
Mitochondrial DNA Copy Number in Cleavage Stage Human Embryos—Impact on Infertility Outcome

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Stimulation Protocol, Oocyte Retrieval
2.3. Embryo Culture, Biopsy and Transfer
2.4. Cell Lysis and Whole Genome Amplification
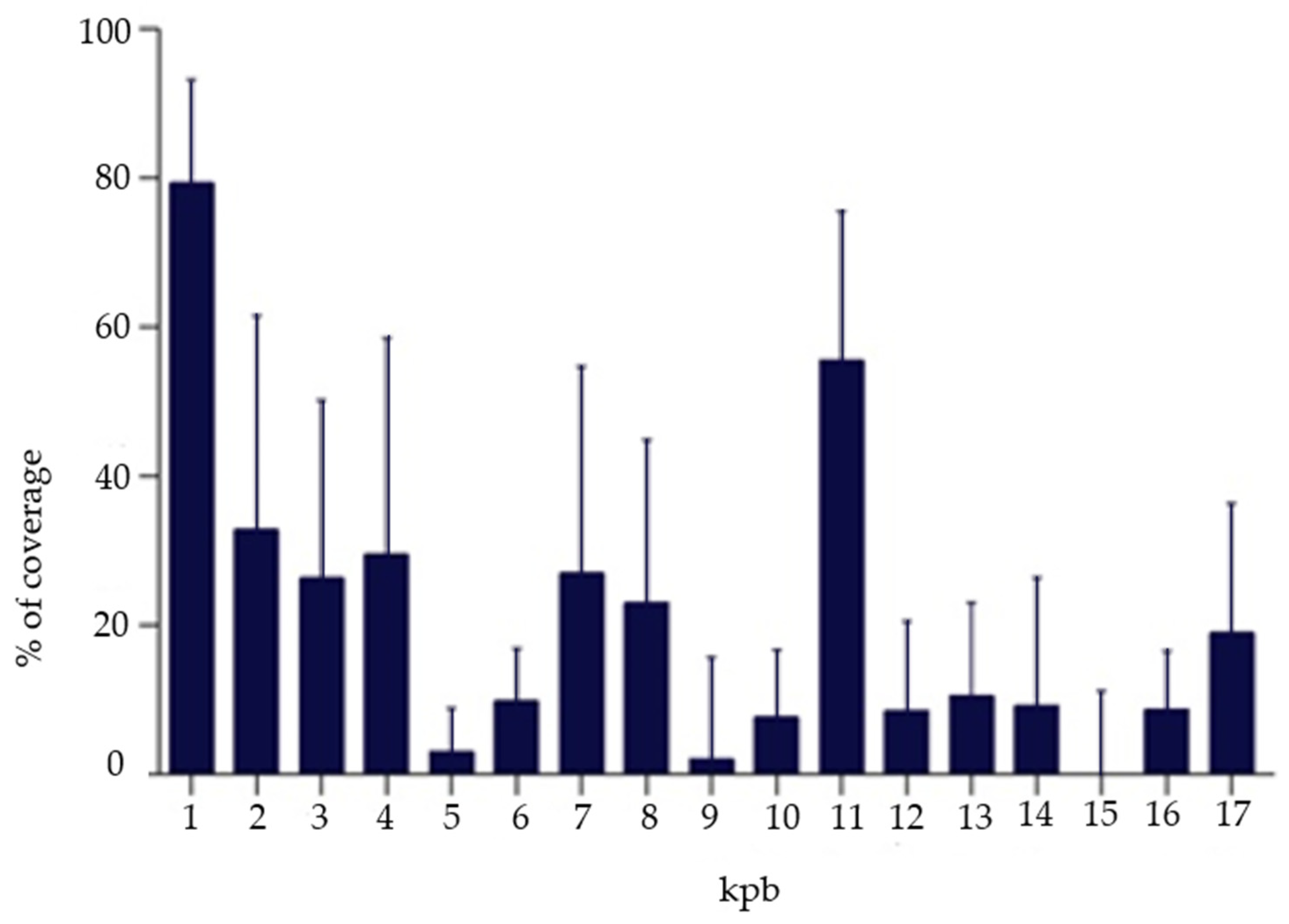
