
Absence of Association between Methylene Blue Reduced Susceptibility and Polymorphisms in 12 Genes Involved in Antimalarial Drug Resistance in African Plasmodium falciparum

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Pfcrt
2.2. Pfdhfr
2.3. Pfmdr1
2.4. Pfmdr5
2.5. Pfmdr6
2.6. PfK13
2.7. Pfubq
2.8. Pfcarl
2.9. Pfugt
2.10. Pfact
2.11. Pfcoronin
2.12. Pfpm2
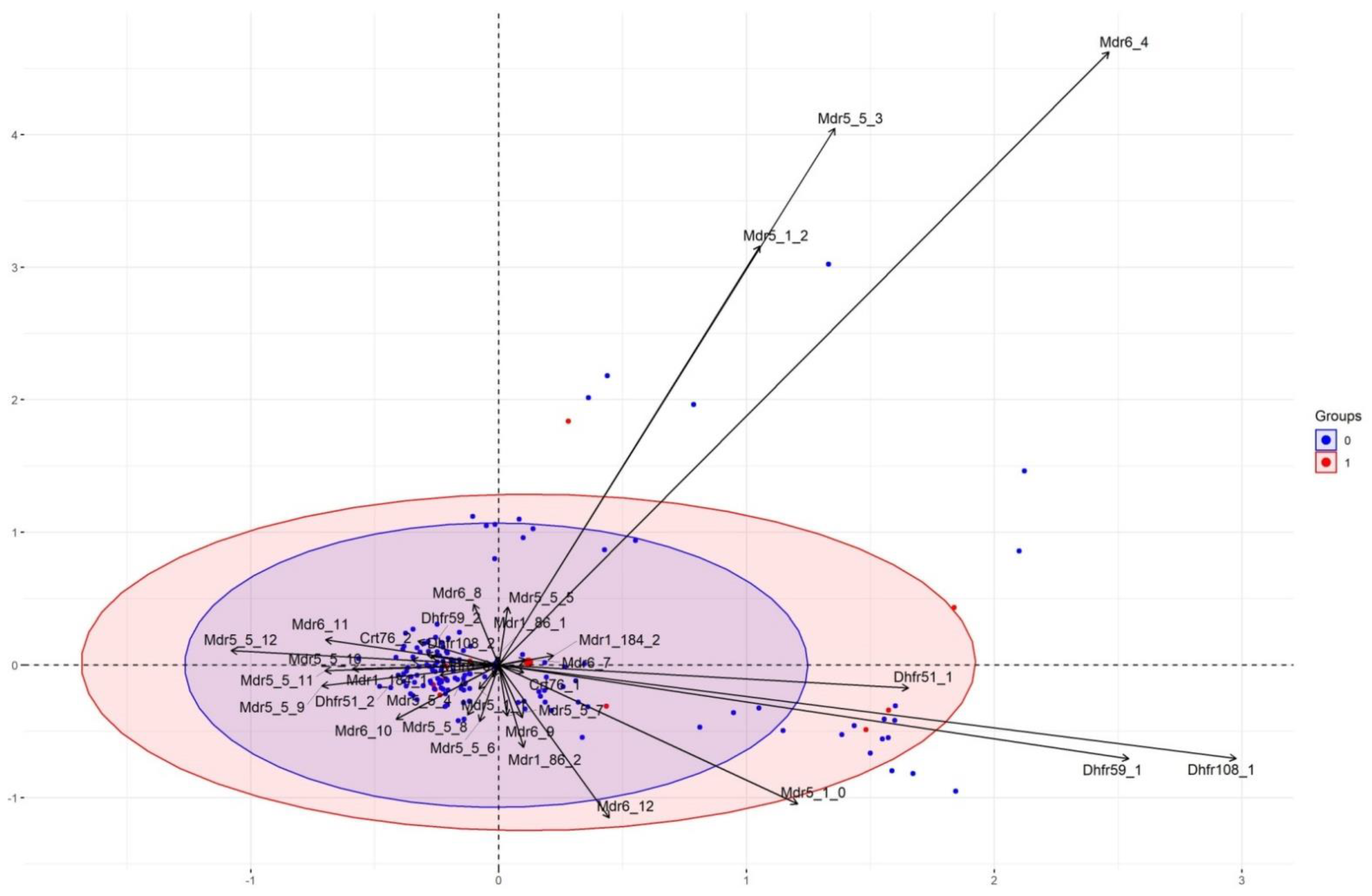
2.13. Multiple Correspondence Analysis (MCA)
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Ex Vivo Assay
4.3. Gene Sequencing
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Malaria Report 2020. Available online: https://www.who.int/publications-detail-redirect/9789240015791 (accessed on 6 January 2021).
- Gendrot, M.; Fawaz, R.; Dormoi, J.; Madamet, M.; Pradines, B. Genetic diversity and deletion of Plasmodium falciparum Histidine-Rich Protein 2 and 3: A Threat to diagnosis of P. falciparum malaria. Clin. Microbiol. Infect. 2019, 25, 580–585. [Google Scholar] [CrossRef]
- World Malaria Report 2019. Available online: http://www.who.int/malaria/publications/world-malaria-report-2019/en/ (accessed on 6 January 2021).
- World Health Organization WHO Status Report on Artemisinin and ACT Resistance (April 2017). Available online: http://www.who.int/malaria/publications/atoz/artemisinin-resistance-april2017/en/ (accessed on 6 January 2021).
