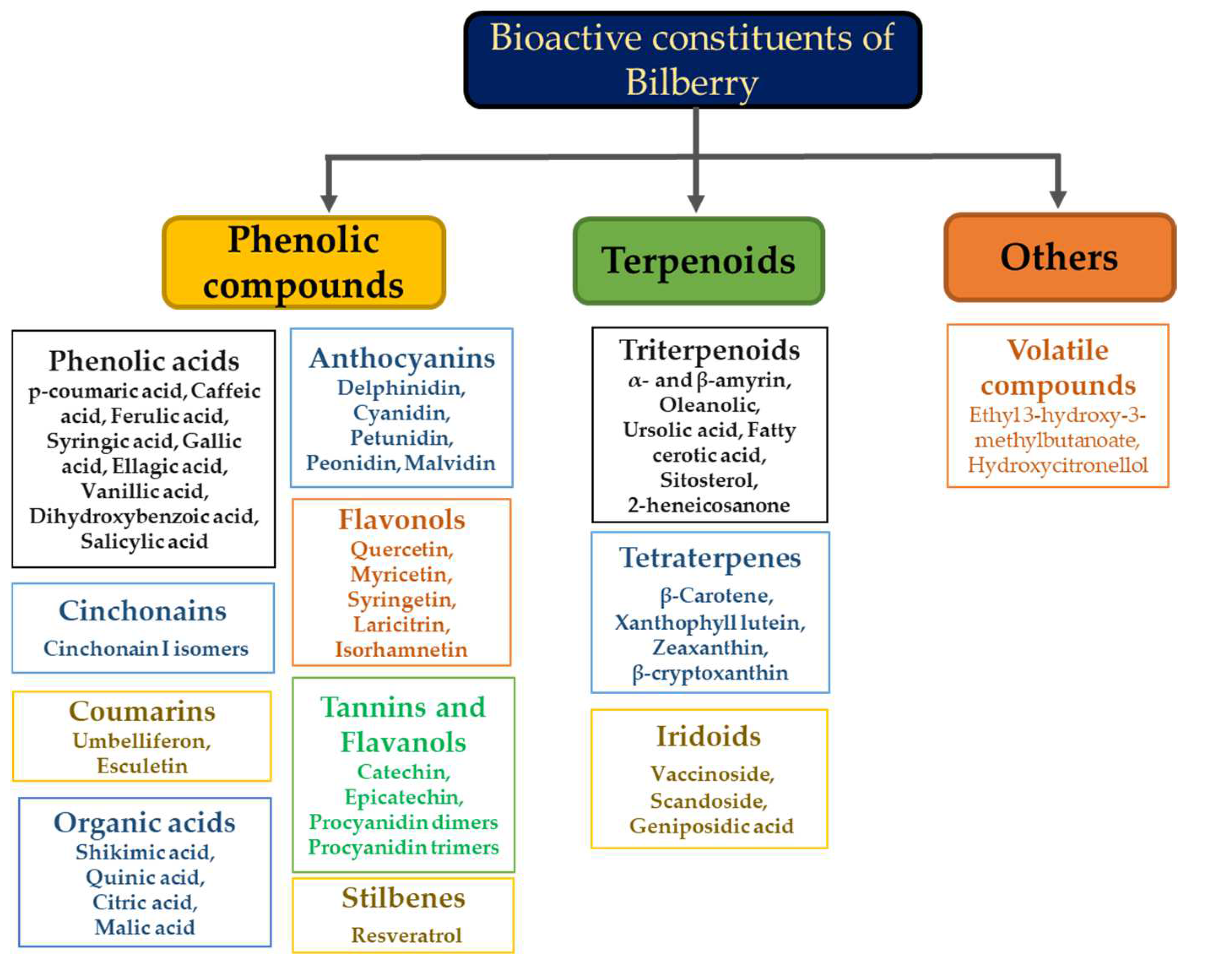
:1. Introduction
2. Bilberry
3. Search Strategy
4. Anti-Inflammatory Activity
4.1. In Vitro and In Vivo Studies

{kind=link}
| Ref. | Treatment | Model | Effects |
|---|---|---|---|
| [46] | Pretreatment with bilberry (40% anthocyanins), 75 µg/mL, 30 min + 40 ng/mL LPS for 6 h | RAW264.7 | ↓TNF-α, ↓IL-1β, ↓IL-6, ↓TNC, ↓PTGS2, ↓COX-2, ↓CCL22, ↓IFI47, and ↓IFI1 |
| [51] | H2O2 (200 μM) + bilberry extract (1, 10, 100 and 200 μg/mL), 24 h | RAW264.7 | ↓COX2, ↓IL-1β, ↑NFE2L2, ↓NF-κB1, ↓NF-κB2, ↓TNFα, inhibited NOS2 |
| [52] | LPS (1 μg/mL) with hydroethanolic extracts (400, 800 μg/mL), 24 h | RAW 264.7 | No cytotoxicity, ↓NO, ↓COX-2, ↓iNOS, ↓IL-6 and ↓TNF-α. IC50 (μg/mL) = DPPH (151.98 ± 1.41), ABTS (57.15 ± 0.95) |
| [54] | Bilberry extract 20 min prior to TNF-α or IFN-γ (100 ng/mL each) | THP-1 | ↓p-STAT1, ↓p-STAT3, ↓MCP-1, ↓IL-6, ↓TNF-α, ↓ICAM-1, ↓T-bet (transcription factor) |
| [56] | Carrageenan (450 μg/paw) 50 and 200 mg/kg, 10 days | Mice | Paw edema reduction at low dose = 35.4% and high dose = 28.8% |
| [57] | Extract concentration 0.01% to 3%, 2 h followed by LPS (25 ng/mL), 8 h HeLa-TLR4, THP-1 (50 ng/mL, 4 h), HEKTLR2/HEK-TLR4 (100 ng/mL overnight incubation) | THP-1, HeLa-TLR4, HEKTLR2/HEK-TLR4 | ↑ cell viability, anti-inflammatory effects |
| [58] | LPS (0.5 mg/kg)-induced liver damage. 50, 100 and 200 mg/kg/day, 7 days | Kunming mice | ↓plasma ALT and AST, ↓histopathogical injury, ↓TNF-α, ↓IL-1β, ↓IL-6, ↓iNOS, ↓NO, ↓NF-κB1, ↓ MDA in the liver |
| [60] | Pretreatment of bilberry−50, 100, 200 mg/kg/day, 5 days + LPS (100 mg/mouse), 24 h | BALB/C mice | ↑ORAC, ↑GSH, ↑vitamin C, ↑SOD ↑GPx, ↓MDA |
| [61] | Acute colitis (2.5% DSS, 7 days), chronic colitis (2.5% DSS, 4 cycles for 7 days interrupted by 7 days). Feed + 20% dried bilberries (10% AC extract) | BALB/C mice | Preserved normal colon length ↓IFN-γ, ↓TNF-α, ↓IL−6 |
| [62] | IFN-γ (10 ng/mL), IL-1β (5 ng/mL), TNF-α (10 ng/mL), 4 or 16 h. Anthocyanins (25−300 μM, 4 h) or BE (10−500 μg/mL, 24 h) | T84 | ↓TNF-α, ↓IP-10, ↓I-TAC, ↓sICAM-1, ↓GRO-α |
| [64] | HFD + 5% or 10% (w/w) of whole bilberries (BB), 24 weeks | C57BL/6J mice | ↓Weight gain, ↓Th1 cells |
| [65] | HF diet. Bilberry anthocyanins dose: 0.1% w/w (35% ACY), 24 weeks | C57BL/6J mice | ↓TNF-α |
| [66] | Anthocyanin rich grape and bilberry juice, 1551 mg ACY/L (15 mg ACY per day), 10 weeks | Fischer rats | ↓serum cholesterol, ↓TG, ↓leptin, ↓resistin No changes in adiponectin, secretion of adipokines from adipose tissue |
4.2. Clinical Studies
| Ref. | Design and Duration | Number | Subjects Type | Age (Y) | Dose/Day and Format | Biomarkers |
|---|---|---|---|---|---|---|
| [67] | Parallel, 2 –arm, controlled, 4 weeks | 62 | Males and females. CVD at risk | 30–70 | 300 mL juice per day | ↓IL-6, ↓IL-15, ↓ hsCRP, ↓MIG, ↑ TNF-α (unexpected) |
| [68] | Crossover, 2 phase, baseline controlled, 33 to 35 days | 110 | Females, overweight and obese | 44.2 | ~100 g of frozen and whole | ↓ TNF-α, ↓ sVCAM-1, ↓ adiponectin |
