
Endothelial Effects of Simultaneous Expression of Human HO-1, E5NT, and ENTPD1 in a Mouse
 , , ,
, , ,  , and
, and
Abstract
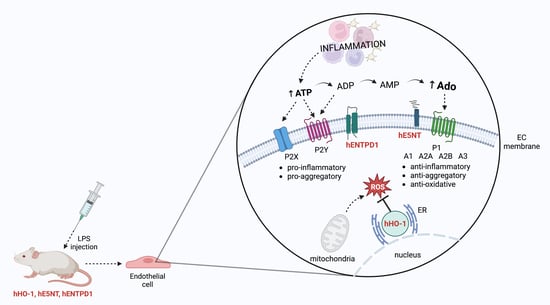
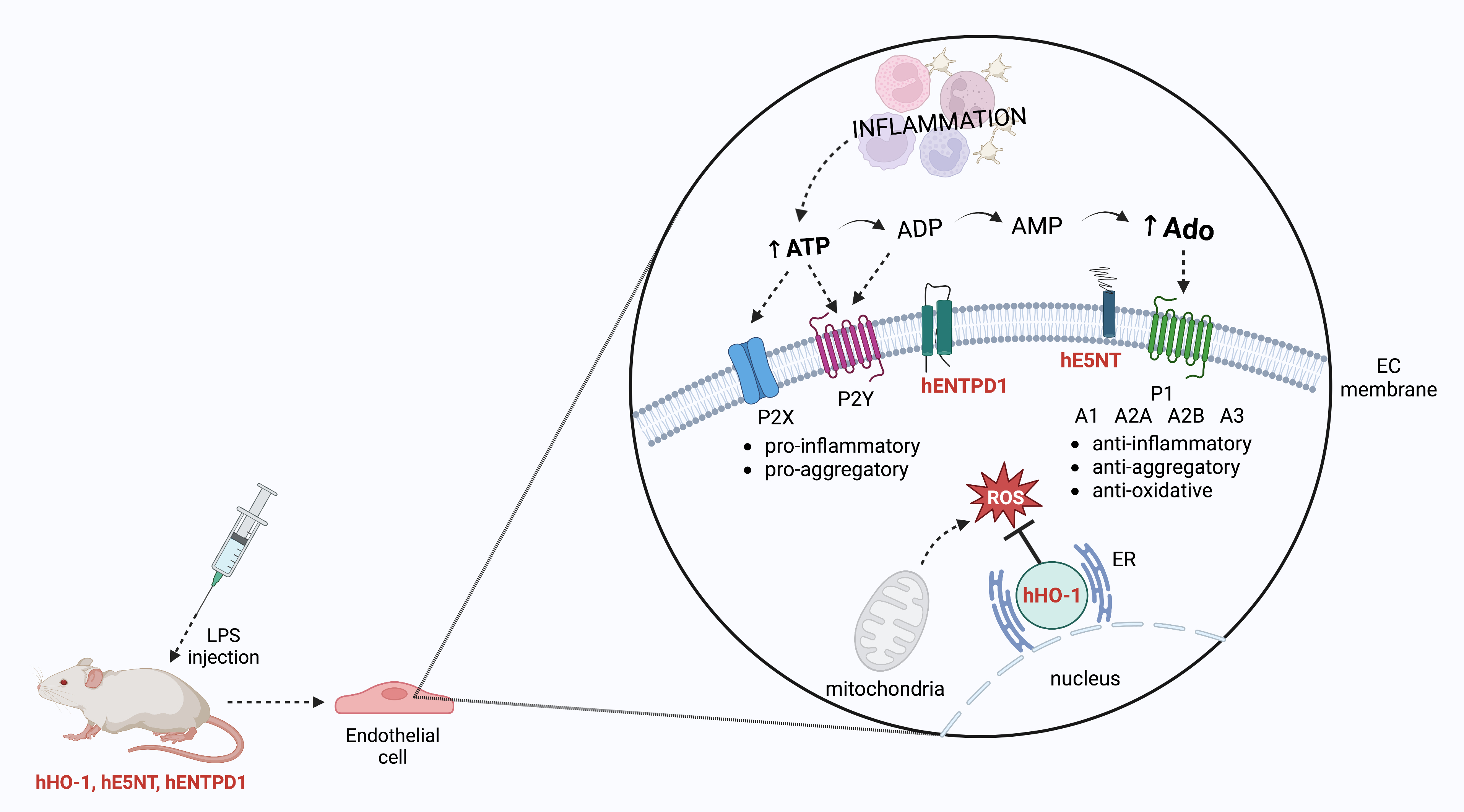
:
1. Introduction
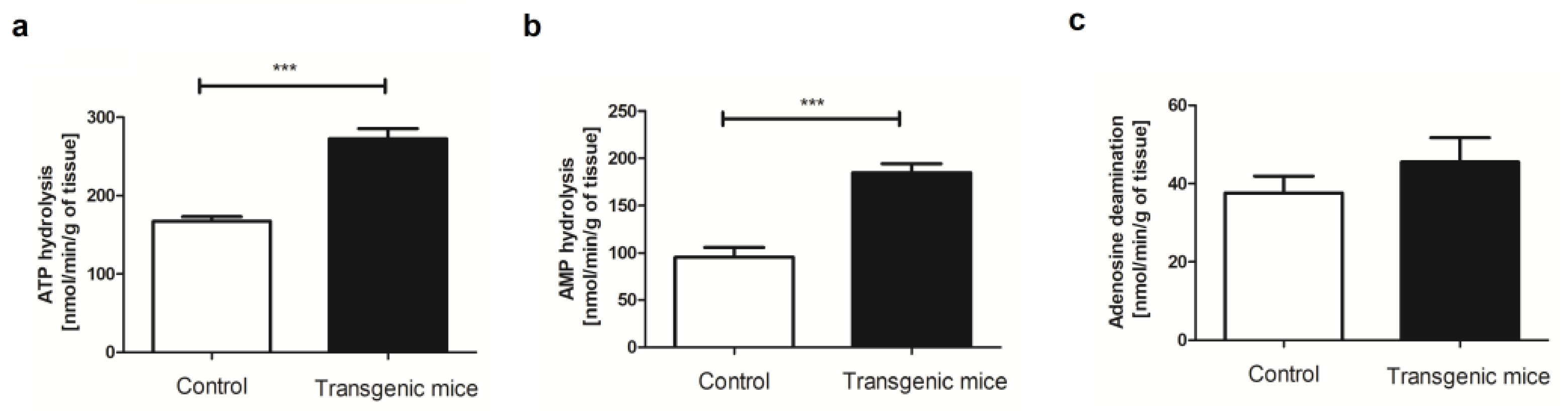
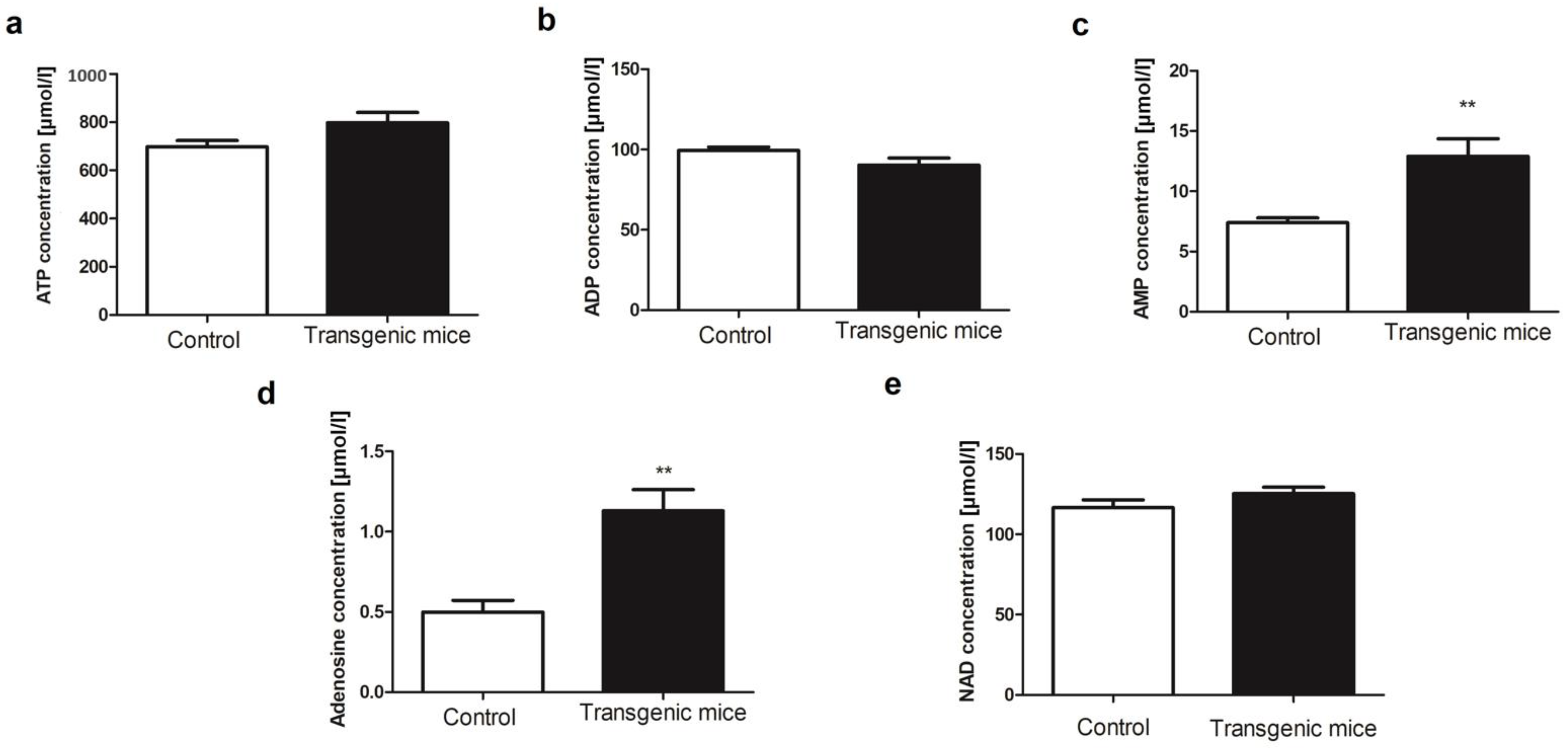
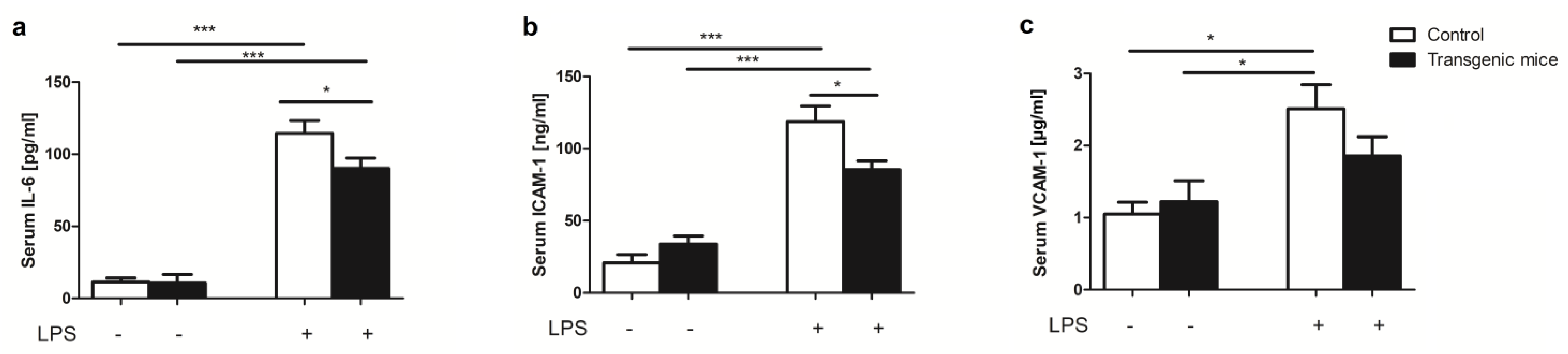
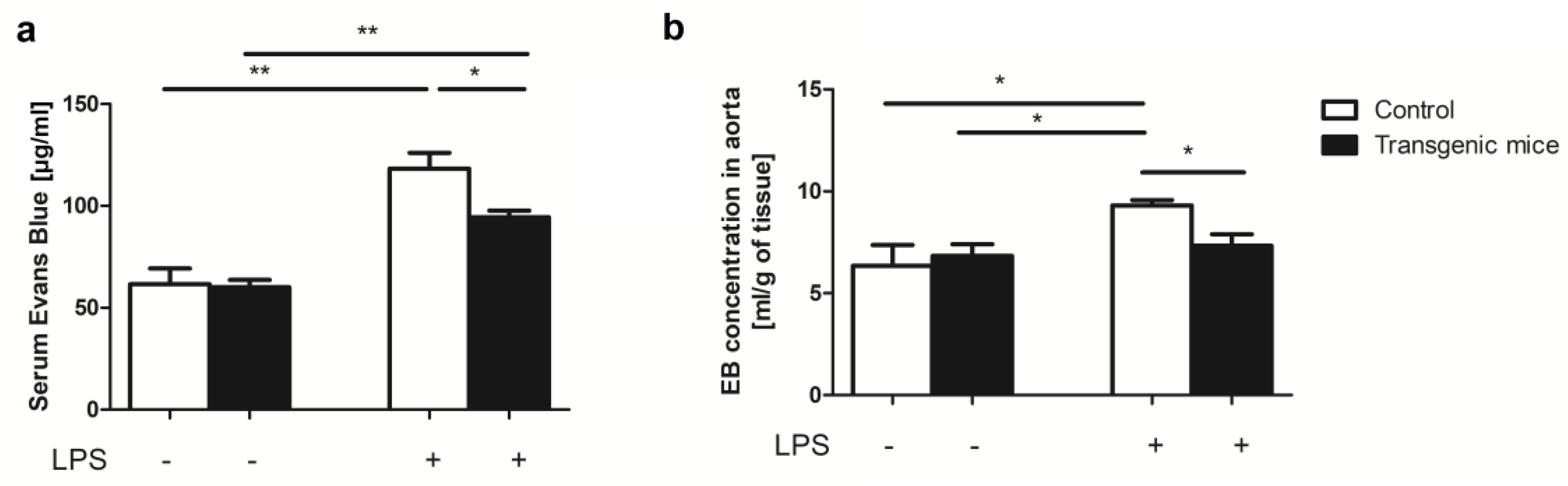
2. Results
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Triple Cistronic Plasmid Construction and Transgenic Mice Production
4.3. Animal Maintenance and Treatment
4.4. Evaluation of Extracellular Catabolism of Adenine Nucleotides on the Aortic Surface
4.5. Determination of Blood Nucleotides and Metabolites Concentration
4.6. Determination of Amino Acids and Derivatives Concentration
4.7. Intracellular Adhesion Molecule-1 (ICAM-1), Vascular Adhesion Molecule-1 (VCAM-1), Interleukin 6 (IL-6) Measurements
4.8. Evaluation of Endothelial Permeability
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lei, T.; Chen, L.; Wang, K.; Du, S.; Gonelle-Gispert, C.; Wang, Y.; Buhler, L.H. Genetic Engineering of Pigs for Xenotransplantation to Overcome Immune Rejection and Physiological Incompatibilities: The First Clinical Steps. Front. Immunol. 2022, 13, 1031185. [Google Scholar] [CrossRef] [PubMed]
- Sykes, M.; Sachs, D.H. Progress in Xenotransplantation: Overcoming Immune Barriers. Nat. Rev. Nephrol. 2022, 18, 745–761. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.K.C.; Ekser, B.; Ramsoondar, J.; Phelps, C.; Ayares, D. The Role of Genetically Engineered Pigs in Xenotransplantation Research. J. Pathol. 2016, 238, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Banz, Y.; Rieben, R. Endothelial Cell Protection in Xenotransplantation: Looking after a Key Player in Rejection. Xenotransplantation 2006, 13, 19–30. [Google Scholar] [CrossRef]