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Ashley, E.A.; Dhorda, M.; Fairhurst, R.M.; Amaratunga, C.; Lim, P.; Suon, S.; Sreng, S.; Anderson, J.M.; Mao, S.; Sam, B.; et al. Spread of artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 2014, 371, 411–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akoachere, M.; Buchholz, K.; Fischer, E.; Burhenne, J.; Haefeli, W.E.; Schirmer, R.H.; Becker, K. In vitro assessment of methylene blue on chloroquine-sensitive and -resistant Plasmodium falciparum strains reveals synergistic action with artemisinins. Antimicrob. Agents Chemother. 2005, 49, 4592–4597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garavito, G.; Bertani, S.; Rincon, J.; Maurel, S.; Monje, M.C.; Landau, I.; Valentin, A.; Deharo, E. Blood schizontocidal activity of methylene blue in combination with antimalarials against Plasmodium falciparum. Parasite 2007, 14, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ademowo, O.G.; Nneji, C.M.; Adedapo, A.D.A. In vitro antimalarial activity of methylene blue against field isolates of Plasmodium falciparum from children in southwest Nigeria. Indian J. Med. Res. 2007, 126, 45–49. [Google Scholar] [PubMed]
- Pascual, A.; Henry, M.; Briolant, S.; Charras, S.; Baret, E.; Amalvict, R.; des Etages, E.H.; Feraud, M.; Rogier, C.; Pradines, B. In vitro activity of Proveblue (methylene blue) on Plasmodium falciparum strains resistant to standard antimalarial drugs. Antimicrob. Agents Chemother. 2011, 55, 2472–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fall, B.; Camara, C.; Fall, M.; Nakoulima, A.; Dionne, P.; Diatta, B.; Diemé, Y.; Wade, B.; Pradines, B. Plasmodium falciparum susceptibility to standard and potential anti-malarial drugs in Dakar, Senegal, during the 2013–2014 malaria season. Malar. J. 2015, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fall, B.; Madamet, M.; Diawara, S.; Briolant, S.; Wade, K.A.; Lo, G.; Nakoulima, A.; Fall, M.; Bercion, R.; Kounta, M.B.; et al. Ex vivo activity of Proveblue, a methylene blue, against field isolates of Plasmodium falciparum in Dakar, Senegal from 2013–2015. Int. J. Antimicrob. Agents 2017, 50, 155–158. [Google Scholar] [CrossRef]
- Gendrot, M.; Madamet, M.; Mosnier, J.; Fonta, I.; Amalvict, R.; Benoit, N.; Briolant, S.; Pradines, B. Baseline and multinormal distribution of ex vivo susceptibilities of Plasmodium falciparum to methylene blue in Africa, 2013-18. J. Antimicrob. Chemother. 2020, 75, 2141–2148. [Google Scholar] [CrossRef] [PubMed]
- Suwanarusk, R.; Russell, B.; Ong, A.; Sriprawat, K.; Chu, C.S.; PyaePhyo, A.; Malleret, B.; Nosten, F.; Renia, L. Methylene blue inhibits the asexual development of vivax malaria parasites from a region of increasing chloroquine resistance. J. Antimicrob. Chemother. 2015, 70, 124–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirjanata, G.; Sebayang, B.F.; Chalfein, F.; Prayoga; Handayuni, I.; Trianty, L.; Kenangalem, E.; Noviyanti, R.; Campo, B.; Poespoprodjo, J.R.; et al. Potent ex vivo activity of naphthoquine and methylene blue against drug-resistant clinical isolates of Plasmodium falciparum and Plasmodium vivax. Antimicrob. Agents Chemother. 2015, 59, 6117–6124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormoi, J.; Pascual, A.; Briolant, S.; Amalvict, R.; Charras, S.; Baret, E.; des Etages, E.H.; Feraud, M.; Pradines, B. Proveblue (methylene blue) as an antimalarial agent: In vitro synergy with dihydroartemisinin and atorvastatin. Antimicrob. Agents Chemother. 2012, 56, 3467–3469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormoi, J.; Pradines, B. Dose responses of Proveblue methylene blue in an experimental murine cerebral malaria model. Antimicrob. Agents Chemother. 2013, 57, 4080–4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormoi, J.; Briolant, S.; Desgrouas, C.; Pradines, B. Efficacy of Proveblue (methylene blue) in an experimental cerebral malaria murine model. Antimicrob. Agents Chemother. 2013, 57, 3412–3414. [Google Scholar] [CrossRef] [Green Version]
- Dormoi, J.; Briolant, S.; Desgrouas, C.; Pradines, B. Impact of methylene blue and atorvastatin combination therapy on the apparition of cerebral malaria in a murine model. Malar. J. 2013, 12, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adjalley, S.H.; Johnston, G.L.; Li, T.; Eastman, R.T.; Ekland, E.H.; Eappen, A.G.; Richman, A.; Sim, B.K.L.; Lee, M.C.S.; Hoffman, S.L.; et al. Quantitative assessment of plasmodium falciparum sexual development reveals potent transmission-blocking activity by methylene blue. Proc. Natl. Acad. Sci. USA 2011, 108, E1214–E1223. [Google Scholar] [CrossRef] [Green Version]
- Delves, M.J.; Ruecker, A.; Straschil, U.; Lelièvre, J.; Marques, S.; López-Barragán, M.J.; Herreros, E.; Sinden, R.E. Male and female Plasmodium falciparum mature gametocytes show different responses to antimalarial drugs. Antimicrob. Agents Chemother. 2013, 57, 3268–3274. [Google Scholar] [CrossRef] [Green Version]