| [71] | Parallel, 2 –arm, controlled. 4 weeks run-in period followed by 4-week recovery period | 27 | Males and females with MetS features | 53 | 60 g of freeze-dried powder, water slurry | ↓ hsCRP, ↓ IL-12, ↓ IL-6, ↓ circulating LPS |
| [72] | Randomized controlled trial, 28 days | 55 | Normal healthy and MetS | 25–75 | MEDOX® food supplement capsules, 320 mg per day | ↓FBG, ↓TG, ↓LDL-C, ↓hsCRP, ↓ ADP-induced platelet |
| [73] | Clinical trial, 4 weeks | 35 | Normal and MetS | 25–75 | MEDOX® (purified anthocyanins) 320 mg per day | ↓ hsCRP, ↓TNF-α, ↓IL-6, ↓IL-1 α, ↓ COX-2, ↓ FBG, ↓TC, ↓TG, ↓LDL-C, ↑ PPAR-γ |
| [74] | Single blind, randomized, placebo-controlled, parallel study, 8 days | 21 | Males and females, recreational runners | -- | 200 mL juice twice per day | ↑ moderate CRP 24 h post-race (unexpected) |
| [77] | Randomized placebo controlled, double-blinded trial, 24 weeks | 150 | Hypercholesterolemia subjects | 40–65 | Anthocyanins (320 mg/day) purified from bilberry and blackcurrant | ↓ hsCRP, ↓ sVCAM-1, ↓ IL-1β, ↓ LDL-C, ↑ HDL-C |
| [78] | Randomized controlled trial, 3 weeks | 120 | Aged healthy volunteer men and women | 40–74 | 2 Medox capsules of 75 mg 2 times per day | ↓IL-8, ↓ RANTES, ↓IFN-α, ↓ IL-4, ↓IL-13 |
| [79] | Open pilot trial, 6 weeks | 13 | Patients with mild to moderate UC | 18–65 | 160 g of bilberry preparation | ↓Fecal calprotectin levels, ↓endoscopic Mayo score |
| [80] | Open pilot trial *, 6 weeks | 13 | Patients with mild to moderate UC | 18–65 | 160 g of bilberry preparation | ↓IFN-γ, ↓TNF-α, ↓p-p65-NF-κB, ↑IL-22, ↑IL-10 |
| [81] | Double blind, placebo-controlled intervention study, 9 weeks | 60 | Aged men with SMI | ≥ 67 | 330 mL of bilberry/red grape preparation, 2 times a day | ↓IL6, ↓TNF-α, ↓EGF, ↓Mip1β, ↓VEGF |
5. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The crucial roles of inflammatory mediators in inflammation: A review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D.; Alon, R.; von Andrian, U.H. Immune cell migration in inflammation: Present and future therapeutic targets. Nat. Immunol. 2005, 6, 1182–1190. [Google Scholar] [CrossRef]
- Joseph, S.V.; Edirisinghe, I.; Burton-Freeman, B.M. Berries: Anti-inflammatory effects in humans. J. Agric. Food Chem. 2014, 62, 3886–3903. [Google Scholar] [CrossRef]
- Roh, J.S.; Sohn, D.H. Damage-associated molecular patterns in inflammatory diseases. Immune. Netw. 2018, 18, e27. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, X.; Zhang, R.; Koci, M.; Si, D.; Ahmad, B.; Guo, H.; Hou, Y. C-terminal amination of a cationic anti-Inflammatory Peptide Improves Bioavailability and Inhibitory Activity Against LPS-Induced Inflammation. Front. Immunol. 2021, 11, 618312. [Google Scholar] [CrossRef]
- Page, M.J.; Kell, D.B.; Pretorius, E. The Role of Lipopolysaccharide-Induced Cell Signalling in Chronic Inflammation. Chronic Stress 2022, 6, 6390. [Google Scholar] [CrossRef]
- Santa-Cecília, F.V.; Socias, B.; Ouidja, M.O.; Sepulveda-Diaz, J.E.; Acuna, L.; Silva, R.L.; Michel, P.P.; Del-Bel, E.; Cunha, T.M.; Raisman-Vozari, R. Doxycycline suppresses microglial activation by inhibiting the p38 MAPK and NF-kB signaling pathways. Neurotox. Res. 2016, 29, 447–459. [Google Scholar] [CrossRef]