- Kummer, L.; Zaradzki, M.; Vijayan, V.; Arif, R.; Weigand, M.A.; Immenschuh, S.; Wagner, A.H.; Larmann, J. Vascular Signaling in Allogenic Solid Organ Transplantation—The Role of Endothelial Cells. Front. Physiol. 2020, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Luft, T.; Dreger, P.; Radujkovic, A. Endothelial Cell Dysfunction: A Key Determinant for the Outcome of Allogeneic Stem Cell Transplantation. Bone Marrow Transplant. 2021, 56, 2326–2335. [Google Scholar] [CrossRef] [PubMed]
- Cinti, A.; De Giorgi, M.; Chisci, E.; Arena, C.; Galimberti, G.; Farina, L.; Bugarin, C.; Rivolta, I.; Gaipa, G.; Smolenski, R.T.; et al. Simultaneous Overexpression of Functional Human HO-1, E5NT and ENTPD1 Protects Murine Fibroblasts against TNF-α-Induced Injury In Vitro. PLoS ONE 2015, 10, e0141933. [Google Scholar] [CrossRef]
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Haskó, G. CD39 and CD73 in Immunity and Inflammation. Trends Mol. Med. 2013, 19, 355–367. [Google Scholar] [CrossRef]
- Mierzejewska, P.; Zabielska, M.A.; Kutryb-Zajac, B.; Tomczyk, M.; Koszalka, P.; Smolenski, R.T.; Slominska, E.M. Impaired L-Arginine Metabolism Marks Endothelial Dysfunction in CD73-Deficient Mice. Mol. Cell Biochem. 2019, 458, 133–142. [Google Scholar] [CrossRef]
- Koszalka, P.; Ozüyaman, B.; Huo, Y.; Zernecke, A.; Flögel, U.; Braun, N.; Buchheiser, A.; Decking, U.K.M.; Smith, M.L.; Sévigny, J.; et al. Targeted Disruption of Cd73/Ecto-5′-Nucleotidase Alters Thromboregulation and Augments Vascular Inflammatory Response. Circ. Res. 2004, 95, 814–821. [Google Scholar] [CrossRef]