- Wadi, I.; Pillai, C.R.; Anvikar, A.R.; Sinha, A.; Nath, M.; Valecha, N. Methylene blue induced morphological deformations in Plasmodium falciparum gametocytes: Implications for transmission-blocking. Malar. J. 2018, 17, 11. [Google Scholar] [CrossRef] [Green Version]
- Bosson-Vanga, H.; Franetich, J.-F.; Soulard, V.; Sossau, D.; Tefit, M.; Kane, B.; Vaillant, J.-C.; Borrmann, S.; Müller, O.; Dereuddre-Bosquet, N.; et al. Differential activity of methylene blue against erythrocytic and hepatic stages of Plasmodium. Malar. J. 2018, 17, 143. [Google Scholar] [CrossRef]
- Bradley, J.; Soumaré, H.M.; Mahamar, A.; Diawara, H.; Roh, M.; Delves, M.; Drakeley, C.; Churcher, T.S.; Dicko, A.; Gosling, R.; et al. Transmission-blocking effects of primaquine and methylene blue suggest Plasmodium falciparum gametocyte sterilization rather than effects on sex ratio. Clin. Infect. Dis. 2019, 69, 1436–1439. [Google Scholar] [CrossRef] [PubMed]
- Coulibaly, B.; Pritsch, M.; Bountogo, M.; Meissner, P.E.; Nebié, E.; Klose, C.; Kieser, M.; Berens-Riha, N.; Wieser, A.; Sirima, S.B.; et al. Efficacy and safety of triple combination therapy with artesunate-amodiaquine-methylene blue for falciparum malaria in children: A randomized controlled trial in Burkina Faso. J. Infect. Dis. 2015, 211, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicko, A.; Roh, M.E.; Diawara, H.; Mahamar, A.; Soumare, H.M.; Lanke, K.; Bradley, J.; Sanogo, K.; Kone, D.T.; Diarra, K.; et al. Efficacy and safety of primaquine and methylene blue for prevention of Plasmodium falciparum transmission in Mali: A phase 2, single-blind, randomised controlled trial. Lancet Infect. Dis. 2018, 18, 627–639. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, K.; Davioud-Charvet, E.; Ke, H.; Vaidya, A.B.; Lanzer, M.; Deponte, M. The Antimalarial activities of methylene blue and the 1,4-naphthoquinone 3-[4-(trifluoromethyl)benzyl]-menadione are not due to inhibition of the mitochondrial electron transport chain. Antimicrob. Agents Chemother. 2013, 57, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, O.; Davioud-Charvet, E.; Elhabiri, M. Interactions of the antimalarial drug methylene blue with methemoglobin and heme targets in Plasmodium falciparum: A physico-biochemical study. Antioxid. Redox Signal. 2012, 17, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Locher, K.P. Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat. Struct. Mol. Biol. 2016, 23, 487–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoulou, F.L.; Kerr, I.D. ABC transporter research: Going strong 40 years on. Biochem. Soc. Trans. 2015, 43, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Pradines, B.; Parquet, V.; Orlandi-Pradines, E. ABC transporters in Plasmodium falciparum and their involvement in resistance to antimalarial drugs. In ABC Transporters in Microorganisms: Research, Innovation and Value as Targets against Drug Resistance; Ponte-Sucre, A., Ed.; Caister Academic Press: Caister, UK, 2009; pp. 113–128. [Google Scholar]
- Wilson, C.M.; Serrano, A.E.; Wasley, A.; Bogenschutz, M.P.; Shankar, A.H.; Wirth, D.F. Amplification of a gene related to mammalian mdr genes in drug-resistant Plasmodium falciparum. Science 1989, 244, 1184–1186. [Google Scholar] [CrossRef]
- Foote, S.J.; Kyle, D.E.; Martin, R.K.; Oduola, A.M.; Forsyth, K.; Kemp, D.J.; Cowman, A.F. Several alleles of the multidrug-resistance gene are closely linked to chloroquine resistance in Plasmodium falciparum. Nature 1990, 345, 255–258. [Google Scholar] [CrossRef]
- Reed, M.B.; Saliba, K.J.; Caruana, S.R.; Kirk, K.; Cowman, A.F. Pgh1 modulates sensitivity and resistance to multiple antimalarials in Plasmodium falciparum. Nature 2000, 403, 906–909. [Google Scholar] [CrossRef]
- Duraisingh, M.T.; Jones, P.; Sambou, I.; von Seidlein, L.; Pinder, M.; Warhurst, D.C. The Tyrosine-86 allele of the Pfmdr1 gene of Plasmodium falciparum is associated with increased sensitivity to the anti-malarials mefloquine and artemisinin. Mol. Biochem. Parasitol. 2000, 108, 13–23. [Google Scholar] [CrossRef]
- Gendrot, M.; Wague Gueye, M.; Foguim, F.T.; Madamet, M.; Wade, K.A.; Bou Kounta, M.; Fall, M.; Diawara, S.; Benoit, N.; Lo, G.; et al. Modulation of in vitro antimalarial responses by polymorphisms in Plasmodium falciparum ABC transporters (Pfmdr1 and Pfmdr5). Acta Trop. 2019, 196, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, N.; Fall, B.; Pascual, A.; Fall, M.; Baret, E.; Camara, C.; Nakoulima, A.; Diatta, B.; Fall, K.B.; Mbaye, P.S.; et al. Role of Pfmdr1 in in vitro Plasmodium falciparum susceptibility to chloroquine, quinine, monodesethylamodiaquine, mefloquine, lumefantrine, and dihydroartemisinin. Antimicrob. Agents Chemother. 2014, 58, 7032–7040. [Google Scholar] [CrossRef] [Green Version]
