
Pharmacological Nature of the Purinergic P2Y Receptor Subtypes That Participate in the Blood Pressure Changes Produced by ADPβS in Rats
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Haemodynamic Parameters under Baseline Conditions
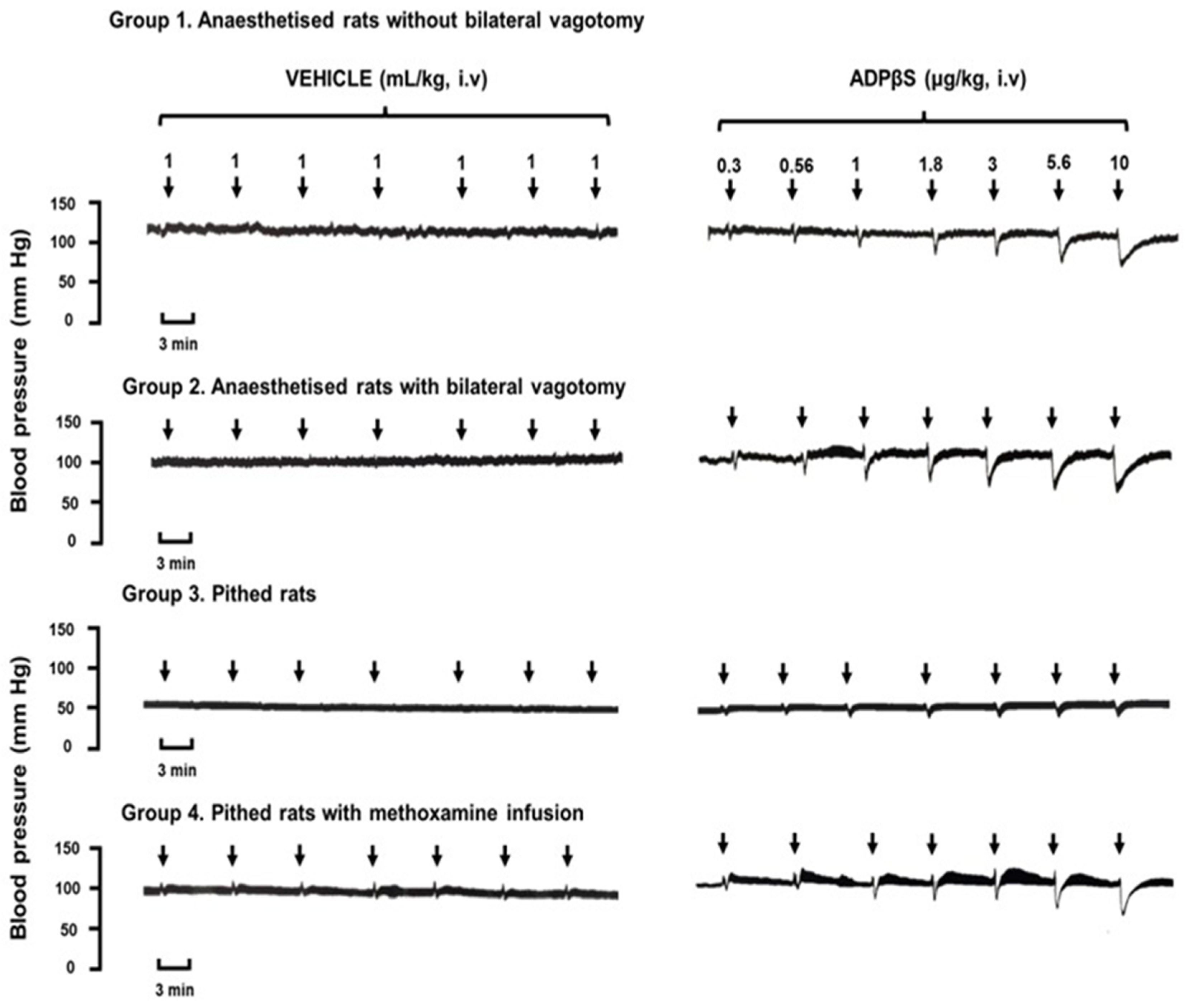
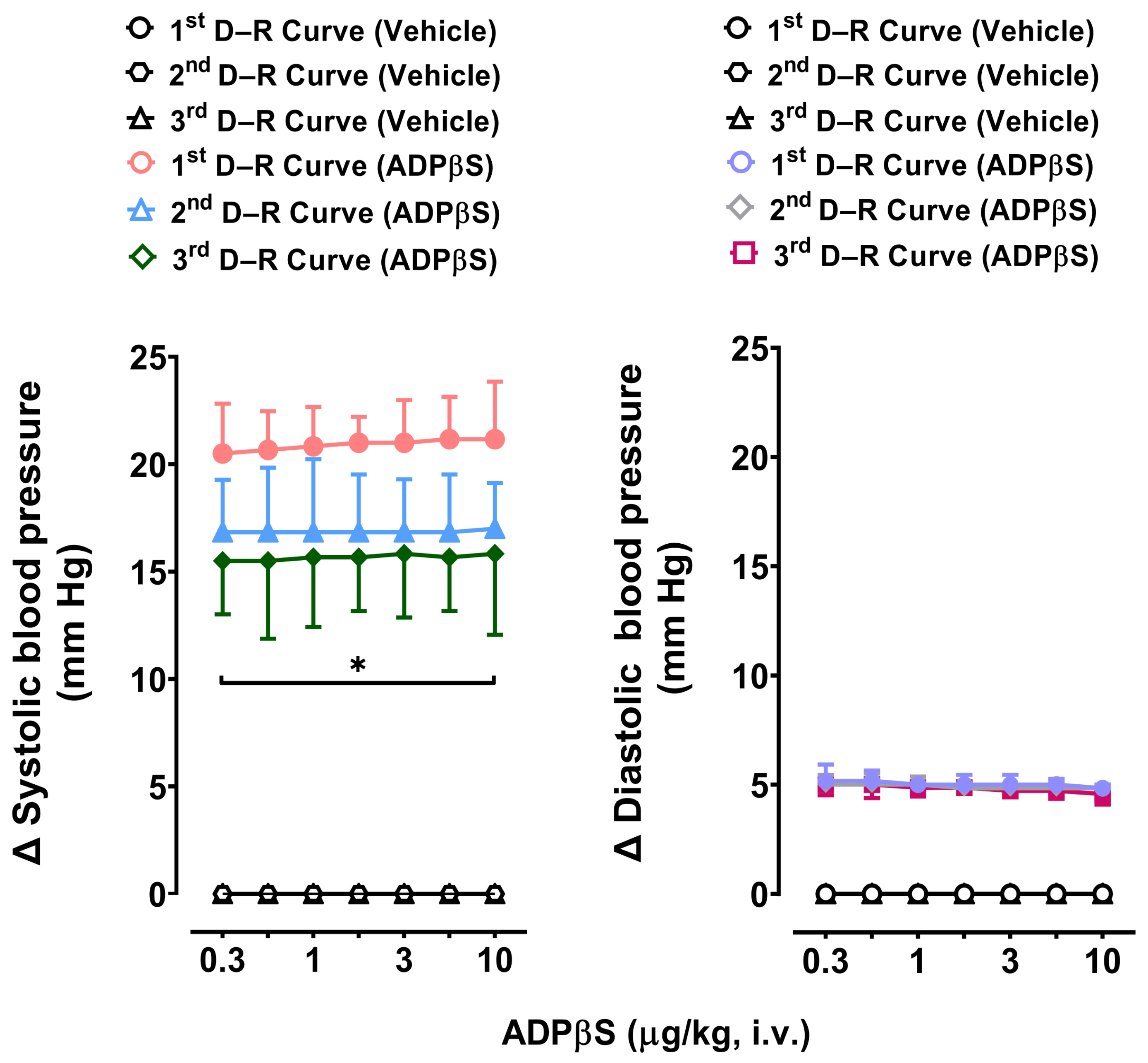
2.2. Immediate Effects of Vehicle or ADPβS on Diastolic Blood Pressure
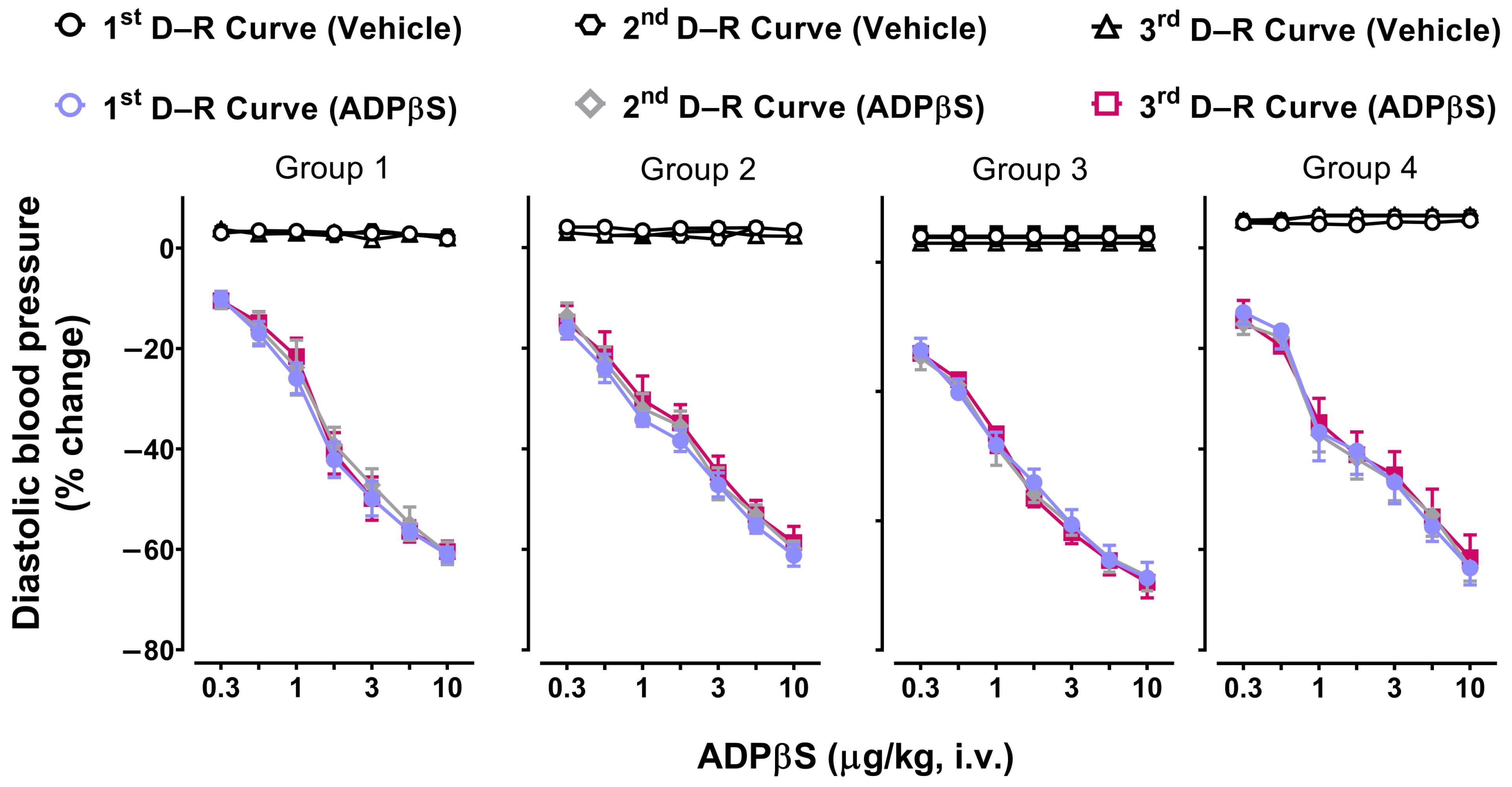
2.3. Diastolic Blood Pressure Values 10 min after Vehicle, ADPβS, or Antagonists
2.4. The Immediate Effects of Vehicle or ADPβS on Diastolic Blood Pressure
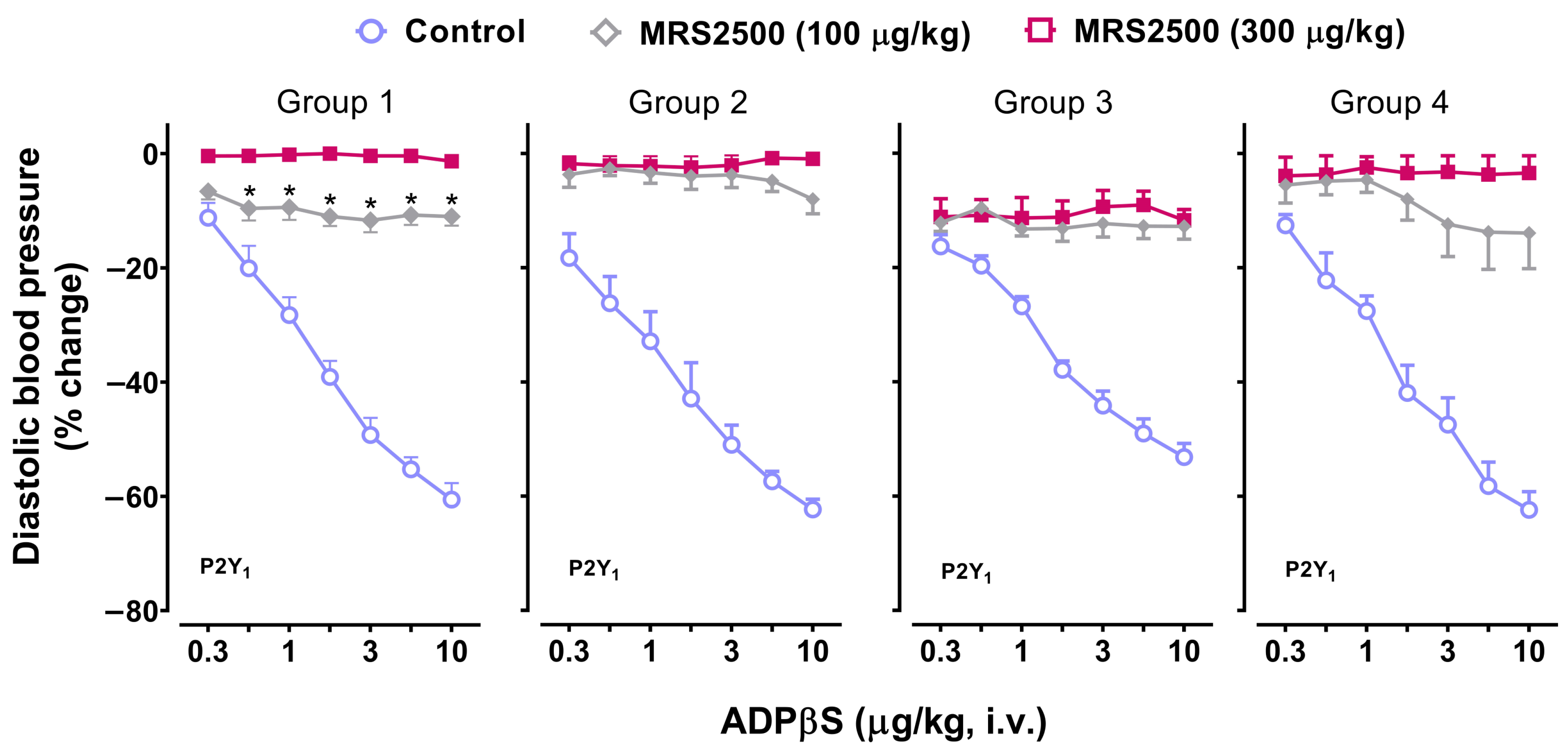
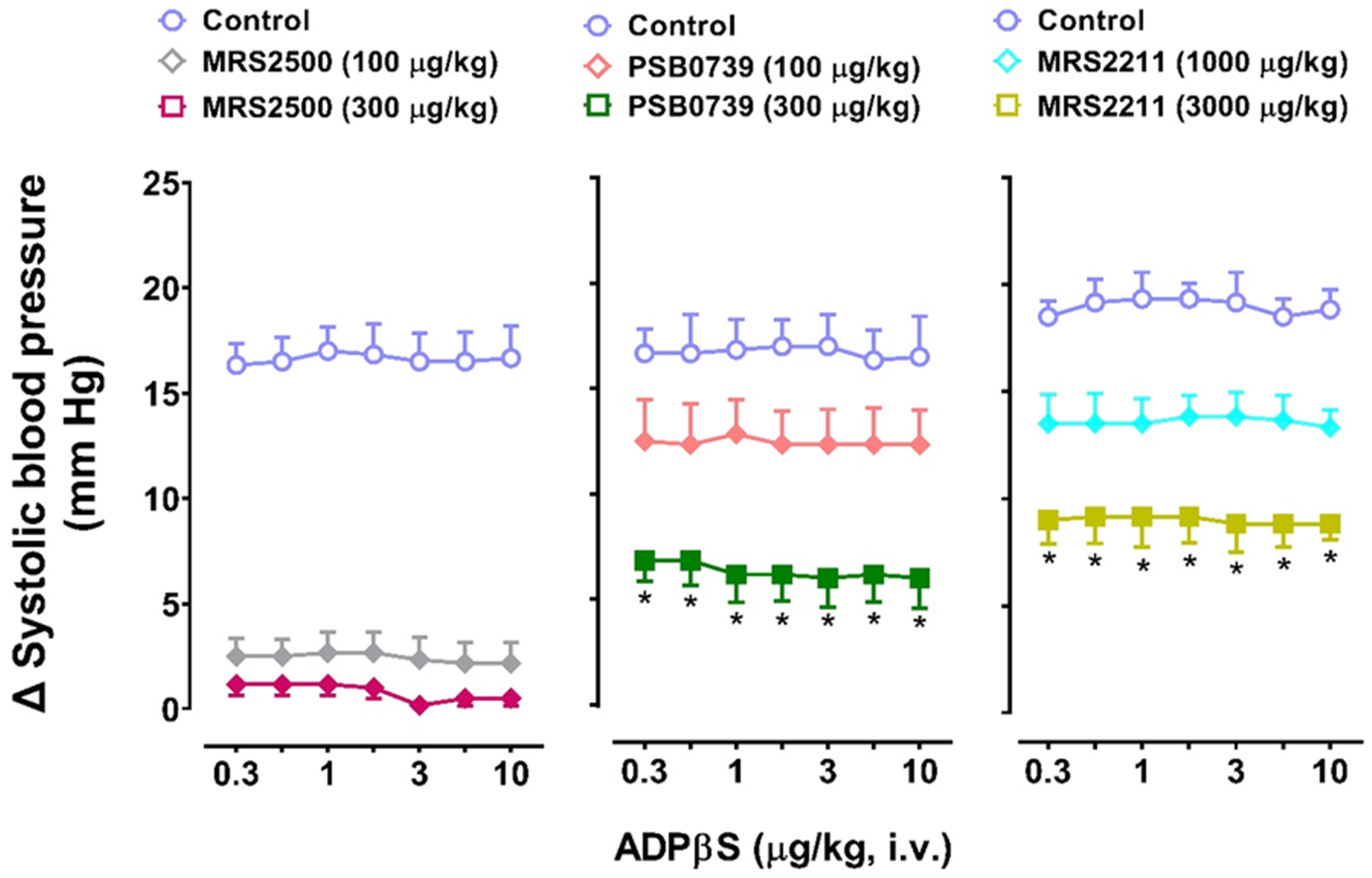
2.5. Effect of the P2Y1 Receptor Antagonist, MRS2500, on the Vasodepressor Responses to ADPβS
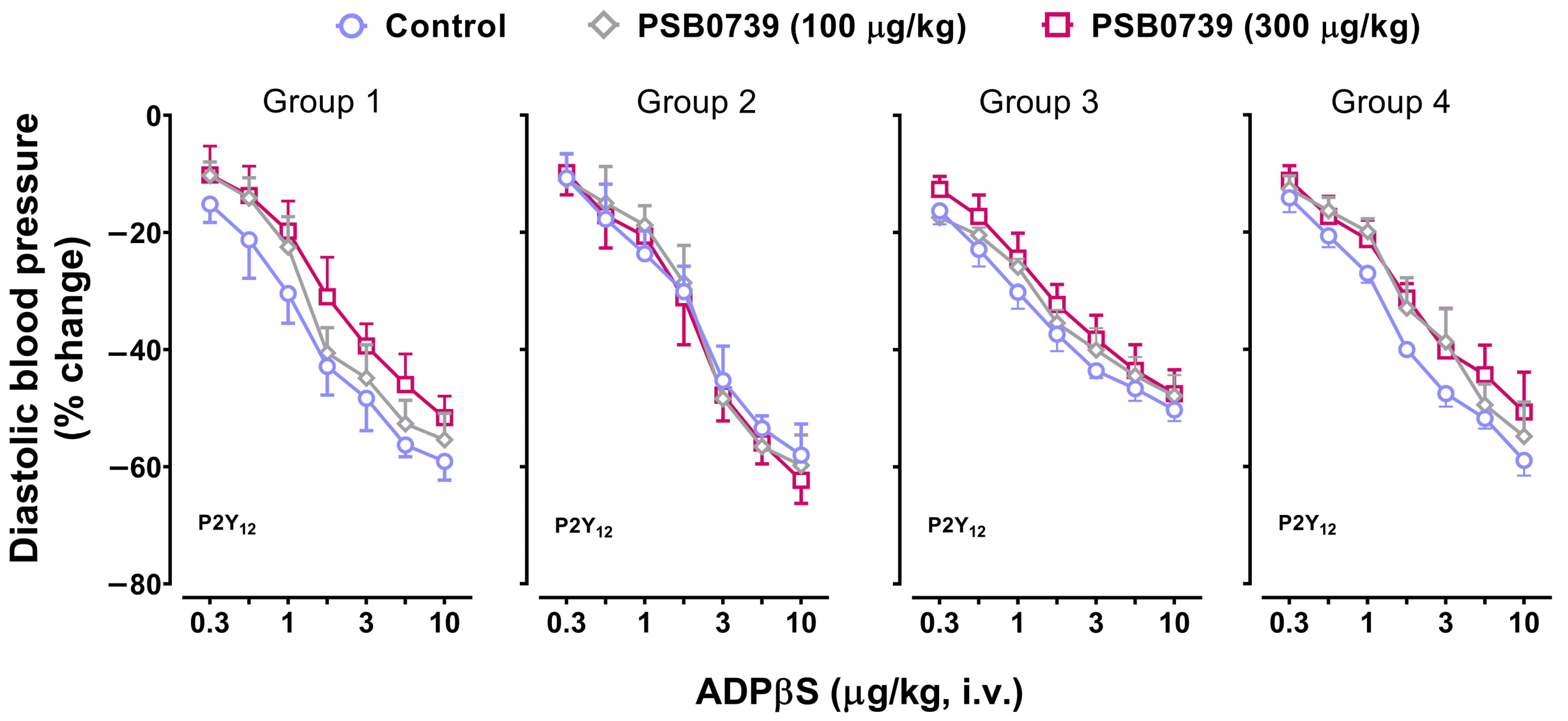
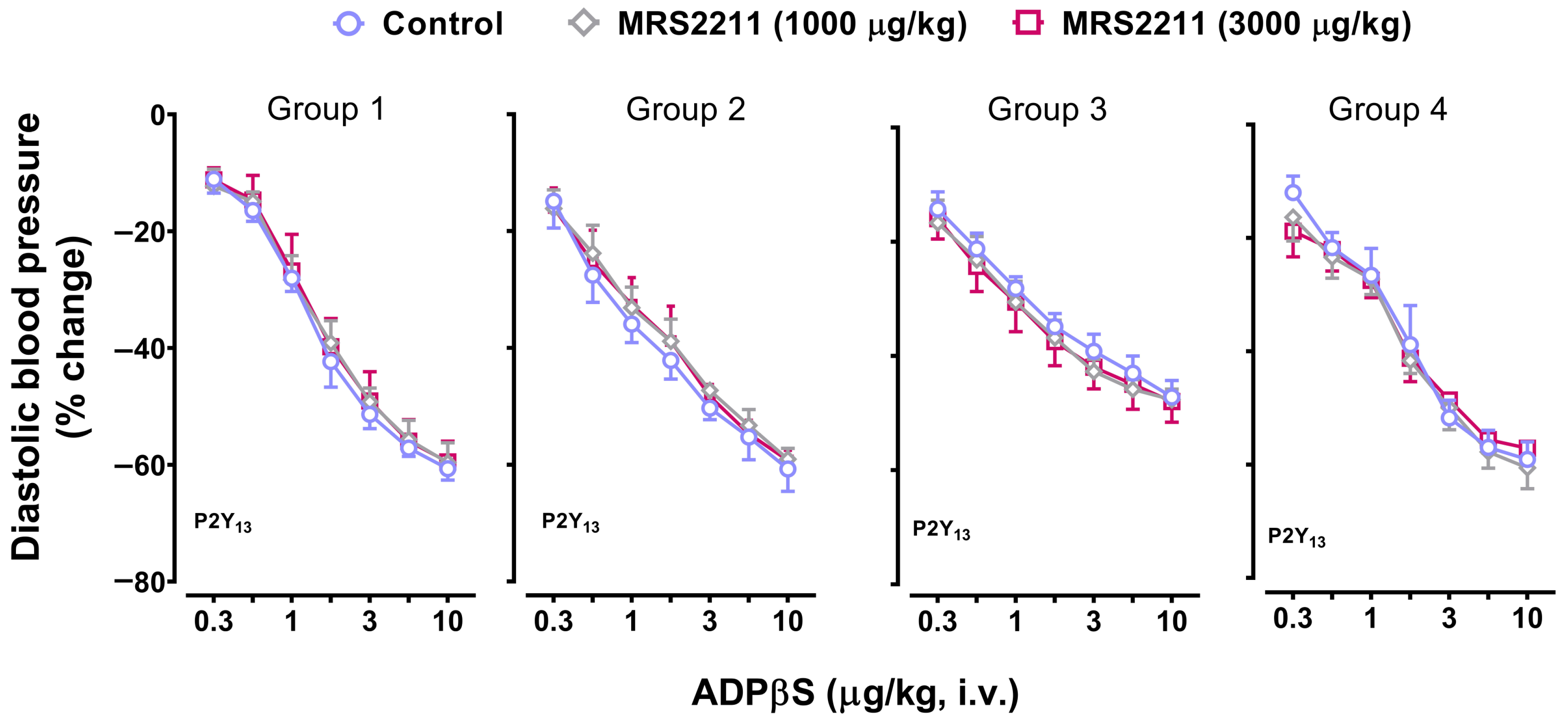
2.6. Effect of PSB0739 (a P2Y12 Receptor Antagonist) or MRS2211 (a P2Y13 Receptor Antagonist) on the Vasodepressor Responses to ADPβS
2.7. The Secondary Systolic Vasopressor Responses to ADPβS (Increases in Systolic Blood Pressure) in Pithed Rats Continuously Infused (i.v.) with Methoxamine (Group 4)
2.8. The Effects of the P2Y Receptor Antagonists MRS2500 (P2Y1), PSB0739 (P2Y12), or MRS2211 (P2Y13) on the Systolic Vasopressor Responses to ADPβS in Group 4
3. Discussion
3.1. General
3.2. Diastolic Blood Pressure Values Remained Unaffected after Administration of Vehicle, ADPβS, or Antagonists
3.3. Profile of the Responses Produced by ADPβS in the Four Groups of Animals
3.4. Reproducibility of the Vasodepressor Responses Elicited by ADPβS
3.5. Possible Participation of P2Y1, P2Y12, and P2Y13 Receptors in the Decreases in Diastolic Blood Pressure and the Increases in Systolic Blood Pressure Elicited by ADPβS
- (i)
- MRS2500, which binds with similar affinities at human and rat P2Y1 receptors [42], blocked both cardiovascular responses produced by ADPβS, namely, the decreases in DBP (i.e., systemic vasodilatation) in all four groups (Figure 3) and the increases in systolic blood pressure (i.e., cardiac left ventricular contractility) in Group 4 (Figure 7). This finding implies the role of P2Y1 receptors in both responses.
- (ii)
