
Aminoglycosides-Related Ototoxicity: Mechanisms, Risk Factors, and Prevention in Pediatric Patients
 ,
,
Abstract
:1. Introduction
Research Methods
2. Pharmacokinetics and Pharmacodynamics of Aminoglycosides
2.1. Aminoglycosides Structure
2.2. Aminoglycoside Absorption and Distribution
2.3. Aminoglycoside Mechanism of Action
2.4. Aminoglycoside Excretion
2.5. Aminoglycoside Pharmacokinetics in Pediatric Patients
3. Clinical Indications for the Use of Aminoglycosides
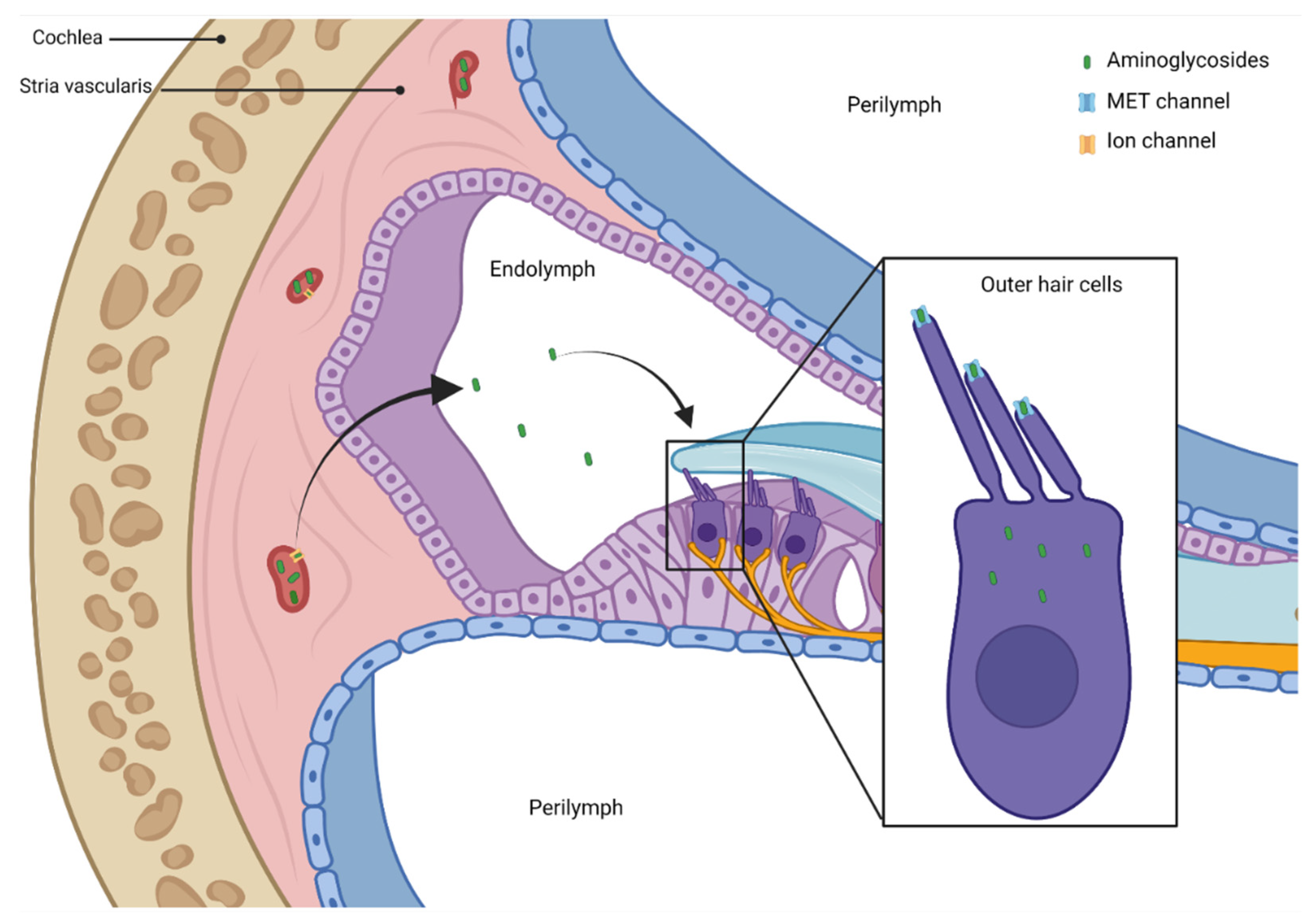
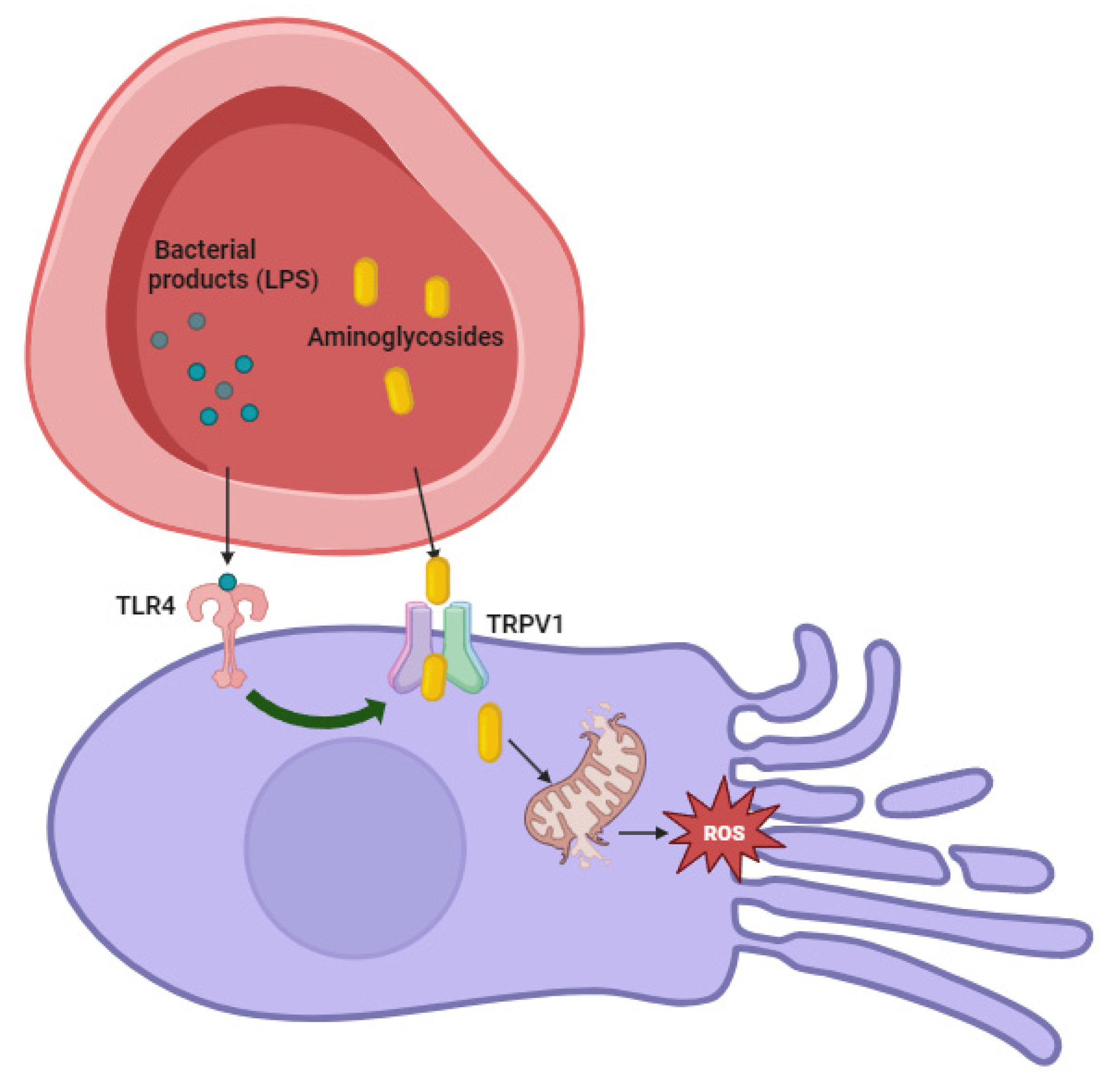
4. Pathogenesis of Aminoglycosides—Related Ototoxicity
5. Genetic Susceptibility to Aminoglycoside Ototoxicity

{kind=link}
{kind=link}
| Mutation | Population Studied | References |
|---|---|---|
| 1555A to G mutation in the 12S rRNA gene | Three Chinese pedigrees and a large Arab–Israeli pedigree with maternally inherited non-syndromic hearing loss | [78] |
| m.1555A to G and m.1494C to T in the 12S rRNA gene of the 39S subunit | Most common in Chinese pedigree, but also in Japanese, Arab–Israeli, USA families, in Mongolia and in Europe | [81,82,83] |
| 1095T to C mutation in the 12S rRNA gene | Three Chinese families | [91] |
6. Factors Enhancing the Risk of Aminoglycoside-Induced Ototoxicity
7. Prevention of Aminoglycoside Ototoxicity
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turnidge, J. Pharmacodynamics and Dosing of Aminoglycosides. Infect. Dis. Clin. N. Am. 2003, 17, 503–528. [Google Scholar] [CrossRef]
- Brunton, L.L.; Hilal-Dandan, R.; Knollmann, B.C. Goodman & Gilman’s the Pharmacological Basis of Therapeutics, 13th ed.; McGraw-Hill Education: New York, NY, USA, 2018. [Google Scholar]
- Cox, G.; Ejim, L.; Stogios, P.J.; Koteva, K.; Bordeleau, E.; Evdokimova, E.; Sieron, A.O.; Savchenko, A.; Serio, A.W.; Krause, K.M.; et al. Plazomicin Retains Antibiotic Activity against Most Aminoglycoside Modifying Enzymes. ACS Infect. Dis. 2018, 4, 980–987. [Google Scholar] [CrossRef]
- Kearns, G.L.; Abdel-Rahman, S.M.; Alander, S.W.; Blowey, D.L.; Leeder, J.S.; Kauffman, R.E. Developmental Pharmacology—Drug Disposition, Action, and Therapy in Infants and Children. N. Engl. J. Med. 2003, 349, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Taber, H.W.; Mueller, J.P.; Miller, P.F.; Arrow, A.S. Bacterial Uptake of Aminoglycoside Antibiotics. Microbiol. Rev. 1987, 51, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.D.; Chen, L.L.; Tai, P.C. Misread Protein Creates Membrane Channels: An Essential Step in the Bactericidal Action of Aminoglycosides. Proc. Natl. Acad. Sci. USA 1986, 83, 6164–6168. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Blanchard, J.S. Molecular Insights into Aminoglycoside Action and Resistance. Chem. Rev. 2005, 105, 477–498. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.B.; Terry, D.S.; Altman, R.B.; Blanchard, S.C. Aminoglycoside Activity Observed on Single Pre-Translocation Ribosome Complexes. Nat. Chem. Biol. 2010, 6, 54–62. [Google Scholar] [CrossRef]
- Misumi, M.; Nishimura, T.; Komai, T.; Tanaka, N. Interaction of Kanamycin and Related Antibiotics with the Large Subunit of Ribosomes and the Inhibition of Translocation. Biochem. Biophys. Res. Commun. 1978, 84, 358–365. [Google Scholar] [CrossRef]
- Okuyama, A.; Watanabe, T.; Tanaka, N. Effects of aminoglycoside antibiotics on initiation of viral RNA-directed protein synthesis. J. Antibiot. 1972, 25, 212–218. [Google Scholar] [CrossRef]
- Bryan, L.E.; Kwan, S. Roles of Ribosomal Binding, Membrane Potential, and Electron Transport in Bacterial Uptake of Streptomycin and Gentamicin. Antimicrob. Agents Chemother. 1983, 23, 835–845. [Google Scholar] [CrossRef]
- Davis, B.D. Mechanism of Bactericidal Action of Aminoglycosides. Microbiol. Rev. 1987, 51, 341–350. [Google Scholar] [CrossRef]
- Germovsek, E.; Barker, C.I.; Sharland, M. What Do I Need to Know about Aminoglycoside Antibiotics? Arch. Dis. Child. Educ. Pract. Ed. 2017, 102, 89–93. [Google Scholar] [CrossRef]
- Craig, W.A. Optimizing Aminoglycoside Use. Crit. Care Clin. 2011, 27, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Chisholm-Burns, M. Pharmacotherapy Principles & Practice, 4th ed.; McGraw-Hill Medical: New York, NY, USA, 2016. [Google Scholar]
- Abdulla, A.; Edwina, E.; Flint, R.B.; Allegaert, K.; Wildschut, E.D.; Koch, B.C.P.; de Hoog, M. Model-Informed Precision Dosing of Antibiotics in Pediatric Patients: A Narrative Review. Front. Pediatr. 2021, 9, 624639. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, R.H.; Thabit, A.K.; Lashkar, M.O.; Ali, A.S. Optimizing gentamicin dosing in different pediatric age groups using population pharmacokinetics and Monte Carlo simulation. Ital. J. Pediatr. 2021, 47, 167. [Google Scholar] [CrossRef]
- Gijsen, M.; Vlasselaers, D.; Spriet, I.; Allegaert, K. Pharmacokinetics of Antibiotics in Pediatric Intensive Care: Fostering Variability to Attain Precision Medicine. Antibiotics 2021, 10, 1182. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, K.; Navaswetha, T.; Vasudevan, H.; Kumar, S.N.; Arun, K.P. Review on Population Pharmacokinetics of Amikacin in Paediatrics. J. Pure Appl. Microbiol. 2022, 16, 2303–2309. [Google Scholar] [CrossRef]
- Zazo, H.; Lagarejos, E.; Prado-Velasco, M.; Sánchez-Herrero, S.; Serna, J.; Rueda-Ferreiro, A.; Martín-Suárez, A.; Calvo, M.V.; Pérez-Blanco, J.S.; Lanao, J.M. Physiologically-based pharmacokinetic modelling and dosing evaluation of gentamicin in neonates using PhysPK. Front. Pharmacol. 2022, 13, 977372. [Google Scholar] [CrossRef]