- Kavishe, R.A.; van den Heuvel, J.M.W.; van de Vegte-Bolmer, M.; Luty, A.J.F.; Russel, F.G.M.; Koenderink, J.B. Localization of the ATP-binding cassette (ABC) transport proteins PfMRP1, PfMRP2, and PfMDR5 at the Plasmodium falciparum plasma membrane. Malar. J. 2009, 8, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okombo, J.; Abdi, A.I.; Kiara, S.M.; Mwai, L.; Pole, L.; Sutherland, C.J.; Nzila, A.; Ochola-Oyier, L.I. Repeat Polymorphisms in the low-complexity regions of Plasmodium falciparum ABC transporters and associations with in vitro antimalarial responses. Antimicrob. Agents Chemother. 2013, 57, 6196–6204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Velden, M.; Rijpma, S.R.; Russel, F.G.; Sauerwein, R.W.; Koenderink, J.B. PfMDR2 and PfMDR5 are dispensable for Plasmodium falciparum asexual parasite multiplication but change in vitro susceptibility to anti-malarial drugs. Malar. J. 2015, 14, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Parker, D.; Meng, H.; Wu, L.; Li, J.; Zhao, Z.; Zhang, R.; Miao, M.; Fan, Q.; Wang, H.; et al. In vitro sensitivity of Plasmodium falciparum from China-Myanmar border area to major act drugs and polymorphisms in potential target genes. PLoS ONE 2012, 7, e30927. [Google Scholar] [CrossRef]
- Gendrot, M.; Diawara, S.; Madamet, M.; Kounta, M.B.; Briolant, S.; Wade, K.A.; Fall, M.; Benoit, N.; Nakoulima, A.; Amalvict, R.; et al. Association between polymorphisms in the Pfmdr6 Gene and ex vivo susceptibility to quinine in Plasmodium falciparum parasites from Dakar, Senegal. Antimicrob. Agents Chemother. 2017, 61, e01183-16. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.E.; Kirk, K. The malaria parasite’s chloroquine resistance transporter is a member of the drug/metabolite transporter superfamily. Mol. Biol. Evol. 2004, 21, 1938–1949. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, A.B.S.; Verdier-Pinard, D.; Fidock, D.A. Chloroquine resistance in Plasmodium falciparum malaria parasites conferred by Pfcrt mutations. Science 2002, 298, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Lim, P.; Chy, S.; Ariey, F.; Incardona, S.; Chim, P.; Sem, R.; Denis, M.B.; Hewitt, S.; Hoyer, S.; Socheat, D.; et al. Pfcrt polymorphism and chloroquine resistance in Plasmodium falciparum strains isolated in Cambodia. Antimicrob. Agents Chemother. 2003, 47, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Pradines, B.; Gendrot, M.; Delandre, O. Implications des pompes membranaires de Plasmodium falciparum dans le transport et la résistance aux antipaludiques. Rev. Francoph. Lab. 2020, 2020, 59–66. [Google Scholar] [CrossRef]
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.-C.; Khim, N.; Kim, S.; Duru, V.; Bouchier, C.; Ma, L.; et al. A Molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2014, 505, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Ménard, D.; Khim, N.; Beghain, J.; Adegnika, A.A.; Shafiul-Alam, M.; Amodu, O.; Rahim-Awab, G.; Barnadas, C.; Berry, A.; Boum, Y.; et al. A worldwide map of Plasmodium falciparum K13-propeller polymorphisms. N. Engl. J. Med. 2016, 374, 2453–2464. [Google Scholar] [CrossRef] [PubMed]
- Tsombeng, F.F.; Gendrot, M.; Robert, M.G.; Madamet, M.; Pradines, B. Are K13 and Plasmepsin II genes, involved in Plasmodium falciparum resistance to artemisinin derivatives and piperaquine in Southeast Asia, reliable to monitor resistance surveillance in Africa? Malar. J. 2019, 18, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uwimana, A.; Legrand, E.; Stokes, B.H.; Ndikumana, J.-L.M.; Warsame, M.; Umulisa, N.; Ngamije, D.; Munyaneza, T.; Mazarati, J.-B.; Munguti, K.; et al. Emergence and clonal expansion of in vitro artemisinin-resistant Plasmodium falciparum Kelch13 R561H mutant Parasites in Rwanda. Nat. Med. 2020, 26, 1602–1608. [Google Scholar] [CrossRef]
- Bergmann, C.; van Loon, W.; Habarugira, F.; Tacoli, C.; Jäger, J.C.; Savelsberg, D.; Nshimiyimana, F.; Rwamugema, E.; Mbarushimana, D.; Ndoli, J.; et al. Increase in Kelch 13 polymorphisms in Plasmodium falciparum, Southern Rwanda. Emerg. Infect. Dis. 2021, 27, 294–296. [Google Scholar] [CrossRef]
- Silva, M.; Ferreira, P.E.; Otienoburu, S.D.; Calçada, C.; Ngasala, B.; Björman, A.; Martensson, A.; Gil, J.P.; Veiga, M.I. Plasmodium falciparum K13 expression associated with parasite clearance during artemisinin-based combination therapy. J. Antimicrob. Chemother. 2019, 7, 1890–1893. [Google Scholar] [CrossRef]
- Witkowski, B.; Duru, V.; Khim, N.; Ross, L.S.; Saintpierre, B.; Beghain, J.; Chy, S.; Kim, S.; Ke, S.; Kloeung, N.; et al. A Surrogate marker of piperaquine-resistant Plasmodium falciparum malaria: A phenotype-genotype association study. Lancet Infect. Dis. 2017, 17, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Amato, R.; Lim, P.; Miotto, O.; Amaratunga, C.; Dek, D.; Pearson, R.D.; Almagro-Garcia, J.; Neal, A.T.; Sreng, S.; Suon, S.; et al. Genetic markers associated with dihydroartemisinin-piperaquine failure in Plasmodium falciparum malaria in Cambodia: A genotype-phenotype association study. Lancet Infect. Dis. 2017, 17, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Briolant, S.; Bogreau, H.; Gil, M.; Bouchiba, H.; Baret, E.; Amalvict, R.; Rogier, C.; Pradines, B. The F423Y mutation in the Pfmdr2 gene and mutations N51I, C59R, and S108N in the Pfdhfr gene are independently associated with pyrimethamine resistance in Plasmodium falciparum isolates. Antimicrob. Agents Chemother. 2012, 56, 2750–2752. [Google Scholar] [CrossRef] [Green Version]