2.5. Next-Generation Sequencing (NGS)
2.6. Relative Mitochondrial DNA Quantification
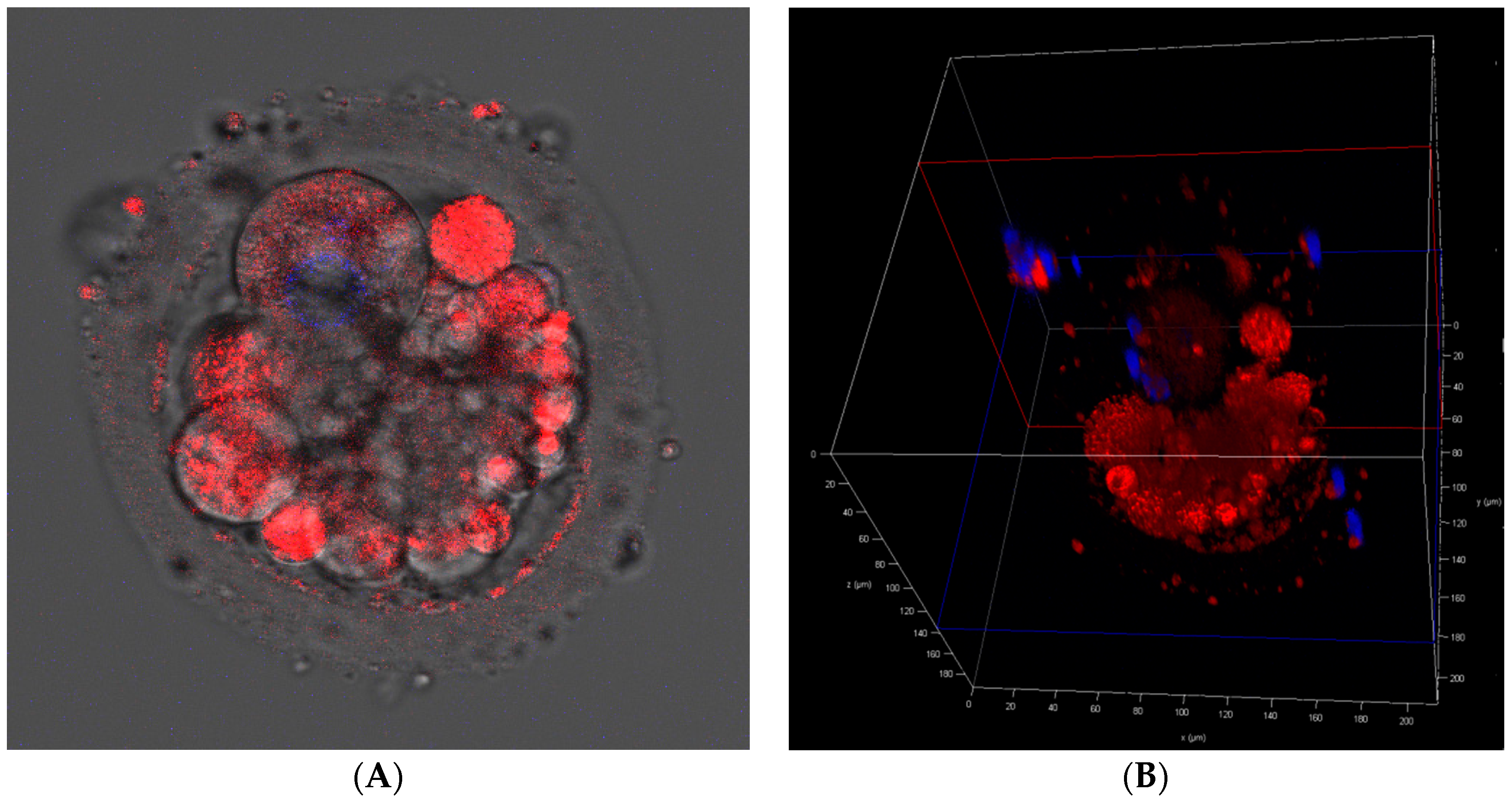
2.7. Cell Fluorescence Staining for Mitochondrial Presence/Activity
2.8. Statistical Tests
3. Results
3.1. Baseline Characteristics
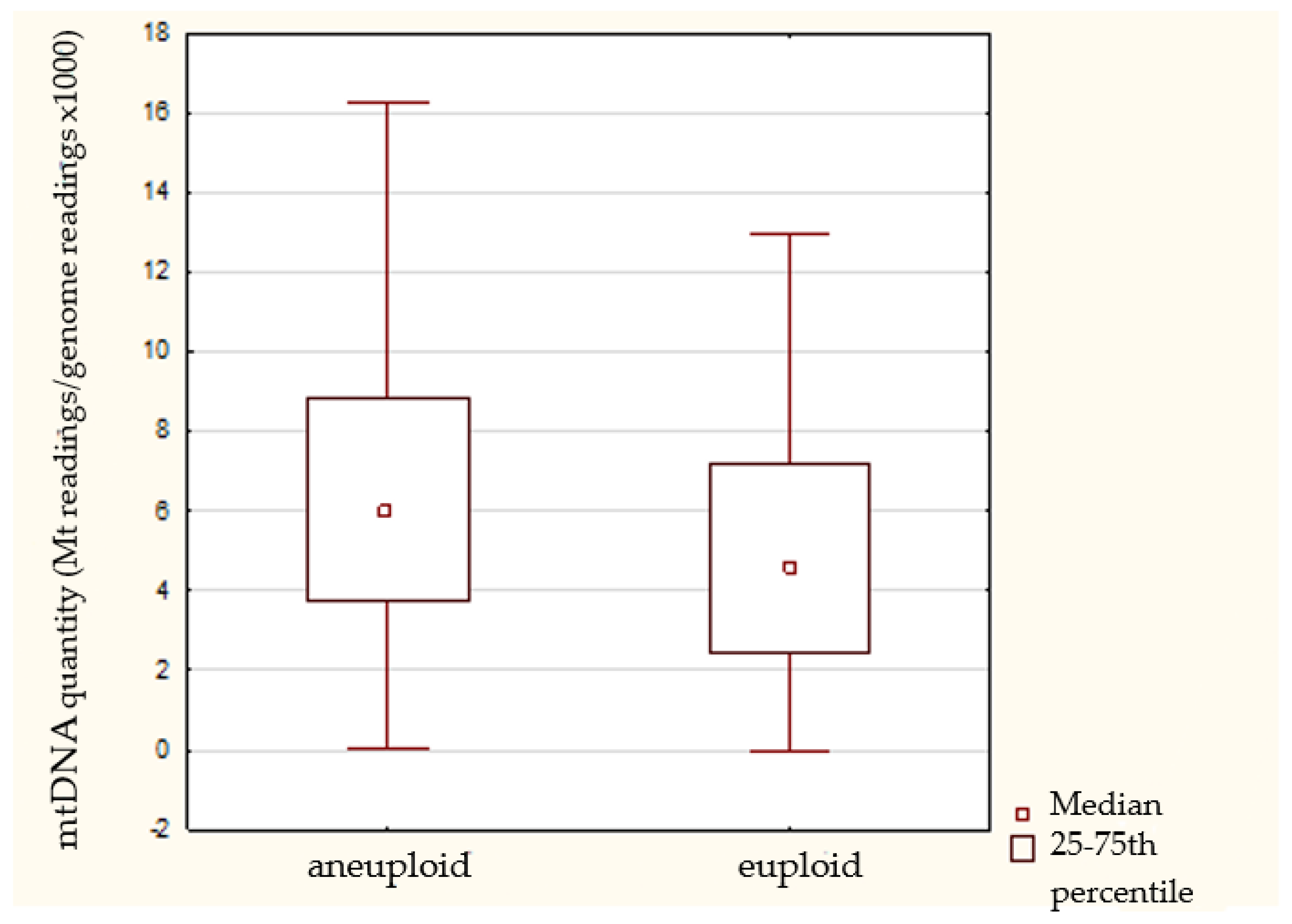