- Thompson, L.F.; Eltzschig, H.K.; Ibla, J.C.; Van De Wiele, C.J.; Resta, R.; Morote-Garcia, J.C.; Colgan, S.P. Crucial Role for Ecto-5′-Nucleotidase (CD73) in Vascular Leakage during Hypoxia. J. Exp. Med. 2004, 200, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Helenius, M.H.; Vattulainen, S.; Orcholski, M.; Aho, J.; Komulainen, A.; Taimen, P.; Wang, L.; de Jesus Perez, V.A.; Koskenvuo, J.W.; Alastalo, T.-P. Suppression of Endothelial CD39/ENTPD1 Is Associated with Pulmonary Vascular Remodeling in Pulmonary Arterial Hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 308, L1046–L1057. [Google Scholar] [CrossRef] [PubMed]
- Huttinger, Z.M.; Milks, M.W.; Nickoli, M.S.; Aurand, W.L.; Long, L.C.; Wheeler, D.G.; Dwyer, K.M.; d’Apice, A.J.F.; Robson, S.C.; Cowan, P.J.; et al. Ectonucleotide Triphosphate Diphosphohydrolase-1 (CD39) Mediates Resistance to Occlusive Arterial Thrombus Formation after Vascular Injury in Mice. Am. J. Pathol. 2012, 181, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.P.; Bach, F.H. Heme Oxygenase-1: From Biology to Therapeutic Potential. Trends Mol. Med. 2009, 15, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Belcher, J.D.; Mahaseth, H.; Welch, T.E.; Otterbein, L.E.; Hebbel, R.P.; Vercellotti, G.M. Heme Oxygenase-1 Is a Modulator of Inflammation and Vaso-Occlusion in Transgenic Sickle Mice. J. Clin. Investig. 2006, 116, 808–816. [Google Scholar] [CrossRef]
- De Giorgi, M.; Cinti, A.; Pelikant-Malecka, I.; Chisci, E.; Lavitrano, M.; Giovannoni, R.; Smolenski, R.T. Co-Expression of Functional Human Heme Oxygenase 1, Ecto-5′-Nucleotidase and Ecto-Nucleoside Triphosphate Diphosphohydrolase-1 by “Self-Cleaving” 2A Peptide System. Plasmid 2015, 79, 22–29. [Google Scholar] [CrossRef]