- Ngo, T.; Duraisingh, M.; Reed, M.; Hipgrave, D.; Biggs, B.; Cowman, A.F. Analysis of Pfcrt, Pfmdr1, Dhfr, and Dhps Mutations and drug sensitivities in Plasmodium falciparum isolates from patients in Vietnam before and after treatment with artemisinin. Am. J. Trop. Med. Hyg. 2003, 68, 350–356. [Google Scholar] [CrossRef]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.J.; Lee, M.; Le Roch, K.G. The ubiquitin system: An essential component to unlocking the secrets of malaria parasite biology. Mol. Biosyst. 2014, 10, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.J.; Lukens, A.K.; Neafsey, D.E.; Schaffner, S.F.; Chang, H.-H.; Valim, C.; Ribacke, U.; Van Tyne, D.; Galinsky, K.; Galligan, M.; et al. Sequence-based association and selection scans identify drug resistance loci in the Plasmodium falciparum malaria parasite. Proc. Natl. Acad. Sci. USA 2012, 109, 13052–13057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, C.P.; Liu, C.-H.; Mayer, S.; Nurhasanah, A.; Cyrklaff, M.; Mu, J.; Ferdig, M.T.; Stein, W.D.; Lanzer, M. A HECT ubiquitin-Protein ligase as a novel candidate gene for altered quinine and quinidine responses in Plasmodium falciparum. PLoS Genet. 2014, 10, e1004382. [Google Scholar] [CrossRef] [PubMed]
- Ribacke, U.; Bartlett, M.; Patel, S.D.; Seneratne, N.; Park, D.J.; Duraisingh, M.; Sabeti, P.C.; Volkman, S.K.; Wirth, D.F. Adaptative evolution of a ring ubiquitin ligase mediates reduced drug sensitivity in Plasmodium falciparum. In Proceedings of the 61st Meeting of American Society of Tropical Medicine and Hygiene, Atlanta, GA, USA, 11–15 November 2012; p. 299. [Google Scholar]
- Gendrot, M.; Fall, B.; Madamet, M.; Fall, M.; Wade, K.A.; Amalvict, R.; Nakoulima, A.; Benoit, N.; Diawara, S.; Diémé, Y.; et al. Absence of association between polymorphisms in the RING E3 ubiquitin protein ligase gene and ex vivo susceptibility to conventional antimalarial drugs in Plasmodium falciparum isolates from Dakar, Senegal. Antimicrob. Agents Chemother. 2016, 60, 5010–5013. [Google Scholar] [CrossRef] [Green Version]
- Demas, A.R.; Sharma, A.I.; Wong, W.; Early, A.M.; Redmond, S.; Bopp, S.; Neafsey, D.E.; Volkman, S.K.; Hartl, D.L.; Wirth, D.F. Mutations in Plasmodium falciparum actin-binding protein coronin confer reduced artemisinin susceptibility. Proc. Natl. Acad. Sci. USA 2018, 115, 12799–12804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olshina, M.A.; Angrisano, F.; Marapana, D.S.; Riglar, D.T.; Bane, K.; Wong, W.; Catimel, B.; Yin, M.-X.; Holmes, A.B.; Frischknecht, F.; et al. Plasmodium falciparum coronin organizes arrays of parallel actin filaments potentially guiding directional motility in invasive malaria parasites. Malar. J. 2015, 14, 280. [Google Scholar] [CrossRef] [Green Version]
- Tardieux, I.; Liu, X.; Poupel, O.; Parzy, D.; Dehoux, P.; Langsley, G. A Plasmodium falciparum novel gene encoding a coronin-like protein which associates with actin filaments. FEBS Lett. 1998, 441, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.I.; Shin, S.H.; Bopp, S.; Volkman, S.K.; Hartl, D.L.; Wirth, D.F. Genetic background and PfKelch13 affect artemisinin susceptibility of PfCoronin mutants in Plasmodium falciparum. PLoS Genet. 2020, 16, e1009266. [Google Scholar] [CrossRef]
- Delandre, O.; Daffe, S.M.; Gendrot, M.; Diallo, M.N.; Madamet, M.; Kounta, M.B.; Diop, M.N.; Bercion, R.; Sow, A.; Ngom, P.M.; et al. Absence of association between polymorphisms in the Pfcoronin and Pfk13 genes and the presence of Plasmodium falciparum parasites after treatment with artemisinin derivatives in Senegal. Int. J. Antimicrob. Agents 2020, 56, 106190. [Google Scholar] [CrossRef]
- Martin, R.E.; Henry, R.I.; Abbey, J.L.; Clements, J.D.; Kirk, K. The “permeome” of the malaria parasite: An overview of the membrane transport proteins of Plasmodium falciparum. Genome Biol. 2005, 6, R26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, M.Y.-X.; LaMonte, G.; Lee, M.C.S.; Reimer, C.; Tan, B.H.; Corey, V.; Tjahjadi, B.F.; Chua, A.; Nachon, M.; Wintjens, R.; et al. UDP-Galactose and Acetyl-CoA transporters as Plasmodium multidrug resistance genes. Nat. Microbiol. 2016, 1, 16166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhen, K.L.; Chatterjee, A.K.; Rottmann, M.; Gagaring, K.; Borboa, R.; Buenviaje, J.; Chen, Z.; Francek, C.; Wu, T.; Nagle, A.; et al. KAF156 Is an antimalarial clinical candidate with potential for use in prophylaxis, treatment, and prevention of disease transmission. Antimicrob. Agents Chemother. 2014, 58, 5060–5067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaMonte, G.; Lim, M.Y.-X.; Wree, M.; Reimer, C.; Nachon, M.; Corey, V.; Gedeck, P.; Plouffe, D.; Du, A.; Figueroa, N.; et al. Mutations in the Plasmodium falciparum cyclic amine resistance locus (PfCARL) confer multidrug resistance. MBio 2016, 7, e00696-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magistrado, P.A.; Corey, V.C.; Lukens, A.K.; LaMonte, G.; Sasaki, E.; Meister, S.; Wree, M.; Winzeler, E.; Wirth, D.F. Plasmodium falciparum cyclic amine resistance locus (PfCARL), a resistance mechanism for two distinct compound classes. ACS Infect. Dis. 2016, 2, 816–826. [Google Scholar] [CrossRef]