3.2. mtDNA Quantification
3.3. Ploidy Status
3.4. Embryo Quality
3.5. Embryo’s Genetic Sex
3.6. Maternal Age
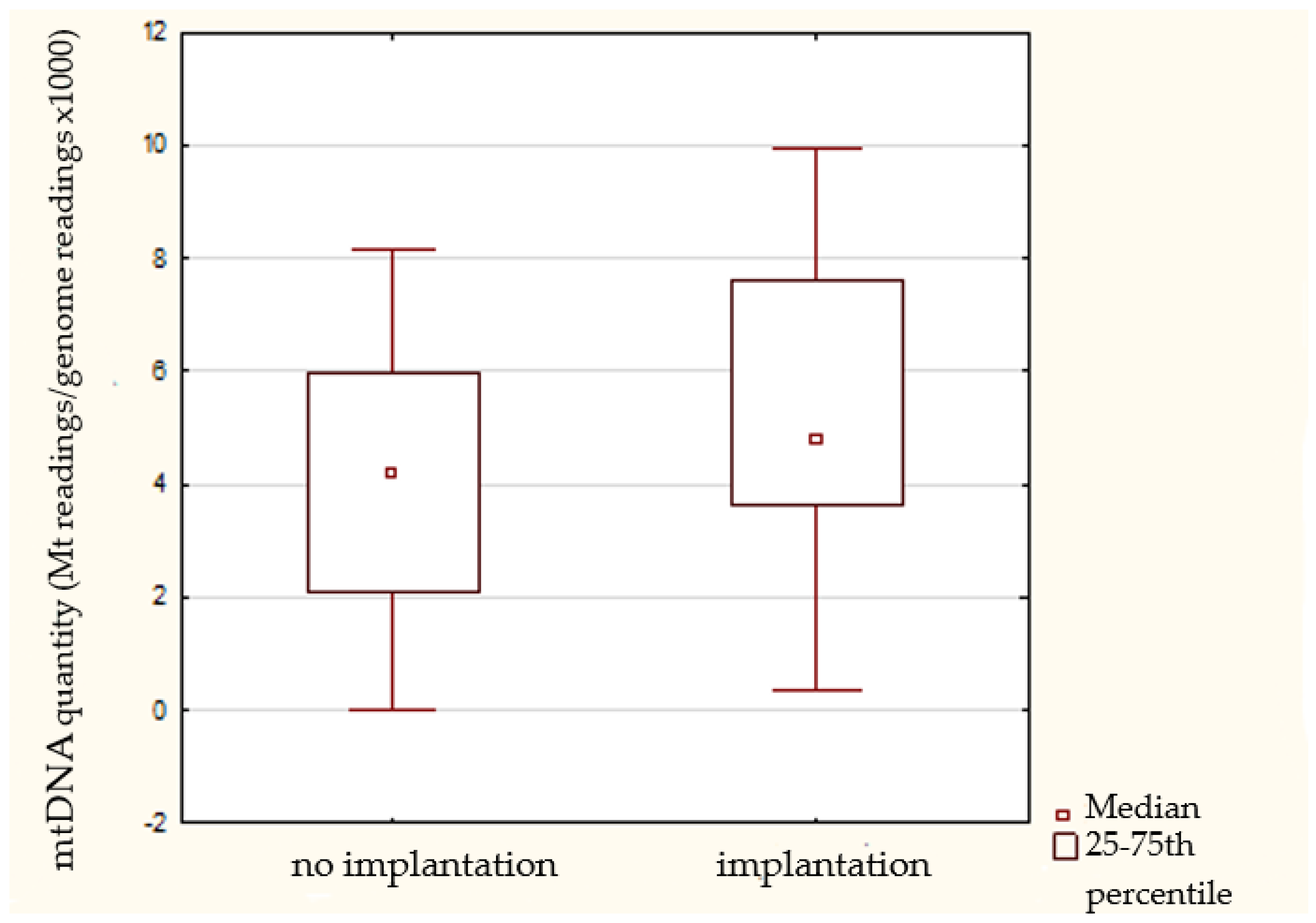
3.7. Embryo Transfer Outcomes
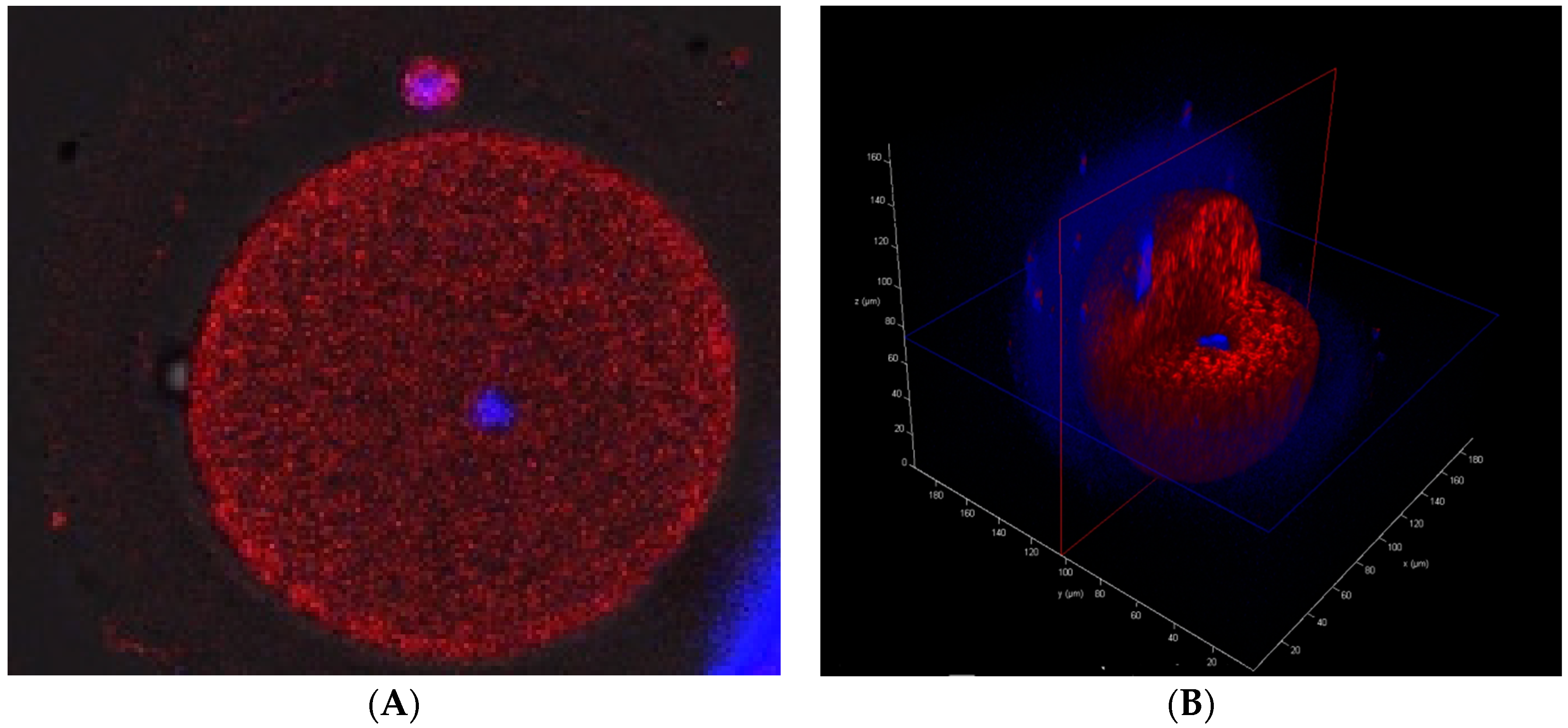
3.8. Mitochondrial Presence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- European Society of Human Reproduction and Embryology More Than 8 million Babies born from IVF Since the World’s First in 1978: European IVF Pregnancy Rates Now Steady at Around 36 Percent, According to ESHRE Monitoring. Available online: www.sciencedaily.com/releases/2018/07/180703084127.htm (accessed on 26 October 2021).