- Crikis, S.; Lu, B.; Murray-Segal, L.M.; Selan, C.; Robson, S.C.; D’Apice, A.J.F.; Nandurkar, H.H.; Cowan, P.J.; Dwyer, K.M. Transgenic Overexpression of CD39 Protects against Renal Ischemia-Reperfusion and Transplant Vascular Injury. Am. J. Transplant. 2010, 10, 2586–2595. [Google Scholar] [CrossRef]
- Minor, M.; Alcedo, K.P.; Battaglia, R.A.; Snider, N.T. Cell Type- and Tissue-Specific Functions of Ecto-5′-Nucleotidase (CD73). Am. J. Physiol. Cell Physiol. 2019, 317, C1079–C1092. [Google Scholar] [CrossRef]
- Zhou, B.; Wan, Y.; Chen, R.; Zhang, C.; Li, X.; Meng, F.; Glaser, S.; Wu, N.; Zhou, T.; Li, S.; et al. The Emerging Role of Cellular Senescence in Renal Diseases. J. Cell Mol. Med. 2020, 24, 2087–2097. [Google Scholar] [CrossRef]
- Behdad, A.; Sun, X.; Khalpey, Z.; Enjyoji, K.; Wink, M.; Wu, Y.; Usheva, A.; Robson, S.C. Vascular Smooth Muscle Cell Expression of Ectonucleotidase CD39 (ENTPD1) Is Required for Neointimal Formation in Mice. Purinergic Signal 2009, 5, 335–342. [Google Scholar] [CrossRef]
- Zukowska, P.; Kutryb-Zajac, B.; Jasztal, A.; Toczek, M.; Zabielska, M.; Borkowski, T.; Khalpey, Z.; Smolenski, R.T.; Slominska, E.M. Deletion of CD73 in Mice Leads to Aortic Valve Dysfunction. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purine and Purinergic Receptors. Brain Neurosci. Adv. 2018, 2, 2398212818817494. [Google Scholar] [CrossRef] [PubMed]
- Cronstein, B.N.; Levin, R.I.; Belanoff, J.; Weissmann, G.; Hirschhorn, R. Adenosine: An Endogenous Inhibitor of Neutrophil-Mediated Injury to Endothelial Cells. J. Clin. Investig. 1986, 78, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Kara, F.M.; Chitu, V.; Sloane, J.; Axelrod, M.; Fredholm, B.B.; Stanley, E.R.; Cronstein, B.N. Adenosine A1 Receptors (A1Rs) Play a Critical Role in Osteoclast Formation and Function. FASEB J. 2010, 24, 2325–2333. [Google Scholar] [CrossRef] [PubMed]
