
Bird Feeder Explorers Are Not Attracted by the Seeds of Invasive Weeds in Winter

Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska Street 17, 31-016 Kraków, Poland
Diversity 2024, 16(2), 81; https://doi.org/10.3390/d16020081
Submission received: 21 December 2023
/
Revised: 19 January 2024
/
Accepted: 24 January 2024
/
Published: 26 January 2024
(This article belongs to the Special Issue Recent Advances in the Impacts of Biological Invasions on European Biodiversity)
Abstract
:Winter feeding of birds is one of the most widespread forms of human influence on animals in the world. Urban areas, however, contain many other food sources apart from the contents of feeders, including contaminated food and seeds of invasive plants. This research aimed to investigate whether birds exploring urban food select it, giving up potentially toxic or unknown seeds, or whether they use every available food. In the winter of the year 2022, an experiment was carried out with 18 feeders in two city parks in Kraków, Poland, which was based on feeding birds using sunflower seeds with various admixtures of the invasive Caucasian hogweeds Heracleum sp. seeds. Despite the detection of pecking marks on the hogweed seeds, only a small part of them disappeared from the feeders compared to the intensive feeding of birds on sunflower seeds. This result shows that not every new food source in the city can be used by birds. Urban explorers remain selective when choosing food, which keeps their behaviour close to non-urban populations.
1. Introduction
The phenomenon of winter feeding of birds is among the commonest interactions between animals and humans on a global scale [1], especially in the Western world [2]. Birds use feeders during critical periods like winter as they depend on available resources. Feeders are particularly common and exploited in urban areas [1], which means that birds living in such areas are provided with considerable amounts and variety of food year-round [2]. The provisioning of birds in feeders can determine their further life-history traits such as survival and the phenology of the following breeding season [3]. Although feeding may have the expected positive effect, i.e., provisioning of nutritious food, it also has negative consequences, such as increased competition or disease transmission [1]. Surprisingly, remarkably little is known about the outcomes and implications of what may be seen as a feeding experiment on a massive scale [2].
Feeding has an element of unpredictability—birds are unable to predict how much and what kind of food they can expect at a specific place and time. Birds’ perceptions of risk are biased mainly towards starvation avoidance in winter [4]. Daily patterns in the foraging behaviour of birds are assumed to balance the counteracting risks of predation and starvation. Thus, foraging in most species is completed before sunset [4]. The foraging activity of birds in areas with feeding also depends on temperature and the need for social life. Individuals with stronger associations in a social network are more likely to share similar feeding activity [5]. On the one hand, species foraging similarly can be more likely facilitated in flocks, but on the other hand, a greater gathering of birds increases unfavourable competition for resources. However, it turns out that both phenomena—social needs and competition—enhance the flocking of foraging birds [6]. The aspect of flocking birds near feeding points should be treated more broadly in studies conducted in areas with supplementary feeding.
The other important aspect is the location of the winter feeding point—the preferred choice should be a feeder that is inaccessible to predators but in an area open enough to allow them to be seen by birds. The visibility of potential threats is easier to spot in more open habitats. It is known from studies in breeding season that birds prefer to feed in pastures with better observation possibilities than in meadows, despite better food availability there [7]. For the birds foraging on the ground (including those using food that fell out of the feeder or which was left on the ground from the beginning), earlier predator detection when vegetation is lower may play a crucial role [8]. Foraging efficiency, as measured by the amount of food eaten in a particular place, is the result of individuals’ ability to reach a feeding site and balance risky foraging behaviours in local conditions [9,10]. The type of vegetation structure also shapes the availability of food items and hence affects the foraging success of birds [11]. Typically, birds move just far enough to reach areas they have not previously searched visually [12]. Therefore, animal behaviour should always be treated as an integral component reflecting birds’ perception of risk in urban landscape connectivity [10].
Five main components describe foraging activity and should be taken into account during its investigation: food search, attack, foraging site, food type, and food handling. Quantifying the foraging site involves the description of the habitat and its surroundings, vertical position, foliage density, and substrate [13]. Regarding food type and food search/evaluation in natural conditions, an animal normally faces more than one food item simultaneously, assesses them, and then chooses the preferred item and rejects the others [14]. Although the disappeared seeds are generally assumed to be a measure of foraging effort, some birds could be more selective, throwing seeds out of the feeder and making them less available for other, less selective, birds. To solve this problem, feeders should be regularly refilled with food to provide equal opportunities for different birds when choosing food items. Moreover, multiple-offer experiments including various food items similarly exposed in feeders are recommended over simple-offer ones because, in this way, the consumer has a natural opportunity to express his dietary choice [15].