- Fragouli, E.; Alfarawati, S.; Spath, K.; Jaroudi, S.; Sarasa, J.; Enciso, M.; Wells, D. The origin and impact of embryonic aneuploidy. Hum. Genet. 2013, 132, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, M.; Marin, D.; Sisti, G. New Frontiers in IVF: mtDNA and autologous germline mitochondrial energy transfer. Reprod. Biol. Endocrinol. 2019, 17, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diez-Juan, A.; Rubio, C.; Marin, C.; Martinez, S.; Al-Asmar, N.; Riboldi, M.; Díaz-Gimeno, P.; Valbuena, D.; Simón, C. Mitochondrial DNA content as a viability score in human euploid embryos: Less is better. Fertil. Steril. 2015, 104, 534–541.e1. [Google Scholar] [CrossRef] [Green Version]
- Leese, H.J. Metabolism of the preimplantation embryo: 40 years on. Reproduction 2012, 143, 417–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Chen, S.; Chen, D.; Liu, P.; Xia, J.; Yang, L.; Song, Z.; Shi, Q.; Xie, L.; Zhu, A.; et al. The mitochondrial DNA content can not predict the embryo viability. bioRxiv 2018, 445940. [Google Scholar] [CrossRef]
- Arnanz, A.; De Munck, N.; Bayram, A.; El-Damen, A.; Abdalla, A.; ElKhatib, I.; Melado, L.; Lawrenz, B.; Fatemi, H.M. Blastocyst mitochondrial DNA (mtDNA) is not affected by oocyte vitrification: A sibling oocyte study. J. Assist. Reprod. Genet. 2020, 37, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- El-Damen, A.; Elkhatib, I.; Bayram, A.; Arnanz, A.; Abdala, A.; Samir, S.; Lawrenz, B.; De Munck, N.; Fatemi, H.M. Does blastocyst mitochondrial DNA content affect miscarriage rate in patients undergoing single euploid frozen embryo transfer? J. Assist. Reprod. Genet. 2021, 38, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.S.-Y.; Weng, S.-P.; Shen, M.-S.; Ma, P.-C.; Wu, P.-K.; Lee, N.-C. Suboptimal trophectoderm mitochondrial DNA level is associated with delayed blastocyst development. J. Assist. Reprod. Genet. 2021, 38, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Boynukalin, F.K.; Gultomruk, M.; Cavkaytar, S.; Turgut, E.; Findikli, N.; Serdarogullari, M.; Coban, O.; Yarkiner, Z.; Rubio, C.; Bahceci, M. Parameters impacting the live birth rate per transfer after frozen single euploid blastocyst transfer. PLoS ONE 2020, 15, e0227619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, R.T., 3rd; Sun, L.; Zhan, Y.; Marin, D.; Tao, X.; Seli, E. Mitochondrial DNA content is not predictive of reproductive competence in euploid blastocysts. Reprod. Biomed. Online 2020, 41, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Huang, Z.; Lin, Y.; Sun, Y.; Chen, Q.; Pan, M.; Zheng, B. Mitochondrial DNA Copy Number in Human Blastocyst: A Novel Biomarker for the Prediction of Implantation Potential. J. Mol. Diagn. 2021, 23, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Diao, Z.; Zhu, L.; Zhu, J.; Lin, F.; Jiang, W.; Fang, J.; Xu, Z.; Xing, J.; Zhou, J.; et al. Trophectoderm Mitochondrial DNA Content Associated with Embryo Quality and Day-5 Euploid Blastocyst Transfer Outcomes. DNA Cell Biol. 2021, 40, 643–651. [Google Scholar] [CrossRef]
- Zhang, Q.; Ji, H.; Shi, J.; Wang, L.; Ding, L.; Jiang, Y.; Huang, X.; Qiu, P.; Li, P. Digital PCR Detection of mtDNA/gDNA Ratio in Embryo Culture Medium for Prediction of Embryo Development Potential. Pharmgenomics Pers. Med. 2021, 14, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Kobayashi, J.; Shirasuna, K.; Iwata, H. Abundance of cell-free mitochondrial DNA in spent culture medium associated with morphokinetics and blastocyst collapse of expanded blastocysts. Reprod. Med. Biol. 2020, 19, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ito, J.; Shirasuna, K.; Kuwayama, T.; Iwata, H. Comparative analysis of cell-free DNA content in culture medium and mitochondrial DNA copy number in porcine parthenogenetically activated embryos. J. Reprod. Dev. 2020, 66, 539–546. [Google Scholar] [CrossRef]
- Klimczak, A.M.; Pacheco, L.E.; Lewis, K.E.; Massahi, N.; Richards, J.P.; Kearns, W.G.; Saad, A.F.; Crochet, J.R. Embryonal mitochondrial DNA: Relationship to embryo quality and transfer outcomes. J. Assist. Reprod. Genet. 2018, 35, 871–877. [Google Scholar] [CrossRef] [PubMed]
- de Los Santos, M.J.; Diez Juan, A.; Mifsud, A.; Mercader, A.; Meseguer, M.; Rubio, C.; Pellicer, A. Variables associated with mitochondrial copy number in human blastocysts: What can we learn from trophectoderm biopsies? Fertil. Steril. 2018, 109, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Desquiret-Dumas, V.; Clément, A.; Seegers, V.; Boucret, L.; Ferré-L’Hotellier, V.; Bouet, P.E.; Descamps, P.; Procaccio, V.; Reynier, P.; May-Panloup, P. The mitochondrial DNA content of cumulus granulosa cells is linked to embryo quality. Hum. Reprod. 2017, 32, 607–614. [Google Scholar] [CrossRef]
- Victor, A.R.; Brake, A.J.; Tyndall, J.C.; Griffin, D.K.; Zouves, C.G.; Barnes, F.L.; Viotti, M. Accurate quantitation of mitochondrial DNA reveals uniform levels in human blastocysts irrespective of ploidy, age, or implantation potential. Fertil. Steril. 2017, 107, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Fragouli, E.; Spath, K.; Alfarawati, S.; Kaper, F.; Craig, A.; Michel, C.-E.; Kokocinski, F.; Cohen, J.; Munne, S.; Wells, D. Altered levels of mitochondrial DNA are associated with female age, aneuploidy, and provide an independent measure of embryonic implantation potential. PLoS Genet. 2015, 11, e1005241. [Google Scholar] [CrossRef] [PubMed]