- Sun, S.C.; Ley, S.C. New insights into NF-kappaB regulation and function. Trends Immunol. 2008, 29, 469–478. [Google Scholar] [CrossRef]
- Amaro-Leal, Â.; Shvachiy, L.; Pinto, R.; Geraldes, V.; Rocha, I.; Mota-Filipe, H. Therapeutic effects of IkB kinase inhibitor during systemic inflammation. Int. Immunopharmacol. 2020, 84, 106509. [Google Scholar] [CrossRef]
- Vallabhapurapu, S.; Karin, M. Regulation and function of NF-kappaB transcription factors in the immune system. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef] [PubMed]
- Limtrakul, P.; Yodkeeree, S.; Pitchakarn, P.; Punfa, W. Anti-inflammatory effects of proanthocyanidin-rich red rice extract via suppression of MAPK, AP-1 and NF-κB pathways in Raw 264.7 macrophages. Nutr. Res. Pract. 2016, 10, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407. [Google Scholar] [CrossRef]
- Chang, H.-C.; Lin, K.-H.; Tai, Y.-T.; Chen, J.-T.; Chen, R.-M. Lipoteichoic acid-induced TNF-α and IL-6 gene expressions and oxidative stress production in macrophages are suppressed by ketamine through downregulating toll-like receptor 2-mediated activation of ERK1/2 and NFκB. Shock 2010, 33, 485–492. [Google Scholar] [CrossRef]
- Ha, A.T.; Kim, M.-Y.; Cho, J.Y. TAK1/AP-1-Targeted anti-inflammatory effects of Barringtonia augusta methanol extract. Molecules 2021, 26, 3053. [Google Scholar] [CrossRef]
- Prasain, J.K.; Grubbs, C.; Barnes, S. Cranberry anti-cancer compounds and their uptake and metabolism: An updated review. J. Berry Res. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Bilawal, A.; Ishfaq, M.; Gantumur, M.-A.; Qayum, A.; Shi, R.; Fazilani, S.A.; Anwar, A.; Jiang, Z.; Hou, J. A review of the bioactive ingredients of berries and their applications in curing diseases. Food Biosci. 2021, 44, 101407. [Google Scholar] [CrossRef]
- Mantzorou, M.; Zarros, A.; Vasios, G.; Theocharis, S.; Pavlidou, E.; Giaginis, C. Cranberry: A promising natural source of potential nutraceuticals with anticancer activity. Anti-Cancer Agents Med. Chem. 2019, 19, 1672–1686. [Google Scholar] [CrossRef]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Cianciosi, D.; Quiles, J.L.; Mezzetti, B.; Xiao, J.; Giampieri, F.; Battino, M. The efficacy of berries against lipopolysaccharide-induced inflammation: A review. Trends Food Sci. Technol. 2021, 117, 74–91. [Google Scholar] [CrossRef]
- Baby, B.; Antony, P.; Vijayan, R. Antioxidant and anticancer properties of berries. Crit. Rev. Food Sci. Nutr. 2018, 58, 2491–2507. [Google Scholar] [CrossRef]
- Bader Ul Ain, H.; Tufail, T.; Javed, M.; Tufail, T.; Arshad, M.U.; Hussain, M.; Gull Khan, S.; Bashir, S.; Al Jbawi, E.; Abdulaali Saewan, S. Phytochemical profile and pro-healthy properties of berries. Int. J. Food Prop. 2022, 25, 1714–1735. [Google Scholar] [CrossRef]
- Albers, R.; Bourdet-Sicard, R.; Braun, D.; Calder, P.C.; Herz, U.; Lambert, C.; Lenoir-Wijnkoop, I.; Meheust, A.; Ouwehand, A.; Phothirath, P. Monitoring immune modulation by nutrition in the general population: Identifying and substantiating effects on human health. Br. J. Nutr. 2013, 110, S1–S30. [Google Scholar] [CrossRef]