- PSB0739 and MRS2211 blocked the ADPβS-induced increases in systolic blood pressure (i.e., cardiac left ventricular contractility) in Group 4 (Figure 7), but not the ADPβS-induced decreases in DBP (which were blocked by MRS2500 in all four groups; Figure 3). This line of evidence suggests the additional secondary involvement of P2Y12 and P2Y13 receptors in ADPβS-induced cardiac left ventricular contractility. Within this context, since the differences in the affinity values of PSB0739 and MRS2211 for P2Y1, P2Y12, and P2Y13 receptors are around 1.5 log units (see above), one could argue that, at the i.v. doses used, the selectivity of these antagonists in pithed rats is rather low and, accordingly, that they also block (to a certain extent) P2Y1 receptors. If this were the case, however, PSB0739 and MRS2211 should have blocked the ADPβS-induced decreases in DBP (i.e., systemic vasodilatation mediated by MRS2500-sensitive P2Y1 receptors in all four groups), which was not the case (Figure 3).
3.6. Limitations of the Study
3.7. Perspectives and Potential Clinical Significance
4. Materials and Methods
4.1. Ethical Endorsement of the Experimental Protocols in Anaesthetised and Pithed Rats
4.2. General Methods
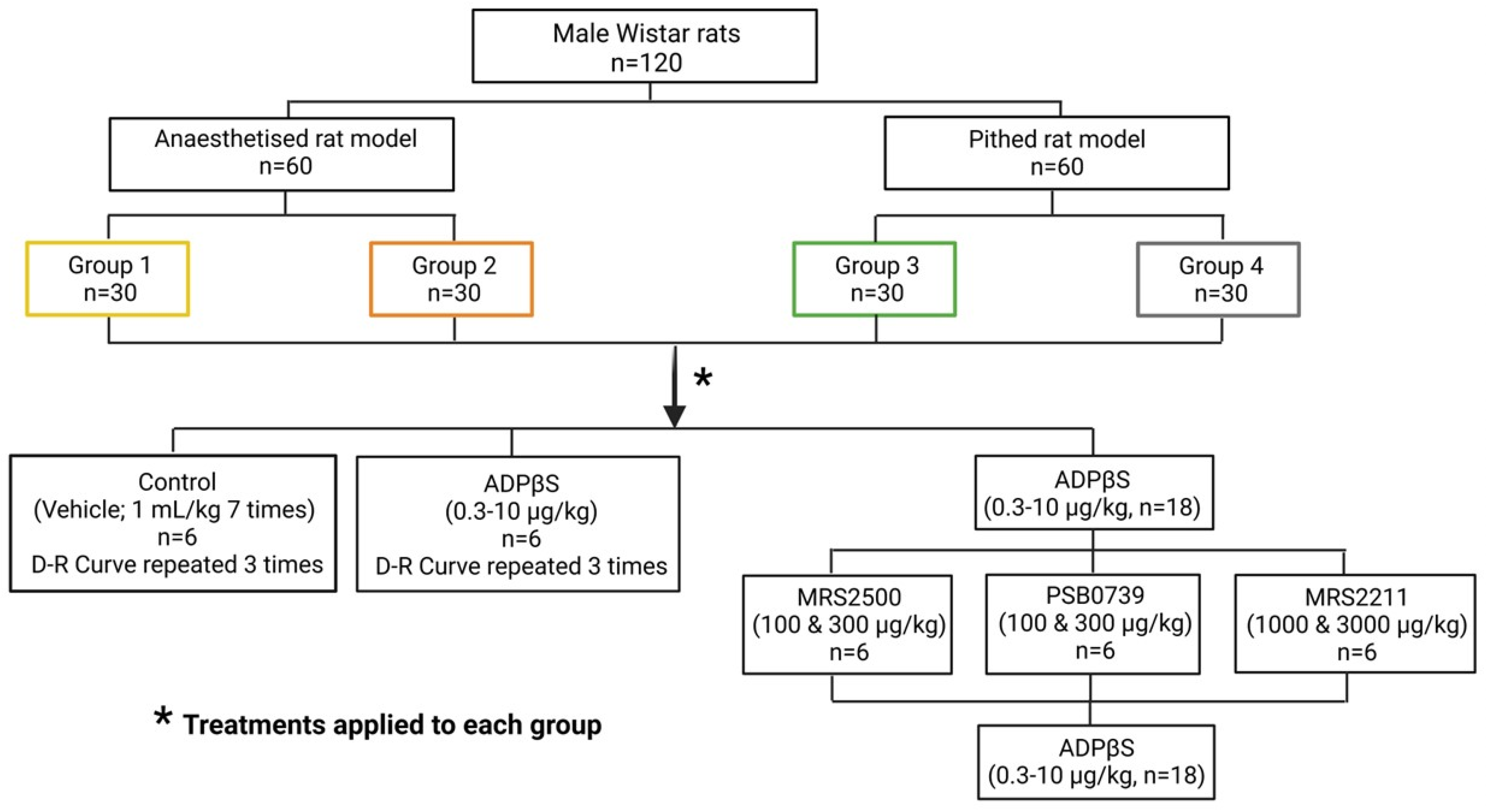
4.3. Experimental Protocols
4.3.1. Protocol I: The Effects of Consecutive i.v. Bolus of Vehicle or ADPβS on Blood Pressure
4.3.2. Protocol II: Effect of the Antagonists MRS2500 (P2Y1), PSB0739 (P2Y12), or MRS2211 (P2Y13) on the Blood Pressure Changes Caused by i.v. Bolus of ADPβS
4.4. Compounds
4.5. Presentation of Data and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADP | Adenosine diphosphate |
| ADPβS | Adenosine 5′-O-2-thiodiphosphate |
| AMP | Adenosine 5′-monophosphate |
| ATP | Adenosine 5′-triphosphate |
| cGMP | Cyclic guanosine monophosphate |
| CGRP | Calcitonin gene-related peptide |
| CNS | Central nervous system |
| DBP | Diastolic blood pressure |
| D–R curves | Dose-response curves |
| I.v. | Intravenous route of administration |
References
- Chopra, S.; Baby, C.; Jacob, J.J. Neuro-endocrine regulation of blood pressure. Indian J. Endocrinol. Metab. 2011, 15 (Suppl. 4), S281–S288. [Google Scholar] [CrossRef]
- Mulvany, M.J.; Aalkjaer, C. Structure and function of small arteries. Physiol. Rev. 1990, 70, 921–961. [Google Scholar] [CrossRef]
- Westcott, E.B.; Segal, S.S. Perivascular innervation: A multiplicity of roles in vasomotor control and myoendothelial signaling. Microcirculation 2013, 20, 217–238. [Google Scholar] [CrossRef] [PubMed]