- Haskó, G.; Pacher, P.; Deitch, E.A.; Vizi, E.S. Shaping of Monocyte and Macrophage Function by Adenosine Receptors. Pharmacol. Ther. 2007, 113, 264–275. [Google Scholar] [CrossRef]
- Barczyk, K.; Ehrchen, J.; Tenbrock, K.; Ahlmann, M.; Kneidl, J.; Viemann, D.; Roth, J. Glucocorticoids Promote Survival of Anti-Inflammatory Macrophages via Stimulation of Adenosine Receptor A3. Blood 2010, 116, 446–455. [Google Scholar] [CrossRef]
- Allard, B.; Longhi, M.S.; Robson, S.C.; Stagg, J. The Ectonucleotidases CD39 and CD73: Novel Checkpoint Inhibitor Targets. Immunol. Rev. 2017, 276, 121–144. [Google Scholar] [CrossRef]
- Hasegawa, T.; Bouïs, D.; Liao, H.; Visovatti, S.H.; Pinsky, D.J. Ecto-5′ Nucleotidase (CD73)-Mediated Adenosine Generation and Signaling in Murine Cardiac Allograft Vasculopathy. Circ. Res. 2008, 103, 1410–1421. [Google Scholar] [CrossRef]
- St Hilaire, C.; Ziegler, S.G.; Markello, T.C.; Brusco, A.; Groden, C.; Gill, F.; Carlson-Donohoe, H.; Lederman, R.J.; Chen, M.Y.; Yang, D.; et al. NT5E Mutations and Arterial Calcifications. N. Engl. J. Med. 2011, 364, 432–442. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Chernogorova, P.; Ayata, K.; Gerlach, U.V.; Rughani, A.; Ritchey, J.W.; Ganesan, J.; Follo, M.; Zeiser, R.; Thompson, L.F.; et al. Deficiency of CD73/Ecto-5′-Nucleotidase in Mice Enhances Acute Graft-versus-Host Disease. Blood 2012, 119, 4554–4564. [Google Scholar] [CrossRef]