- Gaspar, D.P.; Lechtenberg, M.; Hensel, A. Quality assessment of bilberry fruits (Vaccinium myrtillus) and bilberry-containing dietary supplements. J. Agric. Food Chem. 2021, 69, 2213–2225. [Google Scholar] [CrossRef]
- Ulbricht, C.; Basch, E.; Basch, S.; Bent, S.; Boon, H.; Burke, D.; Costa, D.; Falkson, C.; Giese, N.; Goble, M. An evidence-based systematic review of bilberry (Vaccinium myrtillus) by the natural standard research collaboration. J. Diet. Suppl. 2009, 6, 162–200. [Google Scholar] [CrossRef]
- Zoratti, L.; Klemettilä, H.; Jaakola, L. Bilberry (Vaccinium myrtillus L.) ecotypes. In Nutritional Composition of Fruit Cultivars; Elsevier: Amsterdam, The Netherlands, 2016; pp. 83–99. [Google Scholar]
- Chu, W.-k.; Cheung, S.C.; Lau, R.A.; Benzie, I.F. Bilberry (Vaccinium myrtillus L.). In Herbal Medicine: Biomolecular and Clinical Aspects; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011. [Google Scholar]
- Smeriglio, A.; Davide, B.; Laganà, G.; Bellocco, E.; Trombetta, D. Bilberry (Vaccinium myrtyllus L.). In Nonvitamin and Nonmineral Nutritional Supplements; Elsevier: Amsterdam, The Netherlands, 2019; pp. 159–163. [Google Scholar]
- Coudun, C.; Gégout, J.C. Quantitative prediction of the distribution and abundance of Vaccinium myrtillus with climatic and edaphic factors. J. Veg. Sci. 2007, 18, 517–524. [Google Scholar] [CrossRef]
- Elisabetta, B.; Flavia, G.; Paolo, F.; Giorgio, L.; Attilio, S.G.; Fiorella, L.S.; Juri, N. Nutritional profile and productivity of bilberry (Vaccinium myrtillus L.) in different habitats of a protected area of the eastern Italian Alps. J. Food Sci. 2013, 78, C673–C678. [Google Scholar] [CrossRef]
- Vaneková, Z.; Rollinger, J.M. Bilberries: Curative and Miraculous–A Review on Bioactive Constituents and Clinical Research. Front. Pharmacol. 2022, 13, 909914. [Google Scholar] [CrossRef]
- Åkerström, A.; Jaakola, L.; Bång, U.; Jaderlund, A. Effects of latitude-related factors and geographical origin on anthocyanidin concentrations in fruits of Vaccinium myrtillus L.(bilberries). J. Agric. Food Chem. 2010, 58, 11939–11945. [Google Scholar] [CrossRef]
- Yamaura, K.; Ishiwatari, M.; Yamamoto, M.; Shimada, M.; Bi, Y.; Ueno, K. Anthocyanins, but not anthocyanidins, from bilberry (Vaccinium myrtillus L.) alleviate pruritus via inhibition of mast cell degranulation. J. Food Sci. 2012, 77, H262–H267. [Google Scholar] [CrossRef]
- Thibado, S.P.; Thornthwaite, J.T.; Ballard, T.K.; Goodman, B.T. Anticancer effects of Bilberry anthocyanins compared with NutraNanoSphere encapsulated Bilberry anthocyanins. Mol. Clin. Oncol. 2018, 8, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Bujor, O.-C.; Le Bourvellec, C.; Volf, I.; Popa, V.I.; Dufour, C. Seasonal variations of the phenolic constituents in bilberry (Vaccinium myrtillus L.) leaves, stems and fruits, and their antioxidant activity. Food Chem. 2016, 213, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Satoh, Y.; Ishihara, K. Investigation of the antimicrobial activity of bilberry (Vaccinium myrtillus L.) extract against periodontopathic bacteria. J. Oral Biosci. 