Sunflower seeds are generally the most preferred winter food supplement both in urban and rural habitats and are significantly more exploited than any other food simultaneously available (in this case, animal fat, millet seed, and dry fruits of rowanberry) [1]. The preferred food can be collected by birds in both beech and spruce forests with no difference between these habitats [16], allowing for relative freedom in the selection of habitats and location of winter feeding points. Snow cover and periods of extremely low temperatures affect the activity of birds but, surprisingly, have no impact on their food preferences. Feeding preferences, however, change at the onset of breeding (about mid-March in the temperate zone), because the availability of natural food, especially arthropods, increases, and nutritional needs become higher before the upcoming breeding season [16].
Food left in feeders is usually assumed to be beneficial for birds, although this is difficult to assess. This food contains not only fats, but also many chemical components related to the processing of grains, or even deposits of pollutants from urban areas. Meanwhile, the chemical properties of winter food determine which individuals survive this period. Breeding populations of Eurasian blue tits Cyanistes caeruleus which had access to vitamin E-rich food during the previous winter contained birds with reduced feather carotenoid concentrations. This was explained by the enhanced survival and recruitment of lower-quality individuals unable to absorb vitamins without winter feeding [17].
It has not been investigated whether birds using winter feeders are still selective in choosing food in case harmful seeds appear in them. Moreover, urban areas are unexplored as feeding places for birds outside feeders. For example, urban areas are full of seeds of expansive and invasive plant species. Can birds reach for invaders’ seeds as they are easily available? The research presented in this article aimed to investigate whether wintering birds using feeding in urban areas select the seeds of the toxic invasive Caucasian hogweeds Heracleum sp. This research involved the installation of a system of feeders in two city parks, in which preferred sunflower seeds with an admixture of invasive hogweed seeds with other properties were placed. The experiment was conducted at the end of the wintering of birds to increase the likelihood of choosing food items other than oilseeds (e.g., sunflower seeds) preferred in winter. In this way, the following hypotheses were tested: 1. Birds using feeders are selective in the food they choose, i.e., they evaluate the available food and select particular food items. It is expected that the birds will feed on sunflower seeds left in feeders. Since it is not known whether birds feed on Caucasian hogweed seeds, it is difficult to determine whether selection will be positive or if these seeds will be rejected. 2. Because the proportions of the two types of food were assumed to be variable at different feeders, reflecting the unpredictability of food availability in urban areas, it is expected that the location of the feeder and the effects of bird feeding on food in neighbouring feeders will not affect the consumption or abandonment of food at individual feedings points. The conservation aspect of the research is to assess whether birds consider hogweed seeds to be an attractive food and to verify the suspicion that they may spread seeds of plant invaders such as hogweeds.
2. Materials and Methods
2.1. Feeding Sites
The author established feeding sites (total n = 18) in two semi-natural urban parks, namely, Bednarski Park (n = 10 feeding sites) and Jordan Park (n = 8), Kraków, Poland. The feeders were installed constantly in February and March 2022, after the period of the greatest frosts and shortly before the breeding season, when birds begin to eat more diverse food than just sunflowers [16]. In this way, the author tried to increase the likelihood of using seeds other than sunflowers by birds. The sample size was low due to the following reasons: (a) the number of feeders had to reflect the real urban feeding intensity—usually there are only a few feeders per park; (b) the feed mixture contained seeds of invasive and toxic plants, and only a small number of feeders made it possible to control and clean them thoroughly; (c) city parks were small, and feeders were installed at least every 50 m, as a larger number of feeders would be too dense and likely to be visited by the same birds, which would create the problem of pseudoreplication in the analyses.
Feeding sites within each park represented different landscape fragmentation configurations and external contexts, including urban activities and potential risks. During the experimental period, Bednarski Park was undergoing modernization, there was relatively noisy equipment working there, and the number of visitors was small because only a part of it (which contained bird feeders) was open to the public. It is a park located in the old, quiet district of the city. Jordan Park was visited by crowds of people, and it is adjacent to several academic centres and busy streets. Although it is known that the local habitat in urban areas does not influence the use of winter supplementary food by birds [16], the experiment in two parks took into account the variability in habitats. The Bednarski Park contains many old trees, mainly deciduous ones. The part of Jordan Park where the research was conducted contains younger trees than Bednarski Park, with many conifers.