- Lee, S.-C.; Lee, H.; Oh, K.B.; Hwang, I.-S.; Yang, H.; Park, M.-R.; Ock, S.-A.; Woo, J.-S.; Im, G.-S.; Hwang, S. Production and Breeding of Transgenic Cloned Pigs Expressing Human CD73. Dev. Reprod. 2017, 21, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Detmer, S.A.; Chan, D.C. Functions and Dysfunctions of Mitochondrial Dynamics. Nat. Rev. Mol. Cell Biol. 2007, 8, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Köhler, D.; Eckle, T.; Faigle, M.; Grenz, A.; Mittelbronn, M.; Laucher, S.; Hart, M.L.; Robson, S.C.; Müller, C.E.; Eltzschig, H.K. CD39/Ectonucleoside Triphosphate Diphosphohydrolase 1 Provides Myocardial Protection during Cardiac Ischemia/Reperfusion Injury. Circulation 2007, 116, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, D.J.; Broekman, M.J.; Peschon, J.J.; Stocking, K.L.; Fujita, T.; Ramasamy, R.; Connolly, E.S.; Huang, J.; Kiss, S.; Zhang, Y.; et al. Elucidation of the Thromboregulatory Role of CD39/Ectoapyrase in the Ischemic Brain. J. Clin. Investig. 2002, 109, 1031–1040. [Google Scholar] [CrossRef]
- Ni, X.; Xia, Y.; Zhou, S.; Peng, H.; Wu, X.; Lu, H.; Wang, H.; Liu, R.; Blazar, B.R.; Gu, J.; et al. Reduction in Murine Acute GVHD Severity by Human Gingival Tissue-Derived Mesenchymal Stem Cells via the CD39 Pathways. Cell Death Dis. 2019, 10, 13. [Google Scholar] [CrossRef]
- Cai, Z.; Tu, L.; Guignabert, C.; Merkus, D.; Zhou, Z. Purinergic Dysfunction in Pulmonary Arterial Hypertension. J. Am. Heart Assoc. 2020, 9, e017404. [Google Scholar] [CrossRef]
- Knight, J.S.; Mazza, L.F.; Yalavarthi, S.; Sule, G.; Ali, R.A.; Hodgin, J.B.; Kanthi, Y.; Pinsky, D.J. Ectonucleotidase-Mediated Suppression of Lupus Autoimmunity and Vascular Dysfunction. Front. Immunol. 2018, 9, 1322. [Google Scholar] [CrossRef]
- He, Y.; Li, H.; Yao, J.; Zhong, H.; Kuang, Y.; Li, X.; Bian, W. HO-1 Knockdown Upregulates the Expression of VCAM-1 to Induce Neutrophil Recruitment during Renal Ischemia-reperfusion Injury. Int. J. Mol. Med. 2021, 48, 185. [Google Scholar] [CrossRef]