2020, 62, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Ashour, O.M.; Elberry, A.A.; Alahdal, A.M.; Al Mohamadi, A.M.; Nagy, A.A.; Abdel-Naim, A.B.; Abdel-Sattar, E.A.; Mohamadin, A.M. Protective effect of bilberry (Vaccinium myrtillus) against doxorubicin-induced oxidative cardiotoxicity in rats. Med. Sci. Monit. 2011, 17, BR110. [Google Scholar] [CrossRef] [PubMed]
- Osada, H.; Okamoto, T.; Kawashima, H.; Toda, E.; Miyake, S.; Nagai, N.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Neuroprotective effect of bilberry extract in a murine model of photo-stressed retina. PLoS ONE 2017, 12, e0178627. [Google Scholar] [CrossRef]
- Talavéra, S.; Felgines, C.; Texier, O.; Besson, C.; Mazur, A.; Lamaison, J.L.; Rémésy, C. Bioavailability of a bilberry anthocyanin extract and its impact on plasma antioxidant capacity in rats. J. Sci. Food Agric. 2006, 86, 90–97. [Google Scholar] [CrossRef]
- Sakakibara, H.; Ogawa, T.; Koyanagi, A.; Kobayashi, S.; Goda, T.; Kumazawa, S.; Kobayashi, H.; Shimoi, K. Distribution and excretion of bilberry anthocyanins in mice. J. Agric. Food Chem. 2009, 57, 7681–7686. [Google Scholar] [CrossRef]
- Mueller, D.; Jung, K.; Winter, M.; Rogoll, D.; Melcher, R.; Richling, E. Human intervention study to investigate the intestinal accessibility and bioavailability of anthocyanins from bilberries. Food Chem. 2017, 231, 275–286. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, N. Bioavailability of anthocyanins and derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Nurmi, T.; Mursu, J.; Heinonen, M.; Nurmi, A.; Hiltunen, R.; Voutilainen, S. Metabolism of berry anthocyanins to phenolic acids in humans. J. Agric. Food Chem. 2009, 57, 2274–2281. [Google Scholar] [CrossRef]
- Mueller, D.; Jung, K.; Winter, M.; Rogoll, D.; Melcher, R.; Kulozik, U.; Schwarz, K.; Richling, E. Encapsulation of anthocyanins from bilberries–Effects on bioavailability and intestinal accessibility in humans. Food Chem. 2018, 248, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Du, B.; Xu, B. Anti-inflammatory effects of phytochemicals from fruits, vegetables, and food legumes: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Uto, T.; Tanigawa, S.; Kumamoto, T.; Fujii, M.; Hou, D.-X. Expression profiling of genes targeted by bilberry (Vaccinium myrtillus) in macrophages through DNA microarray. Nutr. Cancer 2008, 60, 43–50. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, E.H.; Hahm, K.B. Oxidative stress in inflammation-based gastrointestinal tract diseases: Challenges and opportunities. J. Gastroenterol. Hepatol. 2012, 27, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.; Lee, M.-G. Oxidative stress and antioxidant strategies in dermatology. Redox Rep. 2016, 21, 164–169. [Google Scholar] [CrossRef]
- Kang, K.S. Phytochemical constituents of medicinal plants for the treatment of chronic inflammation. Biomolecules 2021, 11, 672. [Google Scholar] [CrossRef]