In both parks, people fed the birds and other feeders were present, but not in the areas designated for the experiment. Nevertheless, it was assumed that the previous presence of feeders in the studied parks was a sufficient predictor that the birds knew the feeding process and would look for artificial food sources in the selected parks.
2.2. The Concept of Experimental Feeding
All feeders were filled with food 7 times (n = 126 episodes in 18 feeders) at different dates during the experiment, and the volume of food left (i.e., supplied) in the feeders was always the same. Taking into account the recommendation to perform multiple-food experiments rather than those with a single food source [15], a mix of seeds was used in this research. The basis was sunflower seeds, which were variously supplemented with Sosnowsky’s hogweed Heracleum sosnowskyi seeds. Because it is an invasive plant [18], the seeds were previously inactivated using a UV crosslinker (CL-1000, UVP) in one day and with the same dose of radiation to destroy some of the proteins and prevent seed germination. The exposure concerned the internal structure of the seeds and was short, it did not leave any traces on the seeds, so it was assumed it would not affect the results. Previously, field-collected hogweed seeds were stored for one year at room temperature.
For two months, each feeder was visited 15–16 times (i.e., work on it took place for 10 days, i.e., 7 days of food supplementation - including 6 days of seed counting as a result of feeding—with 3 days of cleaning the feeders, including a thorough cleaning of hogweed seeds, while another 5–6 days were to indicate no food supplementation by other people). A single field visit concerned all feeders in one park in various order and lasted from around 7 a.m. until early afternoon. Feeders, if the food was left in them, were visited and observed daily to ensure sufficient contents, that the seed mix did not freeze into a single mass, and to count the seeds each day. After seed utilization counts were completed, the feeders were emptied for the next few days and cleaned.
The feeders were standard plastic pots for plants (among the gardeners called “productive pots”), brown or black, and hung in the field on a jute string at a constant height of about 1.2 m above the ground on shrubs or young trees surrounded by open area (Figure 1). Thus, none of the feeders were lost nor aroused people’s interest. They were too unstable to increase predator pressure. Pots are not used as feeders, which allowed the author to assume that they would not be supplemented with food by other people. As mentioned, during periods when the pots were not filled with food by the researcher, they were visited 5–6 times during the entire experiment to confirm the absence of unknown food.
The productive pot used with a diameter of 12 cm was standard, and its volume was 0.8 L. Because the pot was thinner at the base, filling it halfway did not ensure that half the volume was filled. Therefore, before the experiment, an example pot with holes in the bottom covered with paper was filled with a volume of 0.4 L of sunflower seeds measured in a measuring cup. On the inner wall of the pot, the point reached by the seeds was marked with a waterproof marker, and the distance of this point from the bottom of the pot was measured. A similar marking was placed on the inner wall of each pot.
In the days of pot supplementing with sunflower and hogweed seeds, the volume of the mixture always reached the marked points. At first, to prevent the seeds from spilling out of the pots, a fragment of a flat leaf (resembling the paper—see above) was placed at the bottom to cover the drainage holes. When supplementing the seeds, their types were placed in two layers—sunflowers below, and hogweeds on top, assessing what percentage of the 0.4 L volume each of them constituted (for example, 75% of the volume, half of the volume, or 20% of the volume of the pot next to the bottom, and such simple measures were used). The seeds were mixed during every second food refill and, on other visits, left as divided layers. If subsequent seeds were added to pots, the seeds were always removed (also when mixed) and sorted before placing in the pot.
The hogweed seeds were counted piece by piece before leaving them in the feeder. During counting, hogweed seeds were removed from the pot and searched on the ground in the surrounding area, as some of them were soaked in water, which could affect the birds’ further interest. If a seed mixture was again placed in a pot, there were always sunflowers near the bottom and new hogweed seeds on top filling the pot to a marked line.