- Fragouli, E.; McCaffrey, C.; Ravichandran, K.; Spath, K.; Grifo, J.A.; Munné, S.; Wells, D. Clinical implications of mitochondrial DNA quantification on pregnancy outcomes: A blinded prospective non-selection study. Hum. Reprod. 2017, 32, 2340–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravichandran, K.; McCaffrey, C.; Grifo, J.; Morales, A.; Perloe, M.; Munne, S.; Wells, D.; Fragouli, E. Mitochondrial DNA quantification as a tool for embryo viability assessment: Retrospective analysis of data from single euploid blastocyst transfers. Hum. Reprod. 2017, 32, 1282–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treff, N.R.; Zhan, Y.; Tao, X.; Olcha, M.; Han, M.; Rajchel, J.; Morrison, L.; Morin, S.J.; Scott, R.T.J. Levels of trophectoderm mitochondrial DNA do not predict the reproductive potential of sibling embryos. Hum. Reprod. 2017, 32, 954–962. [Google Scholar] [CrossRef] [Green Version]
- St John, J.C.; Facucho-Oliveira, J.; Jiang, Y.; Kelly, R.; Salah, R. Mitochondrial DNA transmission, replication and inheritance: A journey from the gamete through the embryo and into offspring and embryonic stem cells. Hum. Reprod. Update 2010, 16, 488–509. [Google Scholar] [CrossRef] [Green Version]
- Lenaz, G.; Genova, M.L. Supramolecular Organisation of the Mitochondrial Respiratory Chain: A New Challenge for the Mechanism and Control of Oxidative Phosphorylation. Mitochondrial Oxidative Phosphorylation 2012, 748, 107–144. [Google Scholar]
- Satoh, M.; Kuroiwa, T. Organization of multiple nucleoids and DNA molecules in mitochondria of a human cell. Exp. Cell Res. 1991, 196, 137–140. [Google Scholar] [CrossRef]
- Legros, F.; Malka, F.; Frachon, P.; Lombès, A.; Rojo, M. Organization and dynamics of human mitochondrial DNA. J. Cell Sci. 2004, 117, 2653–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otten, A.B.C.; Smeets, H.J.M. Evolutionary defined role of the mitochondrial DNA in fertility, disease and ageing. Hum. Reprod. Update 2015, 21, 671–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Bener, M.B.; Jiang, Z.; Wang, T.; Esencan, E.; Scott III, R.; Horvath, T.; Seli, E. Mitofusin 1 is required for female fertility and to maintain ovarian follicular reserve. Cell Death Dis. 2019, 10, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Bener, M.B.; Jiang, Z.; Wang, T.; Esencan, E.; Scott, R.; Horvath, T.; Seli, E. Mitofusin 2 plays a role in oocyte and follicle development, and is required to maintain ovarian follicular reserve during reproductive aging. Aging 2019, 11, 3919–3938. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, Q.; Li, H.; Xiang, W.; Zhang, L. Cell-free mitochondrial DNA increases granulosa cell apoptosis and reduces aged oocyte blastocyst development in the mouse. Reprod. Toxicol. 2020, 98, 278–285. [Google Scholar] [CrossRef]
- Van Blerkom, J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef]
- Barritt, J.A.; Kokot, M.; Cohen, J.; Steuerwald, N.; Brenner, C.A. Quantification of human ooplasmic mitochondria. Reprod. Biomed. Online 2002, 4, 243–247. [Google Scholar] [CrossRef]
- Santos, T.A.; El Shourbagy, S.; St John, J.C. Mitochondrial content reflects oocyte variability and fertilization outcome. Fertil. Steril. 2006, 85, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Sánchez, M.; Díez-Juan, A.; Beltrán, D.; Mifsud, A.; Mercader, A.; Vidal, C.; Labarta, E.; Pellicer, A.; Seli, E.; De Los Santos, M.J. Mitochondrial DNA content decreases during in vitro human embryo development: Insights into mitochondrial DNA variation in preimplantation embryos donated for research. Proy. De Investig. 2020, 1, 36–45. [Google Scholar] [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Di Nardo, M.; Adiga, S.K.; Talevi, R. Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells. Antioxidants 2021, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Weiss, Z.R.; Gaur, P.; Forman, E.; Williams, Z. Rapid preimplantation genetic screening using a handheld, nanopore-based DNA sequencer. Fertil. Steril. 2018, 110, 910–916.e2. [Google Scholar] [CrossRef] [Green Version]
- Łukaszuk, K.; Pukszta, S.; Wells, D.; Cybulska, C.; Liss, J.; Płóciennik, Ł.; Kuczyński, W.; Zabielska, J. Routine use of next-generation sequencing for preimplantation genetic diagnosis of blastomeres obtained from embryos on day 3 in fresh in vitro fertilization cycles. Fertil. Steril. 2015, 103, 1031–1036. [Google Scholar] [CrossRef]
- Lukaszuk, K.; Liss, J.; Kunicki, M.; Jakiel, G.; Wasniewski, T.; Woclawek-Potocka, I.; Pastuszek, E. Anti-Müllerian hormone (AMH) is a strong predictor of live birth in women undergoing assisted reproductive technology. Reprod. Biol. 2014, 14, 176–181. [Google Scholar] [CrossRef]
- Wells, D.; Kaur, K.; Grifo, J.; Glassner, M.; Taylor, J.C.; Fragouli, E.; Munne, S. Clinical utilisation of a rapid low-pass whole genome sequencing technique for the diagnosis of aneuploidy in human embryos prior to implantation. J. Med. Genet. 2014, 51, 553–562. [Google Scholar] [CrossRef] [Green Version]
- Homo Sapiens Mitochondrion, Complete Genome. Available online: https://www.ncbi.nlm.nih.gov/nuccore/251831106 (accessed on 10 September 2021).