- Pomari, E.; Stefanon, B.; Colitti, M. Effect of plant extracts on H2O2-induced inflammatory gene expression in macrophages. J. Inflamm. Res. 2014, 7, 103. [Google Scholar]
- Bayazid, A.B.; Chun, E.M.; Al Mijan, M.; Park, S.H.; Moon, S.-K.; Lim, B.O. Anthocyanins profiling of bilberry (Vaccinium myrtillus L.) extract that elucidates antioxidant and anti-inflammatory effects. Food Agric. Immunol. 2021, 32, 713–726. [Google Scholar]
- Wu, L.-H.; Xu, Z.-L.; Dong, D.; He, S.-A.; Yu, H. Protective effect of anthocyanins extract from blueberry on TNBS-induced IBD model of mice. Evid. Based Complement. Altern. Med. 2011, 2011, 525462. [Google Scholar] [CrossRef]
- Roth, S.; Spalinger, M.R.; Müller, I.; Lang, S.; Rogler, G.; Scharl, M. Bilberry-derived anthocyanins prevent IFN-γ-induced pro-inflammatory signalling and cytokine secretion in human THP-1 monocytic cells. Digestion 2014, 90, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Horvath, C.M. The Jak-STAT pathway stimulated by interferon alpha or interferon beta. Sci STKE 2004, 2004, tr10. [Google Scholar] [PubMed]
- Nardi, G.M.; Januario, A.G.F.; Freire, C.G.; Megiolaro, F.; Schneider, K.; Perazzoli, M.R.A.; Do Nascimento, S.R.; Gon, A.C.; Mariano, L.N.B.; Wagner, G. Anti-inflammatory activity of berry fruits in mice model of inflammation is based on oxidative stress modulation. Pharmacogn. Res. 2016, 8, S42. [Google Scholar]
- Schink, A.; Neumann, J.; Leifke, A.L.; Ziegler, K.; Fröhlich-Nowoisky, J.; Cremer, C.; Thines, E.; Weber, B.; Pöschl, U.; Schuppan, D. Screening of herbal extracts for TLR2-and TLR4-dependent anti-inflammatory effects. PLoS ONE 2018, 13, e0203907. [Google Scholar] [CrossRef]
- Luo, H.; Lv, X.-D.; Wang, G.-E.; Li, Y.-F.; Kurihara, H.; He, R.-R. Anti-inflammatory effects of anthocyanins-rich extract from bilberry (Vaccinium myrtillus L.) on croton oil-induced ear edema and Propionibacterium acnes plus LPS-induced liver damage in mice. Int. J. Food Sci. Nutr. 2014, 65, 594–601. [Google Scholar] [CrossRef]
- Maleki, A.; Anesi, S.D.; Look-Why, S.; Manhapra, A.; Foster, C.S. Pediatric uveitis: A comprehensive review. Surv. Ophthalmol. 2022, 67, 510–529. [Google Scholar] [CrossRef]
- Yao, N.; Lan, F.; He, R.-R.; Kurihara, H. Protective effects of bilberry (Vaccinium myrtillus L.) extract against endotoxin-induced uveitis in mice. J. Agric. Food Chem. 2010, 58, 4731–4736. [Google Scholar] [CrossRef]
- Piberger, H.; Oehme, A.; Hofmann, C.; Dreiseitel, A.; Sand, P.G.; Obermeier, F.; Schoelmerich, J.; Schreier, P.; Krammer, G.; Rogler, G. Bilberries and their anthocyanins ameliorate experimental colitis. Mol. Nutr. Food Res. 2011, 55, 1724–1729. [Google Scholar] [CrossRef]
- Triebel, S.; Trieu, H.-L.; Richling, E. Modulation of inflammatory gene expression by a bilberry (Vaccinium myrtillus L.) extract and single anthocyanins considering their limited stability under cell culture conditions. J. Agric. Food Chem. 2012, 60, 8902–8910. [Google Scholar] [CrossRef]
- Duan, Y.; Zeng, L.; Zheng, C.; Song, B.; Li, F.; Kong, X.; Xu, K. Inflammatory links between high fat diets and diseases. Front. Immunol. 2018, 9, 2649. [Google Scholar] [CrossRef]