The analysis used the volume of seeds determined by the percentage of their content in the 0.4 L volume. The volume of seeds during counting (sunflowers and hogweeds separately) was assessed in the field as a percentage of 0.4 L using an empty pot with a marked point, so the difference between the volume of seeds in a pot the day before and that measured was used as a measure of foraging efficiency.
The observations of feeders lasting 3–4 h during a single visit to a given park and observations while counting and supplementing seeds allowed for the author to note the presence of the main bird species visiting feeders and also foraging on the ground below.
2.3. Statistical Analysis
All analyses were prepared in R 4.0.4 [19]. The distribution of dependent variables (i.e., the percentage of food on a given day at each feeding point supplied and eaten separately) was investigated with the Shapiro–Wilk test. The distribution of the proportion of food supplied differed from normality (W = 0.739, p-value < 0.001), as did the one of food eaten (W = 0.699, p-value < 0.001). To confirm that the proportions of food supplied and eaten varied in the feeders, i.e., when choosing given seeds, the birds were not influenced by what was happening in the neighbouring feeders (in the meaning of local context—more favourable location of a pot, lower predation risk, etc.), including the nearby activity of other birds, the autocorrelation between the seed proportions in the nearest feeders (k = 5) was assessed using Moran I test based on the geographical coordinates (N, E) of these feeders. In the same way, it was investigated whether there was a problem of autocorrelation for the proportion of food supplied, as no randomization procedure was used in the field. The test was counted in the “sp” [20,21] and “spdep” [21,22] packages.
A generalized linear mixed model GLMM in the “glmmTMB” package [23] was designed to compare the percentage of food utilization with the food amount that was supplied separately for the percentages of hogweed and sunflower seeds as dependent variables. The distribution of dependent variables was Poisson, and the link function was log. Field visit dates with work on seeds (nominal factor, 1–10) and feeder number (nominal factor, 1–18) were treated as random effects. This model allowed for the author to determine whether the use of each type of seed was complete in all feeders and two parks.
To compare the difference in the proportion of food supplied vs. eaten separately for each type of seed, the non-parametric Kruskal–Wallis test was used. Unlike the above model, the Kruskal–Wallis test took into account variability in the proportions of food types on different dates of the experiment and in different feeders.
To compare whether the proportion of eaten food of one type in individual feeder changed compared to the previously supplied proportion, a non-parametric Wilcoxon signed rank test was calculated in the “PairedData” package [24].
3. Results
The main bird species observed at feeding points were great tit Parus major (dominant), Eurasian blue tit C. caeruleus, Eurasian nuthatch Sitta europaea (Figure 1), and common blackbird Turdus merula. Other birds commonly seen at the feeders include the Eurasian chaffinch Fringilla coelebs, the European greenfinch Chloris chloris, and the bullfinch Pyrrhula pyrrhula. When the seeds fell out of the feeder (such seeds were also counted and included in the food volume and results), pigeons were sometimes feeding on the ground.
A total of 126 episodes of food supplementation were performed in both parks. Of these episodes, 21 cases in Bednarski Park and 24 in Jordan Park also included the addition of hogweed seeds at a volume of, respectively, 10–50 and 30–100 percent of a 0.4-L pot. While sunflower seeds were left after only two episodes, hogweed seeds were left by birds after 26 episodes of feeding with these seeds (Table 1).
The proportion of supplied food was not correlated with supplies in neighbouring feeders (Moran I statistic standard deviate = −3.201, p-value = 0.997) and was independent of the type of food—sunflower and hogweed seeds (Moran I statistic standard deviate = −1.064, p-value = 0.856). The proportion of food eaten in a given feeder was independent of food eaten in the neighbouring feeders (Moran I statistic standard deviate = −3.060, p-value = 0.998) and variable in each type of food (Moran I statistic standard deviate = −0.209, p-value = 0.583).