- Dong, M.; Rodriguez, A.V.; Blankenship, C.A.; McPhail, G.; Vinks, A.A.; Hunter, L.L. Pharmacokinetic modelling to predict risk of ototoxicity with intravenous tobramycin treatment in cystic fibrosis. J. Antimicrob. Chemother. 2021, 76, 2923–2931. [Google Scholar] [CrossRef]
- Sader, H.S.; Farrell, D.J.; Flamm, R.K.; Jones, R.N. Antimicrobial Susceptibility of Gram-Negative Organisms Isolated from Patients Hospitalised with Pneumonia in US and European Hospitals: Results from the SENTRY Antimicrobial Surveillance Program, 2009–2012. Int. J. Antimicrob. Agents 2014, 43, 328–334. [Google Scholar] [CrossRef]
- Landman, D.; Kelly, P.; Backer, M.; Babu, E.; Shah, N.; Bratu, S.; Quale, J. Antimicrobial Activity of a Novel Aminoglycoside, ACHN-490, against Acinetobacter baumannii and Pseudomonas aeruginosa from New York City. J. Antimicrob. Chemother. 2010, 66, 332–334. [Google Scholar] [CrossRef] [PubMed]
- Swenson, J.M.; Wallace, R.J.; Silcox, V.A.; Thornsberry, C. Antimicrobial Susceptibility of Five Subgroups of Mycobacterium Fortuitum and Mycobacterium Chelonae. Antimicrob. Agents Chemother. 1985, 28, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, G.M.; Eliopoulos, C.T. Antibiotic Combinations: Should They Be Tested? Clin. Microbiol. Rev. 1988, 1, 139–156. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.; Rhodes, A.; Alhazzani, W.; Antonelli, M.; Coopersmith, C.M.; French, C.; Machado, F.R.; Mcintyre, L.; Ostermann, M.; Prescott, H.C.; et al. Surviving Sepsis Campaign: International Guidelines for Management of Sepsis and Septic Shock 2021. Intensive Care Med. 2021, 47, 1181–1247. [Google Scholar] [CrossRef]
- Weiss, S.L.; Peters, M.J.; Alhazzani, W.; Agus, M.S.D.; Flori, H.R.; Inwald, D.P.; Nadel, S.; Schlapbach, L.J.; Tasker, R.C.; Argent, A.C.; et al. Surviving Sepsis Campaign International Guidelines for the Management of Septic Shock and Sepsis-Associated Organ Dysfunction in Children. Pediatr. Crit. Care Med. 2020, 21, e52–e106. [Google Scholar] [CrossRef]
- Lehrnbecher, T.; Robinson, P.; Fisher, B.; Alexander, S.; Ammann, R.A.; Beauchemin, M.; Carlesse, F.; Groll, A.H.; Haeusler, G.M.; Santolaya, M.; et al. Guideline for the Management of Fever and Neutropenia in Children with Cancer and Hematopoietic Stem-Cell Transplantation Recipients: 2017 Update. J. Clin. Oncol. 2017, 35, 2082–2094. [Google Scholar] [CrossRef]
- Puopolo, K.M.; Benitz, W.E.; Zaoutis, T.E. Management of Neonates Born at ≥35 0/7 Weeks’ Gestation with Suspected or Proven Early-Onset Bacterial Sepsis. Pediatrics 2018, 142, e20182894. [Google Scholar] [CrossRef]
- Rao, S.C.; Srinivasjois, R.; Moon, K. One Dose per Day Compared to Multiple Doses per Day of Gentamicin for Treatment of Suspected or Proven Sepsis in Neonates. Cochrane Database Syst. Rev. 2016, 12, CD005091. [Google Scholar] [CrossRef]
- American Academy of Pediatrics. Tables of antibacterial drug dosages. In Red Book: Report of the Committee on Infectious Diseases; American Academy of Pediatrics: Elk Grove Village, IL, USA, 2021; p. 876. [Google Scholar]
- Stein, R.; Dogan, H.S.; Hoebeke, P.; Kočvara, R.; Nijman, R.J.M.; Radmayr, C.; Tekgül, S. Urinary Tract Infections in Children: EAU/ESPU Guidelines. Eur. Urol. 2015, 67, 546–558. [Google Scholar] [CrossRef]
- Doré-Bergeron, M.-J.; Gauthier, M.; Chevalier, I.; McManus, B.; Tapiero, B.; Lebrun, S. Urinary Tract Infections in 1- to 3-Month-Old Infants: Ambulatory Treatment with Intravenous Antibiotics. Pediatrics 2009, 124, 16–22. [Google Scholar] [CrossRef]
- Gauthier, M. Treatment of Urinary Tract Infections among Febrile Young Children with Daily Intravenous Antibiotic Therapy at a Day Treatment Center. Pediatrics 2004, 114, e469–e476. [Google Scholar] [CrossRef] [PubMed]
- Elson, E.C.; Meier, E.; Oermann, C.M. The Implementation of an Aminoglycoside Induced Ototoxicity Algorithm for People with Cystic Fibrosis. J. Cyst. Fibros. 2020, 20, 284–287. [Google Scholar] [CrossRef]
- Garinis, A.C.; Liao, S.; Cross, C.P.; Galati, J.; Middaugh, J.L.; Mace, J.C.; Wood, A.; McEvoy, L.; Moneta, L.B.; Lubianski, T.; et al. Effect of Gentamicin and Levels of Ambient Sound on Hearing Screening Outcomes in the Neonatal Intensive Care Unit: A Pilot Study. Int. J. Pediatr. Otorhinolaryngol. 2017, 97, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Diepstraten, F.A.; Hoetink, A.E.; van Grotel, M.; Huitema, A.D.R.; Stokroos, R.J.; van den Heuvel-Eibrink, M.M.; Meijer, A.J.M. Aminoglycoside- and glycopeptide-induced ototoxicity in children: A systematic review. JAC-Antimicrob. Resist. 2021, 3, dlab184. [Google Scholar] [CrossRef]
- Lanvers-Kaminsky, C.; Zehnhoff-Dinnesen, A.a.; Parfitt, R.; Ciarimboli, G. Drug-Induced Ototoxicity: Mechanisms, Pharmacogenetics, and Protective Strategies. Clin. Pharmacol. Ther. 2017, 101, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Talaska, A.E.; Schacht, J. New Developments in Aminoglycoside Therapy and Ototoxicity. Hear. Res. 2011, 281, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Van Hecke, R.; Van Rompaey, V.; Wuyts, F.L.; Leyssens, L.; Maes, L. Systemic Aminoglycosides-Induced Vestibulotoxicity in Humans. Ear Hear. 2017, 38, 653–662. [Google Scholar] [CrossRef]
- Nyberg, S.; Abbott, N.J.; Shi, X.; Steyger, P.S.; Dabdoub, A. Delivery of Therapeutics to the Inner Ear: The Challenge of the Blood-Labyrinth Barrier. Sci. Transl. Med. 2019, 11, eaao0935. [Google Scholar] [CrossRef]