- Mykkänen, O.T.; Huotari, A.; Herzig, K.-H.; Dunlop, T.W.; Mykkänen, H.; Kirjavainen, P.V. Wild blueberries (Vaccinium myrtillus) alleviate inflammation and hypertension associated with developing obesity in mice fed with a high-fat diet. PLoS ONE 2014, 9, e114790. [Google Scholar] [CrossRef] [PubMed]
- van der Heijden, R.A.; Morrison, M.C.; Sheedfar, F.; Mulder, P.; Schreurs, M.; Hommelberg, P.P.; Hofker, M.H.; Schalkwijk, C.; Kleemann, R.; Tietge, U.J. Effects of anthocyanin and flavanol compounds on lipid metabolism and adipose tissue associated systemic inflammation in diet-induced obesity. Mediat. Inflamm. 2016, 2016, 2042107. [Google Scholar] [CrossRef]
- Graf, D.; Seifert, S.; Jaudszus, A.; Bub, A.; Watzl, B. Anthocyanin-rich juice lowers serum cholesterol, leptin, and resistin and improves plasma fatty acid composition in fischer rats. PLoS ONE 2013, 8, e66690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsen, A.; Paur, I.; Bøhn, S.K.; Sakhi, A.K.; Borge, G.I.; Serafini, M.; Erlund, I.; Laake, P.; Tonstad, S.; Blomhoff, R. Bilberry juice modulates plasma concentration of NF-κB related inflammatory markers in subjects at increased risk of CVD. Eur. J. Nutr. 2010, 49, 345–355. [Google Scholar] [CrossRef]
- Lehtonen, H.-M.; Suomela, J.; Tahvonen, R.; Yang, B.; Venojärvi, M.; Viikari, J.; Kallio, H. Different berries and berry fractions have various but slightly positive effects on the associated variables of metabolic diseases on overweight and obese women. Eur. J. Clin. Nutr. 2011, 65, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Stumvoll, M. Adiponectin-its role in metabolism and beyond. Horm. Metab. Res. 2002, 34, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Metabolic syndrome update. Trends Cardiovasc. Med. 2016, 26, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Kolehmainen, M.; Mykkänen, O.; Kirjavainen, P.V.; Leppänen, T.; Moilanen, E.; Adriaens, M.; Laaksonen, D.E.; Hallikainen, M.; Puupponen-Pimiä, R.; Pulkkinen, L. Bilberries reduce low-grade inflammation in individuals with features of metabolic syndrome. Mol. Nutr. Food Res. 2012, 56, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Aboonabi, A.; Meyer, R.R.; Gaiz, A.; Singh, I. Anthocyanins in berries exhibited anti-atherogenicity and antiplatelet activities in a metabolic syndrome population. Nutr. Res. 2020, 76, 82–93. [Google Scholar] [CrossRef]
- Aboonabi, A.; Aboonabi, A. Anthocyanins reduce inflammation and improve glucose and lipid metabolism associated with inhibiting nuclear factor-kappaB activation and increasing PPAR-γ gene expression in metabolic syndrome subjects. Free Radic. Biol. Med. 2020, 150, 30–39. [Google Scholar] [CrossRef]