- Westfall, T.C.M.H.; Westfall, D.P. Neurotransmission: The autonomic and Somatic Nervous Systems. In The Pharmacological Basis of Therapeutics, 11th ed.; Brunton, L.L., Lazo, J.S., Parker, K.L., Eds.; McGraw-Hill: New York, NY, USA, 2006; pp. 137–182. [Google Scholar]
- Westfall, T.C.M.H.; Westfall, D.P. Adrenergic agonists and antagonists. In The Pharmacological Basis of Therapeutics, 11th ed.; Brunton, L.L., Lazo, J.S., Parker, K.L., Eds.; McGraw-Hill: New York, NY, USA, 2006; pp. 237–292. [Google Scholar]
- Marichal-Cancino, B.A.; González-Hernández, A.; Muñoz-Islas, E.; Villalón, C.M. Monoaminergic receptors as modulators of the perivascular sympathetic and sensory CGRPergic outflows. Curr. Neuropharmacol. 2020, 18, 790–808. [Google Scholar] [CrossRef]
- Rand, M.J.; Majewski, H.; Wong-Dusting, H.; Story, D.F.; Loiacono, R.E.; Ziogas, J. Modulation of neuroeffector transmission. J. Cardiovasc. Pharmacol. 1987, 10, S33–S44. [Google Scholar] [CrossRef]
- Smillie, S.J.; Brain, S. Calcitonin gene-related peptide (CGRP) and its role in hypertension. Neuropeptides 2011, 45, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic signalling and neurological diseases: An update. CNS Neurol. Disord. Drug Targ. 2017, 16, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic signaling in the cardiovascular system. Circ. Res. 2017, 120, 207–228. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic signalling: Therapeutic developments. Front. Pharmacol. 2017, 8, 661. [Google Scholar] [CrossRef]
- Okada, S.F.; Nicholas, R.A.; Kreda, S.M.; Lazarowski, E.R.; Boucher, R.C. Physiological regulation of ATP release at the apical surface of human airway epithelia. J. Biol. Chem. 2006, 281, 22992–23002. [Google Scholar] [CrossRef]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Green, H.N.; Stoner, H.B. The effect of purine derivatives on the cardiovascular system. In Biological Actions of the Adenine Nucleotides; Green, H.N., Stoner, H.B., Eds.; H.K. Lewis: London, UK, 1950; pp. 65–103. [Google Scholar]
- Gordon, D.B.; Donald, H.H. Blood pressure lowering action of adenosine diphosphate and related compounds. Am. J. Physiol. 1961, 201, 1123–1125. [Google Scholar] [CrossRef] [PubMed]