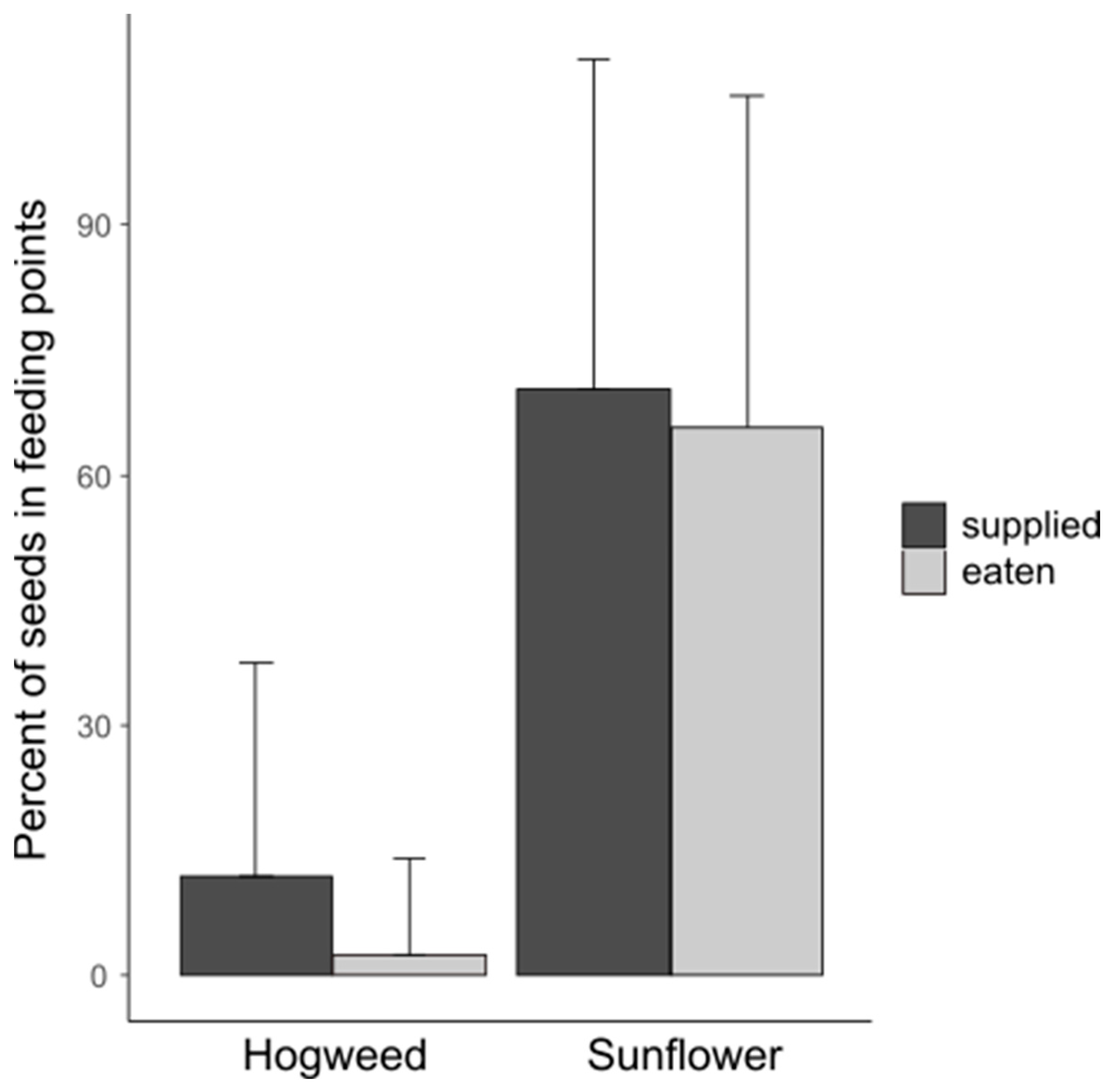
It was confirmed by the GLMM that in the case of both seed types, the supplied food was not fully utilized by the birds, but the eaten food was significantly lower than the previously supplied food (Table 2, Figure 2).
There was a significantly lower proportion of eaten hogweed seeds than the previously supplied one (Kruskal–Wallis chi-squared = 7.347, df = 1, p-value = 0.007) and no differences in these proportions in the case of sunflower seeds (Kruskal–Wallis chi-squared = 1.051, df = 1, p-value = 0.305).
The Wilcoxon paired test showed that the proportion of food eaten compared to the one supplied in specific feeders was lower in the case of hogweed seeds (V = 10, p-value = 0.002), but not in the case of sunflower seeds (V = 262.5, p-value = 0.553).