- Tran Ba Huy, P.; Bernard, P.; Schacht, J. Kinetics of Gentamicin Uptake and Release in the Rat. Comparison of Inner Ear Tissues and Fluids with Other Organs. J. Clin. Investig. 1986, 77, 1492–1500. [Google Scholar] [CrossRef]
- Li, H.; Steyger, P.S. Systemic Aminoglycosides Are Trafficked via Endolymph into Cochlear Hair Cells. Sci. Rep. 2011, 1, 159. [Google Scholar] [CrossRef]
- Koo, J.-W.; Quintanilla-Dieck, L.; Jiang, M.; Liu, J.; Urdang, Z.D.; Allensworth, J.J.; Cross, C.P.; Li, H.; Steyger, P.S. Endotoxemia-Mediated Inflammation Potentiates Aminoglycoside-Induced Ototoxicity. Sci. Transl. Med. 2015, 7, 298ra118. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.F.; Steyger, P.S. A Systemic Gentamicin Pathway across the Stria Vascularis. Hear. Res. 2008, 235, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kachelmeier, A.; Dai, C.; Li, H.; Steyger, P.S. Uptake of Fluorescent Gentamicin by Peripheral Vestibular Cells after Systemic Administration. PLoS ONE 2015, 10, e0120612. [Google Scholar] [CrossRef] [PubMed]
- Marcotti, W.; Van Netten, S.M.; Kros, C.J. The Aminoglycoside Antibiotic Dihydrostreptomycin Rapidly Enters Mouse Outer Hair Cells through the Mechano-Electrical Transducer Channels. J. Physiol. 2005, 567, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Alharazneh, A.; Luk, L.; Huth, M.; Monfared, A.; Steyger, P.S.; Cheng, A.G.; Ricci, A.J. Functional Hair Cell Mechanotransducer Channels Are Required for Aminoglycoside Ototoxicity. PLoS ONE 2011, 6, e22347. [Google Scholar] [CrossRef]
- van Netten, S.M.; Kros, C.J. Insights into the Pore of the Hair Cell Transducer Channel from Experiments with Permeant Blockers. Curr. Top Membr. 2007, 59, 375–398. [Google Scholar] [CrossRef]
- Pan, B.; Akyuz, N.; Liu, X.-P.; Asai, Y.; Nist-Lund, C.; Kurima, K.; Derfler, B.H.; György, B.; Limapichat, W.; Walujkar, S.; et al. TMC1 Forms the Pore of Mechanosensory Transduction Channels in Vertebrate Inner Ear Hair Cells. Neuron 2018, 99, 736–753.e6. [Google Scholar] [CrossRef]
- Fettiplace, R.; Kim, K.X. The Physiology of Mechanoelectrical Transduction Channels in Hearing. Physiol. Rev. 2014, 94, 951–986. [Google Scholar] [CrossRef]
- Leitner, M.G.; Halaszovich, C.R.; Oliver, D. Aminoglycosides Inhibit KCNQ4 Channels in Cochlear Outer Hair Cells via Depletion of Phosphatidylinositol(4,5)Bisphosphate. Mol. Pharmacol. 2010, 79, 51–60. [Google Scholar] [CrossRef]
- Nilius, B.; Szallasi, A. Transient Receptor Potential Channels as Drug Targets: From the Science of Basic Research to the Art of Medicine. Pharmacol. Rev. 2014, 66, 676–814. [Google Scholar] [CrossRef]
- Forge, A.; Schacht, J. Aminoglycoside Antibiotics. Audiol. Neuro-Otol. 2000, 5, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Leonova, E.V.; Raphael, Y. Organization of Cell Junctions and Cytoskeleton in the Reticular Lamina in Normal and Ototoxically Damaged Organ of Corti. Hear. Res. 1997, 113, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Forge, A.; Wright, A.; Davies, S.J. Analysis of Structural Changes in the Stria Vascularis Following Chronic Gentamicin Treatment. Hear. Res. 1987, 31, 253–265. [Google Scholar] [CrossRef]
- Xiong, H.; Chu, H.; Zhou, X.; Huang, X.; Cui, Y.; Zhou, L.; Chen, J.; Li, J.; Wang, Y.; Chen, Q.; et al. Simultaneously Reduced NKCC1 and Na,K-ATPase Expression in Murine Cochlear Lateral Wall Contribute to Conservation of Endocochlear Potential Following a Sensorineural Hearing Loss. Neurosci. Lett. 2011, 488, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, C.; Erostegui, C.; Sugasawa, M.; Dulon, D. Gentamicin Blocks ACh-Evoked K+ Current in Guinea-Pig Outer Hair Cells by Impairing Ca2+ Entry at the Cholinergic Receptor. J. Physiol. 2000, 525 Pt 3, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Avan, P.; Erre, J.-P.; da Costa, D.L.; Aran, J.-M.; Popelár, J. The Efferent-Mediated Suppression of Otoacoustic Emissions in Awake Guinea Pigs and Its Reversible Blockage by Gentamicin. Exp. Brain Res. 1996, 109, 9–16. [Google Scholar] [CrossRef]
- Schacht, J.; Lodhi, S.; Weiner, N.D. Effects of Neomycin on Polyphosphoinositides in Inner Ear Tissues and Monomolecular Films. In Membrane Toxicity; Springer: New York, NY, USA, 1977; pp. 191–208. [Google Scholar] [CrossRef]
- Liu, K.; Jiang, X.; Shi, C.; Shi, L.; Yang, B.; Shi, L.; Xu, Y.; Yang, W.; Yang, S. Cochlear Inner Hair Cell Ribbon Synapse Is the Primary Target of Ototoxic Aminoglycoside Stimuli. Mol. Neurobiol. 2013, 48, 647–654. [Google Scholar] [CrossRef]
- Schacht, J.; Talaska, A.E.; Rybak, L.P. Cisplatin and Aminoglycoside Antibiotics: Hearing Loss and Its Prevention. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2012, 295, 1837–1850. [Google Scholar] [CrossRef]
- Esterberg, R.; Linbo, T.; Pickett, S.B.; Wu, P.; Ou, H.C.; Rubel, E.W.; Raible, D.W. Mitochondrial Calcium Uptake Underlies ROS Generation during Aminoglycoside-Induced Hair Cell Death. J. Clin. Investig. 2016, 126, 3556–3566. [Google Scholar] [CrossRef]
- Huth, M.E.; Ricci, A.J.; Cheng, A.G. Mechanisms of Aminoglycoside Ototoxicity and Targets of Hair Cell Protection. Int. J. Otolaryngol. 2011, 2011, 937861. [Google Scholar] [CrossRef]
- Oishi, N.; Duscha, S.; Boukari, H.; Meyer, M.; Xie, J.; Wei, G.; Schrepfer, T.; Roschitzki, B.; Boettger, E.C.; Schacht, J. XBP1 Mitigates Aminoglycoside-Induced Endoplasmic Reticulum Stress and Neuronal Cell Death. Cell Death Dis. 2015, 6, e1763. [Google Scholar] [CrossRef] [PubMed]