- Jonikas, M.C.; Collins, S.R.; Denic, V.; Oh, E.; Quan, E.M.; Schmid, V.; Weibezahn, J.; Schwappach, B.; Walter, P.; Weissman, J.S.; et al. Comprehensive characterization of genes required for protein folding in the endoplasmic reticulum. Science 2009, 323, 1693–1697. [Google Scholar] [CrossRef] [Green Version]
- Foguim, F.T.; Robert, M.G.; Gueye, M.W.; Gendrot, M.; Diawara, S.; Mosnier, J.; Amalvict, R.; Benoit, N.; Bercion, R.; Fall, B. Low polymorphisms in Pfact, Pfugt and Pfcarl genes in African Plasmodium falciparum isolates and absence of association with susceptibility to common anti-malarial drugs. Malar. J. 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Ridley, R.G.; Dorn, A.; Vippagunta, S.R.; Vennerstrom, J.L. Haematin (haem) polymerization and its inhibition by quinoline antimalarials. Ann. Trop. Med. Parasitol. 1997, 91, 559–566. [Google Scholar] [CrossRef]
- Fidock, D.A.; Nomura, T.; Talley, A.K.; Cooper, R.A.; Dzekunov, S.M.; Ferdig, M.T.; Ursos, L.M.; Sidhu, A.B.; Naudé, B.; Deitsch, K.W.; et al. Mutations in the P. Falciparum digestive vacuole transmembrane protein PfCRT and evidence for their role in chloroquine resistance. Mol. Cell 2000, 6, 861–871. [Google Scholar] [CrossRef]
- Djimdé, A.; Doumbo, O.K.; Cortese, J.F.; Kayentao, K.; Doumbo, S.; Diourté, Y.; Coulibaly, D.; Dicko, A.; Su, X.Z.; Nomura, T.; et al. A molecular marker for chloroquine-resistant falciparum malaria. N. Engl. J. Med. 2001, 344, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Veiga, M.I.; Dhingra, S.K.; Henrich, P.P.; Straimer, J.; Gnädig, N.; Uhlemann, A.-C.; Martin, R.E.; Lehane, A.M.; Fidock, D.A. Globally prevalent PfMDR1 mutations modulate Plasmodium falciparum susceptibility to artemisinin-based combination therapies. Nat. Commun. 2016, 7, 11553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisowath, C.; Strömberg, J.; Mårtensson, A.; Msellem, M.; Obondo, C.; Björkman, A.; Gil, J.P. In vivo selection of Plasmodiumf falciparum Pfmdr1 86N coding alleles by artemether-lumefantrine (Coartem). J. Infect. Dis. 2005, 191, 1014–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dokomajilar, C.; Nsobya, S.L.; Greenhouse, B.; Rosenthal, P.J.; Dorsey, G. Selection of Plasmodium falciparum Pfmdr1 alleles following therapy with artemether-lumefantrine in an area of Uganda where malaria is highly endemic. Antimicrob. Agents Chemother. 2006, 50, 1893–1895. [Google Scholar] [CrossRef] [Green Version]
- Price, R.N.; Uhlemann, A.-C.; Brockman, A.; McGready, R.; Ashley, E.; Phaipun, L.; Patel, R.; Laing, K.; Looareesuwan, S.; White, N.J.; et al. Mefloquine resistance in Plasmodium falciparum and increased Pfmdr1 gene copy number. Lancet 2004, 364, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, M.; Gadalla, N.B.; Stepniewska, K.; Dahal, P.; Nsanzabana, C.; Moriera, C.; Price, R.N.; Mårtensson, A.; Rosenthal, P.J.; Dorsey, G.; et al. Polymorphisms in Plasmodium falciparum chloroquine resistance transporter and multidrug resistance 1 genes: Parasite risk factors that affect treatment outcomes for P. falciparum malaria after Artemether-Lumefantrine and Artesunate-Amodiaquine. Am. J. Trop. Med. Hyg. 2014, 91, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.D.; LeClair, N.; Arinaitwe, E.; Wanzira, H.; Kakuru, A.; Bigira, V.; Muhindo, M.; Kamya, M.R.; Tappero, J.W.; Greenhouse, B.; et al. Comparative impacts over 5 years of artemisinin-based combination therapies on Plasmodium falciparum polymorphisms that modulate drug sensitivity in Ugandan children. J. Infect. Dis. 2014, 210, 344–353. [Google Scholar] [CrossRef] [Green Version]
- Nankabirwa, J.I.; Conrad, M.D.; Legac, J.; Tukwasibwe, S.; Tumwebaze, P.; Wandera, B.; Brooker, S.J.; Staedke, S.G.; Kamya, M.R.; Nsobya, S.L.; et al. Intermittent preventive treatment with dihydroartemisinin-piperaquine in Ugandan schoolchildren selects for Plasmodium falciparum transporter polymorphisms that modify drug sensitivity. Antimicrob. Agents Chemother. 2016, 60, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Holmgren, G.; Gil, J.P.; Ferreira, P.M.; Veiga, M.I.; Obonyo, C.O.; Björkman, A. Amodiaquine resistant Plasmodium falciparum malaria in vivo is associated with selection of Pfcrt 76T and Pfmdr1 86Y. Infect. Genet. Evol. 2006, 6, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, G.; Hamrin, J.; Svärd, J.; Mårtensson, A.; Gil, J.P.; Björkman, A. Selection of Pfmdr1 mutations after amodiaquine monotherapy and amodiaquine plus artemisinin combination therapy in East Africa. Infect. Genet. Evol. 2007, 7, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Danquah, I.; Coulibaly, B.; Meissner, P.; Petruschke, I.; Müller, O.; Mockenhaupt, F.P. Selection of Pfmdr1 and Pfcrt alleles in amodiaquine treatment failure in North-Western Burkina Faso. Acta Trop. 2010, 114, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Yeka, A.; Kigozi, R.; Conrad, M.D.; Lugemwa, M.; Okui, P.; Katureebe, C.; Belay, K.; Kapella, B.K.; Chang, M.A.; Kamya, M.R.; et al. Artesunate/Amodiaquine versus Artemether/Lumefantrine for the treatment of uncomplicated malaria in Uganda: A randomized trial. J. Infect. Dis. 2016, 213, 1134–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picot, S.; Olliaro, P.; de Monbrison, F.; Bienvenu, A.