- Kang, L.; Hillestad, M.L.; Grande, J.P.; Croatt, A.J.; Barry, M.A.; Farrugia, G.; Katusic, Z.S.; Nath, K.A. Induction and Functional Significance of the Heme Oxygenase System in Pathological Shear Stress in Vivo. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H1402–H1413. [Google Scholar] [CrossRef]
- Kawashima, A.; Oda, Y.; Yachie, A.; Koizumi, S.; Nakanishi, I. Heme Oxygenase-1 Deficiency: The First Autopsy Case. Hum. Pathol. 2002, 33, 125–130. [Google Scholar] [CrossRef]
- Ollinger, R.; Bilban, M.; Erat, A.; Froio, A.; McDaid, J.; Tyagi, S.; Csizmadia, E.; Graça-Souza, A.V.; Liloia, A.; Soares, M.P.; et al. Bilirubin: A Natural Inhibitor of Vascular Smooth Muscle Cell Proliferation. Circulation 2005, 112, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Tulis, D.A.; Keswani, A.N.; Peyton, K.J.; Wang, H.; Schafer, A.I.; Durante, W. Local Administration of Carbon Monoxide Inhibits Neointima Formation in Balloon Injured Rat Carotid Arteries. Cell Mol. Biol. 2005, 51, 441–446. [Google Scholar]
- Singh, A.K.; Griffith, B.P.; Goerlich, C.E.; Ayares, D.; Mohiuddin, M.M. The Road to the First FDA-Approved Genetically Engineered Pig Heart Transplantation into Human. Xenotransplantation 2022, 29, e12776. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Smolenski, R.T.; Lachno, D.R.; Ledingham, S.J.; Yacoub, M.H. Determination of Sixteen Nucleotides, Nucleosides and Bases Using High-Performance Liquid Chromatography and Its Application to the Study of Purine Metabolism in Hearts for Transplantation. J. Chromatogr. 1990, 527, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.; Jeffers, L.A.; Koval, M. Measurement of Lung Vessel and Epithelial Permeability In Vivo with Evans Blue. In Permeability Barrier: Methods and Protocols; Humana: New York, NY, USA, 2021; Volume 2367. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid [μmol/L] | Control | Transgenic | p-Value |