- Esterberg, R.; Hailey, D.W.; Coffin, A.B.; Raible, D.W.; Rubel, E.W. Disruption of Intracellular Calcium Regulation Is Integral to Aminoglycoside-Induced Hair Cell Death. J. Neurosci. 2013, 33, 7513–7525. [Google Scholar] [CrossRef] [PubMed]
- Esterberg, R.; Hailey, D.W.; Rubel, E.W.; Raible, D.W. ER-Mitochondrial Calcium Flow Underlies Vulnerability of Mechanosensory Hair Cells to Damage. J. Neurosci. 2014, 34, 9703–9719. [Google Scholar] [CrossRef]
- Hobbie, S.N.; Bruell, C.M.; Akshay, S.; Kalapala, S.K.; Shcherbakov, D.; Böttger, E.C. Mitochondrial Deafness Alleles Confer Misreading of the Genetic Code. Biochemistry 2008, 105, 3244–3249. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Guan, M.-X. Interaction of Aminoglycosides with Human Mitochondrial 12S RRNA Carrying the Deafness-Associated Mutation. Antimicrob. Agents Chemother. 2009, 53, 4612–4618. [Google Scholar] [CrossRef]
- Matt, T.; Ng, C.L.; Lang, K.; Sha, S.-H.; Akbergenov, R.; Shcherbakov, D.; Meyer, M.; Duscha, S.; Xie, J.; Dubbaka, S.R.; et al. Dissociation of Antibacterial Activity and Aminoglycoside Ototoxicity in the 4-Monosubstituted 2-Deoxystreptamine Apramycin. Proc. Natl. Acad. Sci. USA 2012, 109, 10984–10989. [Google Scholar] [CrossRef]
- Campos, A.; López-Escámez, J.A.; Crespo, P.V.; Cañizares, F.J.; Baeyens, J.M. Gentamicin ototoxicity in otoconia: Quantitative electron probe X-ray microanalysis. Acta Otolaryngol. 1994, 114, 18–23. [Google Scholar] [CrossRef]
- López-Escámez, J.A.; Cañizares, F.J.; Crespo, P.V.; Baeyens, J.M.; Campos, A. Electron probe microanalysis of gentamicin-induced changes on ionic composition of the vestibular gelatinous membrane. Hear. Res. 1994, 76, 60–66. [Google Scholar] [CrossRef]
- Davies, J.; Davis, B.D. Misreading of Ribonucleic Acid Code Words Induced by Aminoglycoside Antibiotics. J. Biol. Chem. 1968, 243, 3312–3316. [Google Scholar] [CrossRef]
- Wimberly, B.T.; Brodersen, D.E.; Clemons, W.M.; Morgan-Warren, R.J.; Carter, A.P.; Vonrhein, C.; Hartsch, T.; Ramakrishnan, V. Structure of the 30S Ribosomal Subunit. Nature 2000, 407, 327–339. [Google Scholar] [CrossRef]
- Gutell, R.R.; Larsen, N.; Woese, C.R. Lessons from an Evolving RRNA: 16S and 23S RRNA Structures from a Comparative Perspective. Microbiol. Rev. 1994, 58, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Huth, M.E.; Han, K.-H.; Sotoudeh, K.; Hsieh, Y.-J.; Effertz, T.; Vu, A.A.; Verhoeven, S.; Hsieh, M.H.; Greenhouse, R.; Cheng, A.G.; et al. Designer Aminoglycosides Prevent Cochlear Hair Cell Loss and Hearing Loss. J. Clin. Investig. 2015, 125, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Pesini, E.; Wallace, D.C. Evidence for Adaptive Selection Acting on the TRNA and RRNA Genes of Human Mitochondrial DNA. Hum. Mutat. 2006, 27, 1072–1081. [Google Scholar] [CrossRef]
- Prezant, T.R.; Agapian, J.V.; Bohlman, M.C.; Bu, X.; Öztas, S.; Qiu, W.-Q.; Arnos, K.S.; Cortopassi, G.A.; Jaber, L.; Rotter, J.I.; et al. Mitochondrial Ribosomal RNA Mutation Associated with Both Antibiotic–Induced and Non–Syndromic Deafness. Nat. Genet. 1993, 4, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Bottger, E.C.; Qian, Y.; Guan, M.-X. Mutant A1555G Mitochondrial 12S RRNA and Aminoglycoside Susceptibility. Antimicrob. Agents Chemother. 2010, 54, 3073–3075. [Google Scholar] [CrossRef]
- Guan, M. Biochemical Evidence for Nuclear Gene Involvement in Phenotype of Non- Syndromic Deafness Associated with Mitochondrial 12S RRNA Mutation. Hum. Mol. Genet. 1996, 5, 963–971. [Google Scholar] [CrossRef]
- Nguyen, T.; Jeyakumar, A. Genetic Susceptibility to Aminoglycoside Ototoxicity. Int. J. Pediatr. Otorhinolaryngol. 2019, 120, 15–19. [Google Scholar] [CrossRef]
- Bitner-Glindzicz, M.; Pembrey, M.; Duncan, A.B.; Heron, J.; Ring, S.M.; Hall, A.M.; Rahman, S. Prevalence of Mitochondrial 1555A→G Mutation in European Children. N. Engl. J. Med. 2009, 360, 640–642. [Google Scholar] [CrossRef]
- Johnson, R.F.; Cohen, A.P.; Guo, Y.; Schibler, K.; Greinwald, J.H. Genetic Mutations and Aminoglycoside-Induced Ototoxicity in Neonates. Otolaryngol.–Head Neck Surg. 2010, 142, 704–707. [Google Scholar] [CrossRef]
- Lu, J.; Li, Z.; Zhu, Y.; Yang, A.; Li, R.; Zheng, J.; Cai, Q.; Peng, G.; Zheng, W.; Tang, X.; et al. Mitochondrial 12S RRNA Variants in 1642 Han Chinese Pediatric Subjects with Aminoglycoside-Induced and Nonsyndromic Hearing Loss. Mitochondrion 2010, 10, 380–390. [Google Scholar] [CrossRef]
- Usami, S.; Abe, S.; Akita, J.; Namba, A.; Shinkawa, H.; Ishii, M.; Iwasaki, S.; Hoshino, T.; Ito, J.; Doi, K.; et al. Prevalence of Mitochondrial Gene Mutations among Hearing Impaired Patients. J. Med. Genet. 2000, 37, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, Y.; Yashima, T.; Ito, T.; Sumi, T.; Tsuzuku, T.; Kitamura, K. Audiovestibular Findings in Patients with Mitochondrial A1555G Mutation. Laryngoscope 2004, 114, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Hutchin, T.; Haworth, I.S.; Higashi, K.; Fischel-Ghodsian, N.; Stoneking, M.; Saha, N.; Arnos, C.; Cortopassi, G.A. A Molecular Basis for Human Hypersensitivity of Aminoglyscoside Antibiotics. Nucleic Acids Res. 1993, 21, 4174–4179. [Google Scholar] [CrossRef] [PubMed]