- Dejana, E.; Castelli, M.G.; De Gaetano, G.; Bonaccorsi, A. Contribution of platelets to the cardiovascular effects of ADP in the rat. Thromb. Haemost. 1978, 39, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Haanes, K.A.; Labastida-Ramírez, A.; Blixt, F.W.; Rubio-Beltrán, E.; Dirven, C.M.; Danser, A.H.J.; Edvinsson, L.; MaassenVanDenBrink, A. Exploration of purinergic receptors as potential anti-migraine targets using established pre-clinical migraine models. Cephalalgia 2019, 39, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Von Kügelgen, I. Pharmacological profiles of cloned mammalian P2Y-receptor subtypes. Pharmacol. Ther. 2006, 110, 415–432. [Google Scholar] [CrossRef] [PubMed]
- Zizzo, M.G.; Mulè, F.; Serio, R. Activation of P2Y receptors by ATP and by its analogue, ADPbetaS, triggers two calcium signal pathways in the longitudinal muscle of mouse distal colon. Eur. J. Pharmacol. 2008, 595, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Ralevic, V. Purinergic Signaling and Blood Vessels in Health and Disease. Pharmacol. Rev. 2014, 66, 102–192. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Castillo, B.E.; Rivera-Mancilla, K.; Haanes, K.A.; MaassenVanDenBrink, A.; Villalón, C.M. The role of purinergic P2Y12 and P2Y13 receptors in ADPβS- induced inhibition of the cardioaccelerator sympathetic drive in pithed rats. Purinergic Signal. 2020, 16, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Wenker, I.C.; Sobrinho, C.R.; Takakura, A.C.; Mulkey, D.K.; Moreira, T.S. P2Y1 receptors expressed by C1 neurons determine peripheral chemoreceptor modulation of breathing, sympathetic activity, and blood pressure. Hypertension 2013, 62, 263–273. [Google Scholar] [CrossRef]
- Rajagopal, M.; Kathpalia, P.P.; Thomas, S.V.; Pao, A.C. Activation of P2Y1 and P2Y2 receptors induces chloride secretion via calcium-activated chloride channels in kidney inner medullary collecting duct cells. Am. J. Physiol. Renal. Physiol. 2011, 301, 544–553. [Google Scholar] [CrossRef]
- Kylhammar, D.; Bune, L.T.; Rådegran, G. P2Y1 and P2Y12 receptors in hypoxia- and adenosine diphosphate-induced pulmonary vasoconstriction in vivo in the pig. Eur. J. Appl. Physiol. 2014, 114, 1995–2006. [Google Scholar] [CrossRef]