-L.; Price, R.N.; Ringwald, P. A Systematic review and meta-analysis of evidence for correlation between molecular markers of parasite resistance and treatment outcome in falciparum malaria. Malar. J. 2009, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaillard, T.; Briolant, S.; Houzé, S.; Baragatti, M.; Wurtz, N.; Hubert, V.; Lavina, M.; Pascual, A.; Travaillé, C.; Le Bras, J.; et al. PftetQ and Pfmdt copy numbers as predictive molecular markers of decreased ex vivo doxycycline susceptibility in imported Plasmodium falciparum malaria. Malar. J. 2013, 12, 414. [Google Scholar] [CrossRef] [Green Version]
- Gendrot, M.; Foguim, F.T.; Robert, M.G.; Amalvict, R.; Mosnier, J.; Benoit, N.; Madamet, M.; Pradines, B. The D113N mutation in the RING E3 ubiquitin protein ligase gene is not associated with ex vivo susceptibility to common anti-malarial drugs in African Plasmodium falciparum isolates. Malar. J. 2018, 17, 108. [Google Scholar] [CrossRef] [Green Version]
- Bogreau, H.; Renaud, F.; Bouchiba, H.; Durand, P.; Assi, S.-B.; Henry, M.-C.; Garnotel, E.; Pradines, B.; Fusai, T.; Wade, B.; et al. Genetic diversity and structure of African Plasmodium falciparum populations in urban and rural areas. Am. J. Trop. Med. Hyg. 2006, 74, 953–959. [Google Scholar] [CrossRef]
- Henry, M.; Diallo, I.; Bordes, J.; Ka, S.; Pradines, B.; Diatta, B.; M’Baye, P.S.; Sane, M.; Thiam, M.; Gueye, P.M.; et al. Urban malaria in Dakar, Senegal: Chemosusceptibility and genetic diversity of Plasmodium falciparum isolates. Am. J. Trop. Med. Hyg. 2006, 75, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Foguim, F.T.; Bogreau, H.; Gendrot, M.; Mosnier, J.; Fonta, I.; Benoit, N.; Amalvict, R.; Madamet, M.; Wein, S.; Pradines, B. Prevalence of mutations in the Plasmodium falciparum chloroquine resistance transporter, PfCRT, and association with ex vivo susceptibility to common anti-malarial drugs against African Plasmodium falciparum isolates. Malar. J. 2020, 19, 1–9. [Google Scholar] [CrossRef]
- Torrentino-Madamet, M.; Fall, B.; Benoit, N.; Camara, C.; Amalvict, R.; Fall, M.; Dionne, P.; Ba Fall, K.; Nakoulima, A.; Diatta, B.; et al. Limited polymorphisms in K13 Gene in Plasmodium falciparum isolates from Dakar, Senegal in 2012–2013. Malar. J. 2014, 13, 472. [Google Scholar] [CrossRef] [Green Version]
- Robert, M.G.; Tsombeng, F.F.; Gendrot, M.; Mosnier, J.; Amalvict, R.; Benoit, N.; Torrentino-Madamet, M.; Pradines, B. Absence of a high level of duplication of the Plasmepsin II gene in Africa. Antimicrob. Agents Chemother. 2018, 62, e00374-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, M.G.; Tsombeng, F.F.; Gendrot, M.; Diawara, S.; Madamet, M.; Kounta, M.B.; Wade, K.A.; Fall, M.; Gueye, M.W.; Benoit, N. Baseline ex vivo and molecular responses of Plasmodium falciparum isolates to piperaquine before implementation of dihydroartemisinin-piperaquine in Senegal. Antimicrob. Agents Chemother. 2019, 63, e02445-18. [Google Scholar] [CrossRef] [Green Version]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Veiga, M.I.; Ferreira, P.E.; Malmberg, M.; Jörnhagen, L.; Björkman, A.; Nosten, F.; Gil, J.P. Pfmdr1 amplification is related to increased Plasmodium falciparum in vitro sensitivity to the bisquinoleine piperaquine. Antimicrob. Agents Chemother. 2012, 56, 3615–3619. [Google Scholar] [CrossRef] [Green Version]
- Raj, D.K.; Mu, J.; Jiang, H.; Kabat, J.; Singh, S.; Sullivan, M.; Fay, M.P.; McCutchan, T.F.; Su, X.Z. Disruption of a Plasmodium falciparum multidrug resistance-associated protein (PfMRP) alters its fitness and transport of antimalarial drugs and glutathione. J. Biol. Chem. 2009, 284, 7687–7696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Pluijm, R.W.; Imwong, M.; Chau, N.H.; Hoa, N.T.; Thuy-Nhien, N.T.; Thanh, N.V.; Jittamala, P.; Hanboonkunupakarn, B.; Chutasmit, K.; Saelow, C.; et al. Determinants of dihydroartemisinin-piperaquine treatment failure in Plasmodium falciparum malaria in Cambodia, Thailand, and Vietnam: A prospective clinical, pharmacological, and genetic study. Lancet Infect. Dis. 2019, 19, 952–961. [Google Scholar] [CrossRef] [Green Version]
- Ahn, C.X.; Chavchich, M.; Birrell, G.W.; van Breda, K.; Travers, T.; Rowcliffe, K.; Lord, A.R.; Shanks, G.D.; Edstein, M.D. Pharmacokinetics and ex vivo antimalarial ativity of artesunate-amodiaquine plus methylene blue in healthy volunteers. Antimicrob. Agents Chemother. 2020, 64, e01441-19. [Google Scholar] [CrossRef]
- Jorge, M.M.; Ouermi, L.; Meissner, P.; Compaoré, G.; Coulibaly, B.; Nebie, E.; Krisam, J.; Klose, C.; Kieser, M.; Jahn, A.; et al. Safety and efficacy of artesunate-amodiaquine combined with either methylene blue or primaquine in children with falciparum malaria in Burkiba Faso: A randomized controlled trial. PLoS ONE 2019, 14, e0222993. [Google Scholar] [CrossRef]