- Fischel-Ghodsian, N.; Prezant, T.R.; Bu, X.; Öztas, S. Mitochondrial Ribosomal RNA Gene Mutation in a Patient with Sporadic Aminoglycoside Ototoxicity. Am. J. Otolaryngol. 1993, 14, 399–403. [Google Scholar] [CrossRef]
- Estivill, X.; Govea, N.; Barceló, A.; Perelló, E.; Badenas, C.; Romero, E.; Moral, L.; Scozzari, R.; D’Urbano, L.; Zeviani, M.; et al. Familial Progressive Sensorineural Deafness Is Mainly due to the MtDNA A1555G Mutation and Is Enhanced by Treatment with Aminoglycosides. Am. J. Hum. Genet. 1998, 62, 27–35. [Google Scholar] [CrossRef]
- Zhao, H.; Li, R.; Wang, Q.; Yan, Q.; Deng, J.-H.; Han, D.; Bai, Y.; Young, W.-Y.; Guan, M.-X. Maternally Inherited Aminoglycoside-Induced and Nonsyndromic Deafness Is Associated with the Novel C1494T Mutation in the Mitochondrial 12S RRNA Gene in a Large Chinese Family. Am. J. Hum. Genet. 2004, 74, 139–152. [Google Scholar] [CrossRef]
- Zhao, L.; Young, W.-Y.; Li, R.; Wang, Q.; Qian, Y.; Guan, M.-X. Clinical Evaluation and Sequence Analysis of the Complete Mitochondrial Genome of Three Chinese Patients with Hearing Impairment Associated with the 12S RRNA T1095C Mutation. Biochem. Biophys. Res. Commun. 2004, 325, 1503–1508. [Google Scholar] [CrossRef]
- Thyagarajan, D.; Bressman, S.; Bruno, C.; Przedborski, S.; Shanske, S.; Lynch, T.; Fahn, S.; DiMauro, S. A Novel Mitochondrial 12SrRNA Point Mutation in Parkinsonism, Deafness, and Neuropathy. Ann. Neurol. 2000, 48, 730–736. [Google Scholar] [CrossRef]
- Guan, M.X.; Fischel-Ghodsian, N.; Attardi, G. Nuclear Background Determines Biochemical Phenotype in the Deafness-Associated Mitochondrial 12S RRNA Mutation. Hum. Mol. Genet. 2001, 10, 573–580. [Google Scholar] [CrossRef]
- Li, X.; Guan, M.-X. A Human Mitochondrial GTP Binding Protein Related to TRNA Modification May Modulate Phenotypic Expression of the Deafness-Associated Mitochondrial 12S RRNA Mutation. Mol. Cell. Biol. 2002, 22, 7701–7711. [Google Scholar] [CrossRef]
- Bykhovskaya, Y.; Mengesha, E.; Wang, D.; Yang, H.; Estivill, X.; Shohat, M.; Fischel-Ghodsian, N. Phenotype of Non-Syndromic Deafness Associated with the Mitochondrial A1555G Mutation Is Modulated by Mitochondrial RNA Modifying Enzymes MTO1 and GTPBP3. Mol. Genet. Metab. 2004, 83, 199–206. [Google Scholar] [CrossRef]
- Chen, G.; He, F.; Fu, S.; Dong, J. GJB2 and Mitochondrial DNA 1555A > G Mutations in Students with Hearing Loss in the Hubei Province of China. Int. J. Pediatr. Otorhinolaryngol. 2011, 75, 1156–1159. [Google Scholar] [CrossRef]
- Wrześniok, D.; Beberok, A.; Otręba, M.; Buszman, E. Modulation of Melanogenesis and Antioxidant Defense System in Melanocytes by Amikacin. Toxicol. Vitr. 2013, 27, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.X.; Fischel-Ghodsian, N.; Attardi, G. A Biochemical Basis for the Inherited Susceptibility to Aminoglycoside Ototoxicity. Hum. Mol. Genet. 2000, 9, 1787–1793. [Google Scholar] [CrossRef]
- Ding, D.; Jiang, H.; Salvi, R.J. Mechanisms of Rapid Sensory Hair-Cell Death Following Co-Administration of Gentamicin and Ethacrynic Acid. Hear. Res. 2010, 259, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, H.; Johnson, A.; Karasawa, T.; Zhang, Y.; Meier, W.B.; Taghizadeh, F.; Kachelmeier, A.; Steyger, P.S. Inflammation Up-Regulates Cochlear Expression of TRPV1 to Potentiate Drug-Induced Hearing Loss. Sci. Adv. 2019, 5, aaw1836. [Google Scholar] [CrossRef]
- Li, Y.; Adamek, P.; Zhang, H.; Tatsui, C.E.; Rhines, L.D.; Mrozkova, P.; Li, Q.; Kosturakis, A.K.; Cassidy, R.M.; Harrison, D.J.; et al. The Cancer Chemotherapeutic Paclitaxel Increases Human and Rodent Sensory Neuron Responses to TRPV1 by Activation of TLR4. J. Neurosci. 2015, 35, 13487–13500. [Google Scholar] [CrossRef] [PubMed]
- Cross, C.P.; Liao, S.; Urdang, Z.D.; Srikanth, P.; Garinis, A.C.; Steyger, P.S. Effect of Sepsis and Systemic Inflammatory Response Syndrome on Neonatal Hearing Screening Outcomes Following Gentamicin Exposure. Int. J. Pediatr. Otorhinolaryngol. 2015, 79, 1915–1919. [Google Scholar] [CrossRef]
- Mukherjea, D.; Jajoo, S.; Whitworth, C.; Bunch, J.R.; Turner, J.G.; Rybak, L.P.; Ramkumar, V. Short Interfering RNA against Transient Receptor Potential Vanilloid 1 Attenuates Cisplatin-Induced Hearing Loss in the Rat. J. Neurosci. 2008, 28, 13056–13065. [Google Scholar] [CrossRef]
- Borse, V.; Al Aameri, R.F.H.; Sheehan, K.; Sheth, S.; Kaur, T.; Mukherjea, D.; Tupal, S.; Lowy, M.; Ghosh, S.; Dhukhwa, A.; et al. Epigallocatechin-3-Gallate, a Prototypic Chemopreventative Agent for Protection against Cisplatin-Based Ototoxicity. Cell Death Dis. 2017, 8, e2921. [Google Scholar] [CrossRef]
- Al-Malky, G.; Suri, R.; Dawson, S.J.; Sirimanna, T.; Kemp, D. Aminoglycoside Antibiotics Cochleotoxicity in Paediatric Cystic Fibrosis (CF) Patients: A Study Using Extended High-Frequency Audiometry and Distortion Product Otoacoustic Emissions. Int. J. Audiol. 2011, 50, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The Capsaicin Receptor: A Heat-Activated Ion Channel in the Pain Pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; He, W.; Yang, W.; Hetrick, A.P.; Gonzalez, J.G.; Sargsyan, L.; Wu, H.; Jung, T.T.K.; Li, H. Intratympanic Lipopolysaccharide Elevates Systemic Fluorescent Gentamicin Uptake in the Cochlea. Laryngoscope 2021, 131, E2573–E2582. [Google Scholar] [CrossRef]