- Ishida, K.; Matsumoto, T.; Taguchi, K.; Kamata, K.; Kobayashi, T. Mechanisms underlying reduced P2Y1-receptor-mediated relaxation in superior mesenteric arteries from long-term streptozotocin-induced diabetic rats. Acta Physiol. 2013, 207, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Hechler, B.; Freund, M.; Ravanat, C.; Magnenat, S.; Cazenave, J.P.; Gachet, C. Reduced atherosclerotic lesions in P2Y1/apolipoprotein E double-knockout mice: The contribution of non-hematopoietic-derived P2Y1 receptors. Circulation 2008, 118, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purine and pyrimidine receptors. Cell Mol. Life Sci. 2007, 64, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Pagoulatou, S.; Adamopoulos, D.; Rovas, G.; Bikia, V.; Stergiopulos, N. The effect of left ventricular contractility on arterial hemodynamics: A model-based investigation. PLoS ONE 2021, 16, e0255561. [Google Scholar] [CrossRef] [PubMed]
- Lefferts, W.K.; Lefferts, E.C.; Hibner, B.A.; Fernhall, B. Role of the heart and arterial tree in physiologic adjustments during exercise. In Textbook of Arterial Stiffness and Pulsatile Hemodynamics in Health and Disease, 1st ed.; Chirinos, J.A., Ed.; Academic Press/Elsevier Inc.: Cambridge, UK, 2022; pp. 527–544. [Google Scholar] [CrossRef]
- Miguel-Martínez, A.D.; Linares-Bedolla, J.; Villanueva-Castillo, B.; Haanes, K.A.; MaassenVanDenBrink, A.; Villalón, C.M. Pharmacological profile of the purinergic P2Y receptors that modulate, in response to ADPβS, the vasodepressor sensory CGRPergic outflow in pithed rats. Pharmaceuticals 2023, 16, 475. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Sympathetic purinergic transmission in small blood vessels. Trends Pharmacol. Sci. 1988, 9, 116–117. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, R.; Yarova, P.; Winter, P.; Dora, K. Desensitization of endothelial P2Y1 receptors by PKC-dependent mechanisms in pressurized rat small mesenteric arteries. Br. J. Pharmacol. 2009, 158, 1609–1620. [Google Scholar] [CrossRef]
- Saïag, B.; Hillaire-Buys, D.; Chapal, J.; Petit, P.; Pape, D.; Rault, B.; Allain, H.; Loubatières-Mariani, M.M. Study of the mechanisms involved in adenosine-5′-o-(2-thiodiphosphate) induced relaxation of rat thoracic aorta and pancreatic vascular bed. Br. J. Pharmacol. 1996, 118, 804–810. [Google Scholar] [CrossRef]
- Shipley, R.E.; Tilden, J.H. Pithed rat preparation suitable for assaying pressor substances. Proc. Soc. Exp. Biol. Med. 1947, 64, 453–455. [Google Scholar] [CrossRef]
- Centurión, D.; Glusa, E.; Sánchez-López, A.; Valdivia, L.F.; Saxena, P.R.; Villalón, C.M. 5-HT7, but not 5-HT2B, receptors mediate hypotension in vagosympathectomized rats. Eur. J. Pharmacol. 2004, 502, 239–242. [Google Scholar] [CrossRef] [PubMed]