Hogweed seeds were carefully inspected during their counting, and in approximately half of the inspections involving the counting of these seeds, it was observed that 1–2% of the seeds remaining in the field had traces of damage from the beaks of passerine birds (Figure 3). Hogweed seeds were often found thrown outside the feeder. The first field visits in February with food supply took place in Bednarski Park, and hogweed seeds were left in the feeders from the third day of the experiment, in part of them as a top layer. The next day, hogweed seeds were thrown to the ground from all feeders where the seeds were mixed with sunflowers, despite the presence of 1–2% of seeds disturbed by birds’ beaks on the ground. The birds were not interested in feeders in which sunflower seeds were completely covered by hogweed seeds. In later episodes, hogweed seeds were both thrown out or left in feeders.
4. Discussion
The results of the conducted research showed that in February and March, when the birds were close to finishing their wintering and just before the start of the breeding season, they did not completely consume sunflower seeds, which are their preferred winter food [1], and—compared to relatively intensive feeding on sunflower seeds—they showed only little interest in invasive hogweeds seeds. Based on the previous results of other authors [16], during the experimental period chosen in the presented research, the birds could already use other food sources. This fact could have resulted in the birds being less interested in any artificial winter feeding than might have been expected. However, the incomplete consumption of sunflower seeds could have been influenced by the construction of experimental treatment, i.e., randomness of the dates and locations of the food. Feeding on neighbouring feeders did not affect food consumption in specific locations, which means that food in the feeders could be found by various birds, i.e., both hungry and unneeded birds, which could have resulted in incomplete consumption.
It is worth emphasizing that the weakness of the research conducted and the results obtained is the lack of differentiation of which specific bird species selected the seeds of any plant and threw out those of hogweeds. Their decisions could determine the feeding outcome of the entire flock. On the one hand, seeds lying on the ground became inaccessible to species that do not typically search for food on the ground and, otherwise, could use them if they were accessible in feeders. On the other hand, some birds do not use hanging feeders, so throwing food away may have allowed them to forage, also affecting the results. What is more, based on research conducted on bird foraging on worms in the breeding season, it is known that food deployed in the morning is discovered significantly more often than food deployed in the afternoon [25], as well as potential information about it, should be distributed faster in the morning due to the greater intensity of winter vocalization in such hours [26]. Winter feeders that were filled with food earlier in the day could be detected more effectively, and the preferred food in them could be more likely selected, reflecting the behaviour of only specific early birds. In the presented study, feeders in a given park were visited and refilled in various orders on different days, which allowed for the diversity in bird selection to be taken into account. However, this does not change the fact that to accurately describe the selectivity of all birds participating in the feeding experiment, additional methods should be used, such as recording food consumption using camera traps useful for explaining which species discover novel food sources first, affecting other birds, and successfully avoiding predators [27]. Since the decisions of the urban explorers who first discovered seed feeders certainly influenced the foraging of other birds, this is a limitation of the results discussed, suggesting that they should be treated with caution in interpretation.
The conducted research showed that without controlling the effect of feeder location and feeding date (i.e., according to simple non-parametric tests), there were no differences in the number of seeds eaten and previously supplied in the case of sunflower, while the consumption of hogweed seeds was significantly lower than their assured availability. The conducted research is the first to indicate attempts to consume hogweed seeds and a decisive negative selection of these seeds by birds. This phenomenon makes urban bird explorers similar to birds from more natural populations [14] and confirms the greater validity of multiple-offer experiments than single-offer experiments [15]. Only by offering different food sources in one feeding system at the same time can we learn whether birds make choices. Urban birds can reject food that is less beneficial than sunflower seeds, even if such seeds are twice as large as sunflower seeds (this is the case with hogweed seeds). This is an interesting behavioural observation showing that bird urban explorers likely save time and energy.
At the beginning of the experiment, not knowing the mixed content of the feeders, the birds were not interested in feeders in which sunflower seeds were completely covered by hogweed seeds, although those second seeds were thrown out from feeders with a mix of seeds. This is an unintentional observation in feeders where, according to the methodology, layers of two types of seeds were not mixed or mixed, respectively. The seeds arranged in two layers with hogweeds on top according to the methodology accounted for about half of the hogweed episodes in the experiment, so (although this is speculation) at some stage birds probably started inspecting these seeds. It is difficult to determine whether the most adventurous individuals had finally tried the hogweeds and discovered sunflower seeds underneath them, or whether the attempts for hogweed were because other similar pots contained only sunflowers, so some explorers decided to dig deeper into the same mixed food feeders. The abandonment of feeders with a cover layer of hogweed seeds may confirm that invasive species can influence resource availability and thus suppress or enhance its use by natives [28]. However, the reason might be much simpler, e.g., accidental preparation of feeders with divided layers later, closer to noon than in the morning, when bird explorers notice food faster [25,26]. This is an aspect of foraging experiments where camera traps would be useful [27]. Otherwise, we will never know whether novel food sources are discovered by the best-adapted or risky urban explorers, or whether it is the result of an accident, e.g., spilling seeds outside the feeder.