- Zager, R.A. Endotoxemia, Renal Hypoperfusion, and Fever: Interactive Risk Factors for Aminoglycoside and Sepsis-Associated Acute Renal Failure. Am. J. Kidney Dis. 1992, 20, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.L.; Strologo, L.D.; Bianchi, P.M.; Tieri, L.; Rizzoni, G. Sensorineural Hearing Loss in Patients Reaching Chronic Renal Failure in Childhood. Pediatr. Nephrol. 1996, 10, 38–40. [Google Scholar] [CrossRef]
- Gatland, D.; Tucker, B.; Chalstrey, S.; Keene, M.; Baker, L. Hearing Loss in Chronic Renal Failure-Hearing Threshold Changes Following Haemodialysis. J. R. Soc. Med. 1991, 84, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Thodi, C.; Thodis, E.; Danielides, V.; Pasadakis, P.; Vargemezis, V. Hearing in Renal Failure. Nephrol. Dial. Transplant. 2006, 21, 3023–3030. [Google Scholar] [CrossRef]
- Vilayur, E.; Gopinath, B.; Harris, D.C.; Burlutsky, G.; McMahon, C.M.; Mitchell, P. The Association between Reduced GFR and Hearing Loss: A Cross-Sectional Population-Based Study. Am. J. Kidney Dis. 2010, 56, 661–669. [Google Scholar] [CrossRef]
- Adler, D.; Fiehn, W.; Ritz, E. Inhibition of Na+, K+-Stimulated ATPase in the Cochlea of the Guinea PIG: A Potential Cause of Disturbed Inner Ear Function in Terminal Renal Failirre. Acta Oto-Laryngol. 1980, 90, 55–60. [Google Scholar] [CrossRef]
- Ohashi, T.; Kenmochi, M.; Kinoshita, H.; Ochi, K.; Kikuchi, H. Cochlear Function of Guinea Pigs with Experimental Chronic Renal Failure. Ann. Otol. Rhinol. Laryngol. 1999, 108, 955–962. [Google Scholar] [CrossRef]
- Di Paolo, B.; Di Marco, T.; Cappelli, P.; Spisni, C.; Del Rosso, G.; Palmieri, P.F.; Evangelista, M.; Albertazzi, A. Electrophysiological Aspects of Nervous Conduction in Uremia. Clin. Nephrol. 1988, 29, 253–260. [Google Scholar] [PubMed]
- Lautermann, J.; McLaren, J.; Schacht, J. Glutathione Protection against Gentamicin Ototoxicity Depends on Nutritional Status. Hear. Res. 1995, 86, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-D.; Kao, M.-C.; Tsai, M.-H.; Lai, C.-H.; Wei, I.-H.; Tsai, M.-H.; Tang, C.-H.; Lin, C.-W.; Hsu, C.-J.; Lin, C.-Y. Transient Ischemia/Hypoxia Enhances Gentamicin Ototoxicity via Caspase-Dependent Cell Death Pathway. Lab. Investig. 2011, 91, 1092–1106. [Google Scholar] [CrossRef]
- Brummett, R.E.; Fox, K.E.; Jacobs, F.; Kempton, J.B.; Stokes, Z.; Allyson, B. Augmented Gentamicin Ototoxicity Induced by Vancomycin in Guinea Pigs. Arch. Otolaryngol.-Head Neck Surg. 1990, 116, 61–64. [Google Scholar] [CrossRef]
- Rybak, L.P. Ototoxicity of Loop Diuretics. Otolaryngol. Clin. N. Am. 1993, 26, 829–844. [Google Scholar] [CrossRef]
- Bates, D.E.; Beaumont, S.J.; Baylis, B.W. Ototoxicity Induced by Gentamicin and Furosemide. Ann. Pharmacother. 2002, 36, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Capozza, M.A.; Mastrangelo, S.; Maurizi, P.; Triarico, S.; Rolesi, R.; Attinà, G.; Fetoni, A.R.; Ruggiero, A. Assessment and Management of Platinum-Related Ototoxicity in Children Treated for Cancer. Cancers 2020, 12, 1266. [Google Scholar] [CrossRef]
- Romano, A.; Rivetti, S.; Brigato, F.; Mastrangelo, S.; Attinà, G.; Maurizi, P.; Galli, J.; Fetoni, A.R.; Ruggiero, A. Early and Long-Term Ototoxicity Noted in Children due to Platinum Compounds: Prevalence and Risk Factors. Biomedicines 2023, 11, 261. [Google Scholar] [CrossRef]
- Riggs, L.C.; Brummett, R.E.; Guitjens, S.K.; Matz, G.J. Ototoxicity Resulting from Combined Administration of Cisplatin and Gentamicin. Laryngoscope 1996, 106, 401–406. [Google Scholar] [CrossRef]
- Li, H.; Kachelmeier, A.; Furness, D.N.; Steyger, P.S. Local Mechanisms for Loud Sound-Enhanced Aminoglycoside Entry into Outer Hair Cells. Front. Cell. Neurosci. 2015, 9, 130. [Google Scholar] [CrossRef]
- Wu, F.; Xiong, H.; Sha, S. Noise-Induced Loss of Sensory Hair Cells Is Mediated by ROS/AMPKα Pathway. Redox Biol. 2020, 29, 101406. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.W.P. Synergistic Interactions of Gentamicin and Pure Tones Causing Cochlear Hair Cell Loss in Pigmented Guinea Pigs. Hear. Res. 1988, 36, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.F.; Bone, R.C. Potentiation of Kanamycin Ototoxicity by a History of Noise Exposure. Otolaryngology 1978, 86, ORL-125–ORL-128. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.F.; Bone, R.G. Non-Simultaneous Interaction of Exposure to Noise and Kanamycin Intoxication in the Chinchilla. Am. J. Otolaryngol. 1982, 3, 264–272. [Google Scholar] [CrossRef]
- McKinney, W.; Yonovitz, A.; Smolensky, M.H. Circadian Variation of Gentamicin Toxicity in Rats. Laryngoscope 2015, 125, E252–E256. [Google Scholar] [CrossRef]
- Blunston, M.A.; Yonovitz, A.; Woodahl, E.L.; Smolensky, M.H. Gentamicin-Induced Ototoxicity and Nephrotoxicity Vary with Circadian Time of Treatment and Entail Separate Mechanisms. Chronobiol. Int. 2015, 32, 1223–1232. [Google Scholar] [CrossRef]
- Martins, L.M.N.; Camargos, P.A.M.; Becker, H.M.G.; Becker, C.G.; Guimarães, R.E.S. Hearing Loss in Cystic Fibrosis. Int. J. Pediatr. Otorhinolaryngol. 2010, 74, 469–473. [Google Scholar] [CrossRef]
- Mulheran, M.; Degg, C.; Burr, S.; Morgan, D.W.; Stableforth, D.E. Occurrence and Risk of Cochleotoxicity in Cystic Fibrosis Patients Receiving Repeated High-Dose Aminoglycoside Therapy. Antimicrob. Agents Chemother. 2001, 45, 2502–2509. [Google Scholar] [CrossRef]
- Garinis, A.C.; Cross, C.P.; Srikanth, P.; Carroll, K.; Feeney, M.P.; Keefe, D.H.; Hunter, L.L.; Putterman, D.B.; Cohen, D.M.; Gold, J.A.; et al. The Cumulative Effects of Intravenous Antibiotic Treatments on Hearing in Patients with Cystic Fibrosis. J. Cyst. Fibros. 2017, 16, 401–409. [Google Scholar] [CrossRef]