- De Vries, P.; Villalón, C.M.; Heiligers, J.P.C.; Saxena, P.R. Nature of 5 HT1-like receptors mediating depressor responses in vagosympathectomized rats; close resemblance to the cloned 5-ht7 receptor. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1997, 356, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Rosas, V.H.; Rivera-Mancilla, E.; Marichal-Cancino, B.A.; Manrique-Maldonado, G.; Altamirano-Espinoza, A.H.; MaassenVanDenBrink, A.; Villalón, C.M. Olcegepant blocks neurogenic and non-neurogenic CGRPergic vasodepressor responses and facilitates noradrenergic vasopressor responses in pithed rats. Br. J. Pharmacol. 2017, 174, 2001–2014. [Google Scholar] [CrossRef]
- Villalón, C.M.; Albarrán-Juárez, J.A.; Lozano-Cuenca, J.; Pertz, H.H.; Görnemann, T.; Centurión, D. Pharmacological profile of the clonidine-induced inhibition of vasodepressor sensory outflow in pithed rats: Correlation with alpha (2A/2C)-adrenoceptors. Br. J. Pharmacol. 2008, 154, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Raqeeb, A.; Sheng, J.; Ao, N.; Braun, A.P. Purinergic P2Y2 receptors mediate rapid Ca2+ mobilization, membrane hyperpolarization and nitric oxide production in human vascular endothelial cells. Cell Calcium 2011, 49, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Strassheim, D.; Verin, A.; Batori, R.; Nijmeh, H.; Burns, N.; Kovacs-Kasa, A.; Umapathy, N.S.; Kotamarthi, J.; Gokhale, Y.S.; Karoor, V.; et al. P2Y purinergic receptors, endothelial dysfunction, and cardiovascular diseases. Int. J. Mol. Sci. 2020, 21, 6855. [Google Scholar] [CrossRef]
- Buvinic, S.; Poblete, M.I.; Donoso, M.V.; Delpiano, A.M.; Briones, R.; Miranda, R.; Huidobro-Toro, J.P. P2Y1 and P2Y2 receptor distribution varies along the human placental vascular tree: Role of nucleotides in vascular tone regulation. J. Physiol. 2006, 573, 427–443. [Google Scholar] [CrossRef]
- Traserra, S.; Barber, C.; Maclnnes, J.; Relea, L.; MacPherson, L.C.; Cunningham, M.R.; Vergara, P.; Accarino, A.; Kennedy, C.; Jimenez, M. Different responses of the blockade of the P2Y1 receptor with BPTU in human and porcine intestinal tissues and in cell cultures. Neurogastroenterol. Motil. 2021, 33, e14101. [Google Scholar] [CrossRef]