- Chang, D.D.; Clayton, D.A. Priming of human mitochondrial DNA replication occurs at the light-strand promoter. Proc. Natl. Acad. Sci. USA 1985, 82, 351–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iborra, F.J.; Kimura, H.; Cook, P.R. The functional organization of mitochondrial genomes in human cells. BMC Biol. 2004, 2, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyedhassani, S.M.; Houshmand, M.; Kalantar, S.M.; Modabber, G.; Aflatoonian, A. No mitochondrial DNA deletions but more D-loop point mutations in repeated pregnancy loss. J. Assist. Reprod. Genet. 2010, 27, 641–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Blerkom, J.; Davis, P.; Alexander, S. Differential mitochondrial distribution in human pronuclear embryos leads to disproportionate inheritance between blastomeres: Relationship to microtubular organization, ATP content and competence. Hum. Reprod. 2000, 15, 2621–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakoshi, Y.; Sueoka, K.; Takahashi, K.; Sato, S.; Sakurai, T.; Tajima, H.; Yoshimura, Y. Embryo developmental capability and pregnancy outcome are related to the mitochondrial DNA copy number and ooplasmic volume. J. Assist. Reprod. Genet. 2013, 30, 1367–1375. [Google Scholar] [CrossRef]
- St John, J. The control of mtDNA replication during differentiation and development. Biochim. Biophys. Acta 2014, 1840, 1345–1354. [Google Scholar] [CrossRef]
- Penzias, A.S. Recurrent IVF failure: Other factors. Fertil. Steril. 2012, 97, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cram, D.S.; Shen, J.; Wang, X.; Zhang, J.; Song, Z.; Xu, G.; Li, N.; Fan, J.; Wang, S.; et al. Validation of copy number variation sequencing for detecting chromosome imbalances in human preimplantation embryos. Biol. Reprod. 2014, 91, 37. [Google Scholar] [CrossRef]
- Yin, X.; Tan, K.; Vajta, G.; Jiang, H.; Tan, Y.; Zhang, C.; Chen, F.; Chen, S.; Zhang, C.; Pan, X.; et al. Massively parallel sequencing for chromosomal abnormality testing in trophectoderm cells of human blastocysts. Biol. Reprod. 2013, 88, 69. [Google Scholar] [CrossRef]
- Fiorentino, F.; Biricik, A.; Bono, S.; Spizzichino, L.; Cotroneo, E.; Cottone, G.; Kokocinski, F.; Michel, C.-E. Development and validation of a next-generation sequencing-based protocol for 24-chromosome aneuploidy screening of embryos. Fertil. Steril. 2014, 101, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.; Cervero, A.; Mir, P.; Martinez-Conejero, J.A.; Pellicer, A.; Simón, C. The impact of next-generation sequencing technology on preimplantation genetic diagnosis and screening. Fertil. Steril. 2013, 99, 1054–1061.e3. [Google Scholar] [CrossRef] [PubMed]
- Bayram, A.; De Munck, N.; Elkhatib, I.; Arnanz, A.; Liñán, A.; Lawrenz, B.; Fatemi, H.M. Cleavage stage mitochondrial DNA is correlated with preimplantation human embryo development and ploidy status. J. Assist. Reprod. Genet. 2019, 36, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Wilding, M.; Di Matteo, L.; Dale, B. The maternal age effect: A hypothesis based on oxidative phosphorylation. Zygote 2005, 13, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Tarín, J.J.; Vendrell, F.J.; Ten, J.; Blanes, R.; van Blerkom, J.; Cano, A. The oxidizing agent tertiary butyl hydroperoxide induces disturbances in spindle organization, c-meiosis, and aneuploidy in mouse oocytes. Mol. Hum. Reprod. 1996, 2, 895–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lledo, B.; Ortiz, J.A.; Morales, R.; García-Hernández, E.; Ten, J.; Bernabeu, A.; Llácer, J.; Bernabeu, R. Comprehensive mitochondrial DNA analysis and IVF outcome. Hum. Reprod. Open 2018, 2018, hoy023. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, M.B.; Govindaraj, P.; Chawla, L.; Malhotra, N.; Singh, N.; Mittal, S.; Talwar, P.; Thangaraj, K.; Dada, R. Mitochondrial DNA variations in ova and blastocyst: Implications in assisted reproduction. Mitochondrion 2013, 13, 96–105. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Analyzed Group |