- Piltcher, O.B.; Teixeira, V.N.; de Oliveira, M.W.; Scattolin, I.; Piltcher, S.L. The Prevalence of Neurosensorial Hearing Loss among Cystic Fibrosis Patients from Hospital de Clínicas de Porto Alegre. Int. J. Pediatr. Otorhinolaryngol. 2003, 67, 939–941. [Google Scholar] [CrossRef]
- Tarshish, Y.; Huang, L.; Jackson, F.I.; Edwards, J.; Fligor, B.; Wilkins, A.; Uluer, A.; Sawicki, G.; Kenna, M. Risk Factors for Hearing Loss in Patients with Cystic Fibrosis. J. Am. Acad. Audiol. 2016, 27, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Geyer, L.B.; Menna Barreto, S.S.; Weigert, L.L.; Teixeira, A.R. High Frequency Hearing Thresholds and Product Distortion Otoacoustic Emissions in Cystic Fibrosis Patients. Braz. J. Otorhinolaryngol. 2015, 81, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Mulheran, M.; Hyman-Taylor, P.; Tan, K.H.-V.; Lewis, S.; Stableforth, D.; Knox, A.; Smyth, A. Absence of Cochleotoxicity Measured by Standard and High-Frequency Pure Tone Audiometry in a Trial of Once- versus Three-Times-Daily Tobramycin in Cystic Fibrosis Patients. Antimicrob. Agents Chemother. 2006, 50, 2293–2299. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, J.; Jahnke, N.; Smyth, A.R. Once-Daily versus Multiple-Daily Dosing with Intravenous Aminoglycosides for Cystic Fibrosis. Cochrane Database Syst. Rev. 2019, 9, CD002009. [Google Scholar] [CrossRef] [PubMed]
- Cotanche, D.A. Genetic and Pharmacological Intervention for Treatment/Prevention of Hearing Loss. J. Commun. Disord. 2008, 41, 421–443. [Google Scholar] [CrossRef]
- Pirvola, U.; Xing-Qun, L.; Virkkala, J.; Saarma, M.; Murakata, C.; Camoratto, A.M.; Walton, K.M.; Ylikoski, J. Rescue of Hearing, Auditory Hair Cells, and Neurons by CEP-1347/KT7515, an Inhibitor of C-Jun N-Terminal Kinase Activation. J. Neurosci. 2000, 20, 43–50. [Google Scholar] [CrossRef]
- Wang, J.; Van De Water, T.R.; Bonny, C.; de Ribaupierre, F.; Puel, J.L.; Zine, A. A Peptide Inhibitor of C-Jun N-Terminal Kinase Protects against Both Aminoglycoside and Acoustic Trauma-Induced Auditory Hair Cell Death and Hearing Loss. J. Neurosci. 2003, 23, 8596–8607. [Google Scholar] [CrossRef]
- Conlon, B.J.; Aran, J.-M.; Erre, J.-P.; Smith, D.W. Attenuation of Aminoglycoside-Induced Cochlear Damage with the Metabolic Antioxidant α-Lipoic Acid. Hear. Res. 1999, 128, 40–44. [Google Scholar] [CrossRef]
- Song, B.-B.; Sha, S.-H.; Schacht, J. Iron Chelators Protect from Aminoglycoside-Induced Cochleo- and Vestibulo-Toxicity. Free Radic. Biol. Med. 1998, 25, 189–195. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, W.-G.; Zha, D.-J.; Qiu, J.-H.; Wang, J.-L.; Sha, S.-H.; Schacht, J. Aspirin Attenuates Gentamicin Ototoxicity: From the Laboratory to the Clinic. Hear. Res. 2007, 226, 178–182. [Google Scholar] [CrossRef]
- Jiang, H.; Sha, S.-H.; Schacht, J. NF-κB Pathway Protects Cochlear Hair Cells from Aminoglycoside-Induced Ototoxicity. J. Neurosci. Res. 2005, 79, 644–651. [Google Scholar] [CrossRef]
- Ramakrishnan, R.; Jusko, W.J. Interactions of Aspirin and Salicylic Acid with Prednisolone for Inhibition of Lymphocyte Proliferation. Int. Immunopharmacol. 2001, 1, 2035–2042. [Google Scholar] [CrossRef] [PubMed]
- Sha, S.-H.; Schacht, J. Antioxidants Attenuate Gentamicin-Induced Free Radical Formation in Vitro and Ototoxicity in Vivo: D-Methionine Is a Potential Protectant. Hear. Res. 2000, 142, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.C.M.; Meech, R.P.; Klemens, J.J.; Gerberi, M.T.; Dyrstad, S.S.W.; Larsen, D.L.; Mitchell, D.L.; El-Azizi, M.; Verhulst, S.J.; Hughes, L.F. Prevention of Noise- and Drug-Induced Hearing Loss with D-Methionine. Hear. Res. 2007, 226, 92–103. [Google Scholar] [CrossRef]
- Fox, D.J.; Cooper, M.D.; Speil, C.A.; Roberts, M.H.; Yanik, S.C.; Meech, R.P.; Hargrove, T.L.; Verhulst, S.J.; Rybak, L.P.; Campbell, K.C.M. D-Methionine Reduces Tobramycin-Induced Ototoxicity without Antimicrobial Interference in Animal Models. J. Cyst. Fibros. 2016, 15, 518–530. [Google Scholar] [CrossRef]
- Rybak, L.P.; Whitworth, C.A. Ototoxicity: Therapeutic Opportunities. Drug Discov. Today 2005, 10, 1313–1321. [Google Scholar] [CrossRef]
- Noack, V.; Pak, K.; Jalota, R.; Kurabi, A.; Ryan, A.F. An Antioxidant Screen Identifies Candidates for Protection of Cochlear Hair Cells from Gentamicin Toxicity. Front. Cell. Neurosci. 2017, 11, 242. [Google Scholar] [CrossRef] [PubMed]
- Kranzer, K.; Elamin, W.F.; Cox, H.; Seddon, J.A.; Ford, N.; Drobniewski, F. A Systematic Review and Meta-Analysis of the Efficacy and Safety of N-Acetylcysteine in Preventing Aminoglycoside-Induced Ototoxicity: Implications for the Treatment of Multidrug-Resistant TB. Thorax 2015, 70, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Vural, A.; Koçyiğit, İ.; Şan, F.; Eroğlu, E.; Ketenci, İ.; Ünal, A.; Tokgöz, B.; Ünlü, Y. Long-Term Protective Effect of N-Acetylcysteine against Amikacin-Induced Ototoxicity in End-Stage Renal Disease: A Randomized Trial. Perit. Dial. Int. 2017, 38, 57–62. [Google Scholar] [CrossRef]
- Parry, M.F.; Neu, H.C. Effect of N-Acetylcysteine on Antibiotic Activity and Bacterial Growth in Vitro. J. Clin. Microbiol. 1977, 5, 58–61. [Google Scholar] [CrossRef]