- Abbracchio, M.P.; Burnstock, G. Purinoceptors: Are there families of P2X and P2Y purinoceptors? Pharmacol. Ther. 1994, 64, 445–475. [Google Scholar] [CrossRef]
- Cabou, C.; Honorato, P.; Briceño, L.; Ghezali, L.; Duparc, T.; León, M.; Combes, G.; Frayssinhes, L.; Fournel, A.; Abot, A.; et al. Pharmacological inhibition of the F1-ATPase/P2Y1 pathway suppresses the effect of apolipoprotein A1 on endothelial nitric oxide synthesis and vasorelaxation. Acta Physiol. 2019, 226, 13268. [Google Scholar] [CrossRef] [PubMed]
- Cabou, C.; Martinez, L.O. The Interplay of Endothelial P2Y Receptors in Cardiovascular Health: From Vascular Physiology to Pathology. Int. J. Mol. Sci. 2022, 23, 5883. [Google Scholar] [CrossRef]
- Li, X.; Zhu, L.J.; Lv, J.; Cao, X. Purinoceptor: A novel target for hypertension. Purinergic Signal. 2023, 19, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Von Kügelgen, I. Pharmacological characterization of P2Y receptor subtypes—An update. Purinergic Signal. 2023. [Google Scholar] [CrossRef] [PubMed]
- Bird, J.E.; Wang, X.; Smith, P.L.; Barbera, F.; Huang, C.; Schumacher, W.A. A platelet target for venous thrombosis? P2Y1 deletion or antagonism protects mice from vena cava thrombosis. J. Thromb. Thrombolysis 2012, 34, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Léon, C.; Ravanat, C.; Freund, M.; Cazenave, J.P.; Gachet, C. Differential involvement of the P2Y1 and P2Y12 receptors in platelet procoagulant activity. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1941–1947. [Google Scholar] [CrossRef]
- Ribes, A.; Garcia, C.; Gratacap, M.P.; Kostenis, E.; Martínez, L.O.; Payrastre, B.; Sénard, J.M.; Galés, C.; Pons, V. Platelet P2Y1 receptor exhibits constitutive G protein signaling and β-arrestin 2 recruitment. BMC Biol. 2023, 21, 14. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Ding, Z. Role of P2Y12 receptor in thrombosis. Adv. Exp. Med. Biol. 2017, 906, 307–324. [Google Scholar] [CrossRef]
- NOM-062-ZOO-1999; Especificaciones Técnicas para la Producción, Cuidado y Uso de los Animales de Laboratorio. NORMA Oficial Mexicana: Mexico City, Mexico, 1999. Available online: https://www.fmvz.unam.mx/fmvz/principal/archivos/062ZOO.PDF (accessed on 29 November 2023).
- Bayne, K. Revised Guide for the Care and Use of Laboratory Animals Available. American Physiological Society. Physiologist 1996, 39, 208–211. [Google Scholar]