|---|---|---|---|
| Arginine | 84.06 ± 5.18 | 92.65 ± 3.25 | 0.075 |
| ADMA | 0.91 ± 0.05 | 0.78 ± 0.04 | 0.112 |
| SDMA | 0.52 ± 0.09 | 0.50 ± 0.06 | 0.868 |
| L-NMMA | 0.36 ± 0.01 | 0.28 ± 0.02 | 0.043 * |
| Ornithine | 140.20 ± 16.48 | 147.70 ± 17.73 | 0.777 |
| Citrulline | 89.46 ± 2.98 | 98.48 ± 6.12 | 0.246 |
| Ornithine/Arginine | 1.85 ± 0.05 | 1.32 ± 0.12 | 0.012 * |
| Ornithine/Citrulline | 1.71 ± 0.13 | 1.32 ± 0.17 | 0.131 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mierzejewska, P.; Di Marzo, N.; Zabielska-Kaczorowska, M.A.; Walczak, I.; Slominska, E.M.; Lavitrano, M.; Giovannoni, R.; Kutryb-Zajac, B.; Smolenski, R.T. Endothelial Effects of Simultaneous Expression of Human HO-1, E5NT, and ENTPD1 in a Mouse. Pharmaceuticals 2023, 16, 1409. https://doi.org/10.3390/ph16101409
Mierzejewska P, Di Marzo N, Zabielska-Kaczorowska MA, Walczak I, Slominska EM, Lavitrano M, Giovannoni R, Kutryb-Zajac B, Smolenski RT. Endothelial Effects of Simultaneous Expression of Human HO-1, E5NT, and ENTPD1 in a Mouse. Pharmaceuticals. 2023; 16(10):1409. https://doi.org/10.3390/ph16101409
Chicago/Turabian StyleMierzejewska, Paulina, Noemi Di Marzo, Magdalena A. Zabielska-Kaczorowska, Iga Walczak, Ewa M. Slominska, Marialuisa Lavitrano, Roberto Giovannoni, Barbara Kutryb-Zajac, and Ryszard T. Smolenski. 2023. "Endothelial Effects of Simultaneous Expression of Human HO-1, E5NT, and ENTPD1 in a Mouse" Pharmaceuticals 16, no. 10: 1409. https://doi.org/10.3390/ph16101409