|---|---|
| No. of subjects | 69 |
| Mean (SD) age | 36.5 (4.9) |
| BMI (kg/m2) | 23.5 (4.3) |
| No. of subjects with indicated cause of infertility (%) | |
| Repeated implantation failure | 18 (26.1) |
| Advanced maternal age | 19 (27.5) |
| Male factor | 8 (11.6) |
| Unexplained | 22 (31.9) |
| Endometriosis | 2 (2.9) |
| Mean (SD) duration of infertility (years) | 4.3 (3.2) |
| AMH (ng/mL) | 2.42 (1.7) |
| AFC | 13.1 (9.4) |
| No. of cycles | 69 |
| No. of transfers | 41 |
| Duration of stimulation—days (SD) | 8.6 (1.3) |
| hMG dose (IU) (SD) | 2032.5 (427.5) |
| No. oocytes retrieved (SD) | 7.1 (3.3) |
| Fertilization rate (%) | 75.1 |
| No. of embryos transferred (mean per ET) | 62 (1.5) |
| No. of pregnancies | 28 |
| Pregnancy rate per cycle (%) | 40.6 |
| Pregnancy rate per ET (%) | 68.3 |
| Implantation rate (%) | 61.2 |
| Multiple pregnancy rate of pregnancies (%) | 11 (39.3) |
| Ectopic pregnancy (%) | 0 |
| OHSS (%) | 0 |
| Spontaneous abortion rate (%) | 4 (14.3) |
| Variable | Analysed Group | N (%) | Mean mtDNA/gDNA Ratio (±SD) | Median mtDNA/gDNA Ratio (Quartiles) | U Mann–Whitney p Value |
|---|---|---|---|---|---|
| Analyzed embryos | |||||
| Ploidy status | Euploid | 89 (28.3) | 6.3 ± 7.5 | 4.6 (2.8–7.2) | 0.004 |
| Aneuploid | 204 (64.9) | 7.1 ± 5.8 | 5.9 (3.7–8.9) | ||
| Embryo morphology (Cummins scale) | A-B class | 261 (89.1) | 6.6 ± 4.8 | 5.7 (3.5–8.6) | 0.09 |
| C class | 32 (10.9) | 8.5 ± 13.6 | 4.8 (2.4–6.9) | ||
| Sex | With chr. Y | 131 (44.7) | 6.6 ± 4.1 | 5.5 (3.8–8.5) | 0.16 |
| Without chr. Y | 162 (55.3) | 6.2 ± 6.8 | 5.0 (3.1–7.3) | ||
| Maternal age | <37 years | 151 (51.5) | 6.9 ± 7.8 | 5.1 (3.2–8.0) | 0.14 |
| ≥37 years | 142 (48.5) | 6.7 ± 4.5 | 5.7 (3.8–8.6) | ||
| Euploid embryos | |||||
| Embryo morphology (Cummins scale) | A-B class | 79 (88.7) | 5.6 ± 4.3 | 4.7 (3.1–7.2) | 0.32 |
| C class | 10 (11.3) | 11.5 ± 18.9 | 2.5 (1.5–12.9) | ||
| Sex | With chr. Y | 39 (43.8) | 6.1 ± 4.4 | 4.8 (3.6–7.3) | 0.22 |
| Without chr. Y | 50 (56.2) | 6.3 ± 9.1 | 4.1 (2.2–7.2) | ||
| Maternal age | <37 years | 61 (68.5) | 6.2 ± 8.4 | 4.1 (2.3–7.2) | 0.29 |
| ≥37 years | 28 (31.5) | 6.4 ± 5.2 | 4.7 (3.2–7.3) | ||
| Transferred euploid embryos | |||||
| Implantation status | Implanted | 38 (61.3) | 7.4 ± 6.6 | 5.0 (3.7–7.8) | 0.18 |
| Non-implanted | 24 (38.7) | 5.1 ± 4.6 | 4.1 (3.3–4.6) | ||
| Pregnancy outcomes | Livebirth | 34 (89.5) | 7.4 ± 6.5 | 5.0 (3.7–7.8) | 0.22 |
| Spontaneous abortion | 4 (10.5) | 7.0 ± 6.9 | 4.6 (3.9–7.6) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podolak, A.; Liss, J.; Kiewisz, J.; Pukszta, S.; Cybulska, C.; Rychlowski, M.; Lukaszuk, A.; Jakiel, G.; Lukaszuk, K. Mitochondrial DNA Copy Number in Cleavage Stage Human Embryos—Impact on Infertility Outcome. Curr. Issues Mol. Biol. 2022, 44, 273-287. https://doi.org/10.3390/cimb44010020
Podolak A, Liss J, Kiewisz J, Pukszta S, Cybulska C, Rychlowski M, Lukaszuk A, Jakiel G, Lukaszuk K. Mitochondrial DNA Copy Number in Cleavage Stage Human Embryos—Impact on Infertility Outcome. Current Issues in Molecular Biology. 2022; 44(1):273-287. https://doi.org/10.3390/cimb44010020
Chicago/Turabian StylePodolak, Amira, Joanna Liss, Jolanta Kiewisz, Sebastian Pukszta, Celina Cybulska, Michal Rychlowski, Aron Lukaszuk, Grzegorz Jakiel, and Krzysztof Lukaszuk. 2022. "Mitochondrial DNA Copy Number in Cleavage Stage Human Embryos—Impact on Infertility Outcome" Current Issues in Molecular Biology 44, no. 1: 273-287. https://doi.org/10.3390/cimb44010020