- McGrath, J.C.; Drummond, G.B.; McLachlan, E.M.; Kilkenny, C.; Wainwright, C.L. Guidelines for reporting experiments involving animals: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1573–1576. [Google Scholar] [CrossRef]
- González-Hernández, A.; Marichal-Cancino, B.A.; MaassenVanDenBrink, A.; Villalón, C.M. Serotonergic modulation of neurovascular transmission: A focus on prejunctional 5-ht receptors/mechanisms. Biomedicines 2023, 11, 1864. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biomedical Approach, 2nd ed.; McGraw-Hill, Kogakusha Ltd.: Tokyo, Japan, 1980. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Doses | DBP (mm Hg) | |||
|---|---|---|---|---|---|
| Anaesthetised Rats without Vagotomy (Group 1) | Anaesthetised Rats with Bilateral Vagotomy (Group 2) | Pithed Rats (Group 3) | Pithed Rats Continuously Infused (i.v.) with Methoxamine (Group 4) | ||
| Control | No treatment a | 104 ± 3 | 103 ± 5 | 37 ± 4 | 113 ± 4 |
| Vehicle | 1 mL/kg b | 105 ± 2 | 103 ± 5 | 38 ± 3 | 113 ± 2 |
| ADPβS | 0.3–10 µg/kg c | 105 ± 5 | 103 ± 2 | 33 ± 2 | 115 ± 4 |
| MRS2500 | 300 µg/kg | 111 ± 5 | 120 ± 5 | 46 ± 4 | 112 ± 4 |
| PSB0739 | 100 µg/kg | 114 ± 5 | 114 ± 4 | 45 ± 4 | 119 ± 5 |
| PSB0739 | 300 µg/kg | 102 ± 4 | 116 ± 3 | 43 ± 5 | 121 ± 2 |
| MRS2211 | 1000 µg/kg | 102 ± 4 | 112 ± 4 | 45 ± 5 | 120 ± 6 |
| MRS2211 | 3000 µg/kg | 104 ± 6 | 112 ± 4 | 44 ± 4 | 117 ± 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Velasco, R.C.; Villanueva-Castillo, B.; Haanes, K.A.; MaassenVanDenBrink, A.; Villalón, C.M. Pharmacological Nature of the Purinergic P2Y Receptor Subtypes That Participate in the Blood Pressure Changes Produced by ADPβS in Rats. Pharmaceuticals 2023, 16, 1683. https://doi.org/10.3390/ph16121683
Silva-Velasco RC, Villanueva-Castillo B, Haanes KA, MaassenVanDenBrink A, Villalón CM. Pharmacological Nature of the Purinergic P2Y Receptor Subtypes That Participate in the Blood Pressure Changes Produced by ADPβS in Rats. Pharmaceuticals. 2023; 16(12):1683. https://doi.org/10.3390/ph16121683
Chicago/Turabian StyleSilva-Velasco, Roberto C., Belinda Villanueva-Castillo, Kristian A. Haanes, Antoinette MaassenVanDenBrink, and Carlos M. Villalón. 2023. "Pharmacological Nature of the Purinergic P2Y Receptor Subtypes That Participate in the Blood Pressure Changes Produced by ADPβS in Rats" Pharmaceuticals 16, no. 12: 1683. https://doi.org/10.3390/ph16121683