{kind=link}
| Gene | Polymorphisms Previously Described | Association with Antimalarial Susceptibility Modulation | References |
|---|---|---|---|
| Pfmdr1 | N86Y, Y184, S1034C, N1042D, D1246Y mutations | chloroquine, quinine, monodesethylamodiaquine, mefloquine, lumefantrine, dihydroartemisinin | [33,34,36,37] |
| Pfmdr5 | DNNN, DHHNDHNNDNNN repetitions | lumefantrine, piperaquine | [36,39] |
| Pfmdr6 | Asn repetitions between amino acid positions 103 to 109 in 3D7 | lumefantrine (6 and 7 Asn), piperaquine (8 Asn), dihydroartemisinin (9 Asn) | [39,41,42] |
| Pfcrt | M74I, N75E, K76T | chloroquine, amodiaquine | [44] |
| PfK13 | M476I, Y493H, R539T, I543T, R561H, P574L, C580Y mutations | artemisinin | [7,47,48,50,51] |
| Pfmp2 | Gene copy amplification | piperaquine | [53,54] |
| Pfdhfr | N51I, C59R, S108N mutations | pyrimethamine, proguanil | [36,55,56] |
| Pfubq | D113N mutation | chloroquine, amodiaquine | [61] |
| Pfcoronin | G50E, R100K, E107V | artemisinin | [63,66] |
| Pfact | A94T, R108K, S110R, D165N, G559K mutations | imidazolopiperazines | [69] |
| Pfugt | F37V | imidazolopiperazines | [69] |
| Pfcarl | K734M, L830V, S1076N/I, V1103L, I1139K | pyronaridine, imidazolopiperazines | [69,70,71,72,74] |
| Haplotype | Number | Frequency (%) | Mean IC50 in nM | 95% CI in nM | p-Value |
|---|---|---|---|---|---|
| N86 vs. 86Y | 433/37 | 90.2/8.5 | 11.7/9.7 | 10.5–12.8/5.9–13.5 | 0.31 |
| Y184 vs. 184 F | 209/237 | 44.0/49.8 | 11.4/11.1 | 9.8–13.0/9.6–12.5 | 0.78 |
| D1246 vs. 1246 Y | 441/2 | 99.5/0.5 | 11.3/7.4 | ||
| N86-Y184 vs. N86-184F | 187/209 | 47.2/52.8 | 11.2/11.6 | 9.5–12.9/10.0–13.1 | 0.74 |
| N86-184F vs. all | 209/260 | 44.6/55.4 | 11.6/11.4 | 10.0–13.1/10.0–12.7 | 0.58 |
| 86Y-184F vs. all | 23/446 | 4.9/95.1 | 8.0/11.6 | 3.3–12.7/10.6–12.8 | 0.03 |
| 86Y-Y184 vs. all | 13/456 | 2.8/97.2 | 11.4/12.9 | 10.4–12.5/6.6–19.3 | 0.70 |
| Allele | Number | Frequency (%) | Methylene Blue IC50 in nM | ||
|---|---|---|---|---|---|
| Mean | 95% CI | p-Value | |||
| 3R1-1R2 | 3 | 0.8 | 10.3 | ||
| 4R1-1R2 | 3 | 0.8 | 9.3 | ||
| 5R1-1R2 | 110 | 29.0 | 12.7 | 10.3–15.1 | 0.76 a |
| 6R1-1R2 | 82 | 21.6 | 13.3 | 10.5–16.1 | 0.46 a |
| 7R1-1R2 | 101 | 26.6 | 12.4 | 9.6–14.9 | 0.99 a |
| 8R1-1R2 | 22 | 5.8 | 10.7 | 5.3–16.1 | 0.47 a |
| 9R1-1R2 | 7 | 1.8 | 7.4 | ||
| 10R1-1R2 | 3 | 0.8 | 8.5 | ||
| 11R1-1R2 | 1 | 0.3 | 8.9 | ||
| 12R1-1R2 | 1 | 0.3 | 0.2 | ||
| 13R1-1R2 | 1 | 0.3 | 17.5 | ||
| 5R1-2R2 | 13 | 3.4 | 11.7 | 4.7–13.4 | 0.83 a |
| 0R2 | 4 | 1.1 | 9.1 | ||
| 1R2 | 351 | 92.6 | 12.1 | 10.8–13.4 | 0.36 a |
| 2R2 | 24 | 6.3 | 13.3 | 8.2–18.3 | 0.67 a |
| ≤8R1-1R1 | 349 | 84.6 | 12.6 | 11.2–13.9 | 0.08 b |
| >8R1-1R1 | 13 | 3.2 | 8.0 | 1.0–14.9 | 0.08 b |
| Asn Repetition | Number | Frequency (%) | Mean IC50 in nM | 95% CI in nM | p-Value |
|---|---|---|---|---|---|
| 5 | 2 | 0.5 | 11.9 | ||
| 6 | 6 | 1.7 | 15.7 | 5.8–25.6 | 0.53 |
| 7 | 28 | 8.0 | 11.2 | 6.6–15.8 | 0.72 |
| 8 | 56 | 16.0 | 13.2 | 10.0–16.5 | 0.47 |
| 9 | 177 | 50.7 | 12.7 | 10.9–14.5 | 0.31 |
| 10 | 51 | 17.8 | 10.3 | 5.1–15.5 | 0.45 |
| 11 | 23 | 6.5 | 10.3 | 5.1–15.5 | 0.45 |
| 12 | 2 | 0.5 | 8.9 | ||
| 13 | 2 | 0.5 | 3.4 | ||
| 14 | 3 | 0.8 | 12.0 |
| Asn Repetition | Number | Frequency (%) | Mean IC50 in nM | 95% CI in nM | p-Value |
|---|---|---|---|---|---|
| 4 | 5 | 1 | 13.3 | 2.8–23.7 | 0.81 |
| 5 | 1 | 0.2 | |||
| 6 | 217 | 45.0 | 11.6 | 10.0–13.2 | 0.98 |
| 7 | 71 | 14.7 | 10.6 | 7.8–13.3 | 0.42 |
| 8 | 83 | 17.2 | 12.5 | 9.9–15.0 | 0.41 |
| 9 | 64 | 13.3 | 12.8 | 9.9–15.8 | 0.46 |
| 10 | 25 | 5.2 | 10.6 | 6.0–15.3 | 0.62 |
| 11 | 7 | 1.5 | 11.4 | 2.6–20.2 | 0.95 |
| 12 | 7 | 1.5 | 4.6 | 0.1–13.4 | 0.01 |
| 14 | 1 | 0.2 | |||
| 18 | 1 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gendrot, M.; Delandre, O.; Robert, M.G.; Foguim, F.T.; Benoit, N.; Amalvict, R.; Fonta, I.; Mosnier, J.; Madamet, M.; Pradines, B.; et al. Absence of Association between Methylene Blue Reduced Susceptibility and Polymorphisms in 12 Genes Involved in Antimalarial Drug Resistance in African Plasmodium falciparum. Pharmaceuticals 2021, 14, 351. https://doi.org/10.3390/ph14040351
Gendrot M, Delandre O, Robert MG, Foguim FT, Benoit N, Amalvict R, Fonta I, Mosnier J, Madamet M, Pradines B, et al. Absence of Association between Methylene Blue Reduced Susceptibility and Polymorphisms in 12 Genes Involved in Antimalarial Drug Resistance in African Plasmodium falciparum. Pharmaceuticals. 2021; 14(4):351. https://doi.org/10.3390/ph14040351
Chicago/Turabian StyleGendrot, Mathieu, Océane Delandre, Marie Gladys Robert, Francis Tsombeng Foguim, Nicolas Benoit, Rémy Amalvict, Isabelle Fonta, Joel Mosnier, Marylin Madamet, Bruno Pradines, and et al. 2021. "Absence of Association between Methylene Blue Reduced Susceptibility and Polymorphisms in 12 Genes Involved in Antimalarial Drug Resistance in African Plasmodium falciparum" Pharmaceuticals 14, no. 4: 351. https://doi.org/10.3390/ph14040351