- Lynn, A.; Garner, S.; Nelson, N.; Simper, T.N.; Hall, A.C.; Ranchordas, M.K. Effect of bilberry juice on indices of muscle damage and inflammation in runners completing a half-marathon: A randomised, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2018, 15, 22. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xia, M.; Ma, J.; Hao, Y.; Liu, J.; Mou, H.; Cao, L.; Ling, W. Anthocyanin supplementation improves serum LDL-and HDL-cholesterol concentrations associated with the inhibition of cholesteryl ester transfer protein in dyslipidemic subjects. Am. J. Clin. Nutr. 2009, 90, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xia, M.; Yang, Y.; Liu, F.; Li, Z.; Hao, Y.; Mi, M.; Jin, T.; Ling, W. Purified anthocyanin supplementation improves endothelial function via NO-cGMP activation in hypercholesterolemic individuals. Clinic. Chem. 2011, 57, 1524–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Ling, W.; Guo, H.; Song, F.; Ye, Q.; Zou, T.; Li, D.; Zhang, Y.; Li, G.; Xiao, Y. Anti-inflammatory effect of purified dietary anthocyanin in adults with hypercholesterolemia: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 843–849. [Google Scholar] [CrossRef]
- Karlsen, A.; Retterstøl, L.; Laake, P.; Paur, I.; Kjølsrud-Bøhn, S.; Sandvik, L.; Blomhoff, R. Anthocyanins inhibit nuclear factor-κ B activation in monocytes and reduce plasma concentrations of pro-inflammatory mediators in healthy adults. J. Nutr. 2007, 137, 1951–1954. [Google Scholar] [CrossRef]
- Biedermann, L.; Mwinyi, J.; Scharl, M.; Frei, P.; Zeitz, J.; Kullak-Ublick, G.A.; Vavricka, S.R.; Fried, M.; Weber, A.; Humpf, H.-U. Bilberry ingestion improves disease activity in mild to moderate ulcerative colitis—An open pilot study. J. Crohns Colitis 2013, 7, 271–279. [Google Scholar] [CrossRef]
- Roth, S.; Spalinger, M.R.; Gottier, C.; Biedermann, L.; Zeitz, J.; Lang, S.; Weber, A.; Rogler, G.; Scharl, M. Bilberry-derived anthocyanins modulate cytokine expression in the intestine of patients with ulcerative colitis. PLoS ONE 2016, 11, e0154817. [Google Scholar]
- Bøhn, S.K.; Myhrstad, M.C.W.; Thoresen, M.; Erlund, I.; Vasstrand, A.K.; Marciuch, A.; Carlsen, M.H.; Bastani, N.E.; Engedal, K.; Flekkøy, K.M.; et al. Bilberry/red grape juice decreases plasma biomarkers of inflammation and tissue damage in aged men with subjective memory impairment—a randomized clinical trial. BMC Nutr. 2021, 7, 75. [Google Scholar] [CrossRef]
- Upton, R. Bilberry Fruit: Vaccinium myrtillus L. Standards of Analysis, Quality Control, and Therapeutics; American Herbal Pharmacopoeia: Santa Cruz, CA, USA, 2001. [Google Scholar]
- Pan, P.; Lam, V.; Salzman, N.; Huang, Y.-W.; Yu, J.; Zhang, J.; Wang, L.-S. Black raspberries and their anthocyanin and fiber fractions alter the composition and diversity of gut microbiota in F-344 rats. Nutr. Cancer 2017, 69, 943–951. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Lee, H.-J. Anti-Inflammatory Activity of Bilberry (Vaccinium myrtillus L.). Curr. Issues Mol. Biol. 2022, 44, 4570-4583. https://doi.org/10.3390/cimb44100313
Sharma A, Lee H-J. Anti-Inflammatory Activity of Bilberry (Vaccinium myrtillus L.). Current Issues in Molecular Biology. 2022; 44(10):4570-4583. https://doi.org/10.3390/cimb44100313
Chicago/Turabian StyleSharma, Anshul, and Hae-Jeung Lee. 2022. "Anti-Inflammatory Activity of Bilberry (Vaccinium myrtillus L.)" Current Issues in Molecular Biology 44, no. 10: 4570-4583. https://doi.org/10.3390/cimb44100313