- Forcheron, F.; Cachefo, A.; Thevenon, S.; Pinteur, C.; Beylot, M. Mechanisms of the Triglyceride- and Cholesterol-Lowering Effect of Fenofibrate in Hyperlipidemic Type 2 Diabetic Patients. Diabetes 2002, 51, 3486–3491. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Ji, H.M.; Kim, S.J.; Kil, S.H.; Lee, J.N.; Kwak, S.; Choe, S.K.; Park, R. Fenofibrate Exerts Protective Effects against Gentamicin-Induced Toxicity in Cochlear Hair Cells by Activating Antioxidant Enzymes. Int. J. Mol. Med. 2017, 39, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Broderick, M.T.; Prince, A.D.P.; Dhukhwa, A.; Mukherjea, D.; Jiang, P.; Campbell, K.C.M.; Rybak, L.P.; Brenner, M.J. Effects of Moringa Extract on Aminoglycoside-Induced Hair Cell Death and Organ of Corti Damage. Otol. Neurotol. 2021, 42, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, N.K.; O’Reilly, M.; Derudas, M.; Kenyon, E.J.; Huckvale, R.; van Netten, S.M.; Ward, S.E.; Richardson, G.P.; Kros, C.J. D-Tubocurarine and Berbamine: Alkaloids That Are Permeant Blockers of the Hair Cell’s Mechano-Electrical Transducer Channel and Protect from Aminoglycoside Toxicity. Front. Cell. Neurosci. 2017, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Vélez-Ortega, A.C.; Freeman, M.J.; Indzhykulian, A.A.; Grossheim, J.M.; Frolenkov, G.I. Mechanotransduction Current Is Essential for Stability of the Transducing Stereocilia in Mammalian Auditory Hair Cells. eLife 2017, 6, e24661. [Google Scholar] [CrossRef]
- Vu, A.A.; Nadaraja, G.S.; Huth, M.E.; Luk, L.; Kim, J.; Chai, R.; Ricci, A.J.; Cheng, A.G. Integrity and Regeneration of Mechanotransduction Machinery Regulate Aminoglycoside Entry and Sensory Cell Death. PLoS ONE 2013, 8, e54794. [Google Scholar] [CrossRef]
- Coffin, A.B.; Reinhart, K.E.; Owens, K.N.; Raible, D.W.; Rubel, E.W. Extracellular Divalent Cations Modulate Aminoglycoside-Induced Hair Cell Death in the Zebrafish Lateral Line. Hear. Res. 2009, 253, 42–51. [Google Scholar] [CrossRef]
- Kim, S.; Jung, G.; Kim, S.-J.; Koo, J.S. Novel Peptide Vaccine GV1001 Rescues Hearing in Kanamycin/Furosemide-Treated Mice. Front. Cell. Neurosci. 2018, 12, 3. [Google Scholar] [CrossRef]
- Christensen, E.; Reiffenstein, J.C.; Madissoo, H. Comparative Ototoxicity of Amikacin and Gentamicin in Cats. Antimicrob. Agents Chemother. 1977, 12, 178–184. [Google Scholar] [CrossRef]
- Shao, W.; Zhong, D.; Jiang, H.; Han, Y.; Cui, Q.; Ruining, L.; Qian, X.; Chen, D.; Jing, L. A New Aminoglycoside Etimicin Shows Low Nephrotoxicity and Ototoxicity in Zebrafish Embryos. J. Appl. Toxicol. 2020, 41, 1063–1075. [Google Scholar] [CrossRef]
- Ferriols-Lisart, R.; Alos-Alminana, M. Effectiveness and Safety of Once-Daily Aminoglycosides: A Meta-Analysis. Am. J. Health-Syst. Pharm. 1996, 53, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Hatala, R. Once-Daily Aminoglycoside Dosing in Immunocompetent Adults. Ann. Intern. Med. 1996, 124, 717. [Google Scholar] [CrossRef] [PubMed]
- Barza, M.; Ioannidis, J.P.; Cappelleri, J.C.; Lau, J. Single or Multiple Daily Doses of Aminoglycosides: A Meta-Analysis. BMJ 1996, 312, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.C.; Little, J.R.; Littenberg, B.; Reichley, R.M.; Dunagan, W.C. A Meta-Analysis of Extended-Interval Dosing versus Multiple Daily Dosing of Aminoglycosides. Clin. Infect. Dis. 1997, 24, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Best, E.J.; Gazarian, M.; Cohn, R.; Wilkinson, M.; Palasanthiran, P. Once-Daily Gentamicin in Infants and Children: A Prospective Cohort Study Evaluating Safety and the Role of Therapeutic Drug Monitoring in Minimizing Toxicity. Pediatr. Infect. Dis. J. 2011, 30, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Freeman, C.D.; Nicolau, D.P.; Belliveau, P.P.; Nightingale, C.H. Once-Daily Dosing of Aminoglycosides: Review and Recommendations for Clinical Practice. J. Antimicrob. Chemother. 1997, 39, 677–686. [Google Scholar] [CrossRef] [PubMed]


| Aminoglycoside | Cochleotoxicity a | Vestibulotoxicity b |
|---|---|---|
| Amikacin | yes | Not toxic |
| Gentamicin | minor | yes |
| Kanamycin | yes | minor |
| Netilmicin | yes | yes |
| Neomycin | very toxic | minor |
| Streptomicin | minor | very toxic |
| Tobramycin | yes | yes |
| Uncontrollable | Controllable |
|---|---|
| Genetic susceptibility * | Sepsis |
| Cystic fibrosis (multiple exposures) Neonatal age (high Vd, renal immaturity with low glomerular filtration rate and slow elimination) | Otitis media |
| Renal failure (acute or chronic kidney disease) | |
| Depletion of endogenous antioxidant (e.g., glutathione) | |
| Cochlea ischemia | |
| Concomitant ototoxic drugs (e.g., loop diuretics, vancomycin, platinum compounds) | |
| Loud sound exposure | |
| Administration during rest time |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivetti, S.; Romano, A.; Mastrangelo, S.; Attinà, G.; Maurizi, P.; Ruggiero, A. Aminoglycosides-Related Ototoxicity: Mechanisms, Risk Factors, and Prevention in Pediatric Patients. Pharmaceuticals 2023, 16, 1353. https://doi.org/10.3390/ph16101353
Rivetti S, Romano A, Mastrangelo S, Attinà G, Maurizi P, Ruggiero A. Aminoglycosides-Related Ototoxicity: Mechanisms, Risk Factors, and Prevention in Pediatric Patients. Pharmaceuticals. 2023; 16(10):1353. https://doi.org/10.3390/ph16101353
Chicago/Turabian StyleRivetti, Serena, Alberto Romano, Stefano Mastrangelo, Giorgio Attinà, Palma Maurizi, and Antonio Ruggiero. 2023. "Aminoglycosides-Related Ototoxicity: Mechanisms, Risk Factors, and Prevention in Pediatric Patients" Pharmaceuticals 16, no. 10: 1353. https://doi.org/10.3390/ph16101353