Invasive hogweed seeds have an intense smell indicating their rich chemical composition. This smell may have distracted birds. Caucasian hogweeds significantly change habitats outside their natural range and have many other properties that can, directly and indirectly, threaten living organisms. They are, for example, known for allelopathic properties consisting of secretion into the soil of chemicals negatively affecting the soil biota of fungi [29], as well as reducing the number and species richness of nematodes [30]. The effect of chemicals present in all parts of the plants known as Caucasian hogweeds on birds is unknown. There is a possibility that bird feeder explorers rejected hogweed seeds as food with at least a specific smell. Regardless of the reason, the avoidance of potentially novel seeds of invaders shows that there is a lack of research exploring the adaptations of native organisms facing this invasion. Similar examples have already included the relationship between fruit-eating birds and invasive plants with fruits used by these birds [31,32]. The meaning of Caucasian hogweeds seeds for birds can be an interesting research direction in the future. Having a huge reproductive capacity, one invasive hogweed produces 5–20 thousand seeds per year in natural conditions [33], making them highly accessible for birds.
5. Conclusions
Bird urban explorers using feeders noted the presence of potentially novel food, the seeds of invasive Caucasian hogweeds, but apart from a small percentage of them trying this food, it was not attractive to them. This shows that birds searching for food in urban areas are still selective, which brings them closer to more natural, non-urban, populations. If the deterrent, in this case, was the smell or the assessment of the hogweed seeds as less attractive than sunflower seeds, birds exploring the feeders would still recognize and reject the unnecessary food. It is also difficult to expect that birds, mostly ignoring invasive hogweed seeds, contribute to their spread in urban areas.
Funding
This research was funded by the statutory fund of the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data can be available upon a reasonable request.
Acknowledgments
The author kindly thank the two anonymous referees for their useful comments that helped in improving the article.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Tryjanowski, P.; Møller, A.P.; Morelli, F.; Indykiewicz, P.; Zduniak, P.; Myczko, Ł. Food preferences by birds using bird-feeders in winter: A large-scale experiment. Avian Res. 2018, 9, 16. [Google Scholar] [CrossRef]
- Jones, D.N.; Reynolds, S.J. Feeding birds in our towns and cities: A global research opportunity. J. Avian Res. 2008, 39, 265–271. [Google Scholar] [CrossRef]
- Robb, G.N.; McDonald, R.A.; Chamberlain, D.E.; Bearhop, S. Food for thought: Supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 2008, 6, 476–484. [Google Scholar] [CrossRef]
- Bonter, D.N.; Zuckerberg, B.; Sedgwick, C.W.; Hochachka, W.M. Daily foraging patterns in free-living birds: Exploring the predation–starvation trade-off. Proc. R. Soc. B 2013, 280, 20123087. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.E.; Meiden, L.N.V.; Shizuka, D. Social partners and temperature jointly affect morning foraging activity of small birds in winter. Behav. Ecol. 2021, 32, 407–415. [Google Scholar] [CrossRef]
- Jones, H.H.; Walters, M.J.; Robinson, S.K. Do similar foragers flock together? Nonbreeding foraging behavior and its impact on mixed-species flocking associations in subtropical region. Auk 2020, 137, ukz079. [Google Scholar] [CrossRef]
- Romanowski, J.; Żmihorski, M. Selection of foraging habitat by grassland birds: Effect of prey abundance or availability? Pol. J. Ecol. 2008, 56, 365–370. [Google Scholar]
- Whittingham, M.J.; Devereux, C.L.; Evans, A.D.; Bradbury, R.B. Altering perceived predation risk and food availability: Management prescriptions to benefit farmland birds on stubble fields. J. Appl. Ecol. 2006, 43, 640–650. [Google Scholar] [CrossRef]
- Bedoya-Perez, M.A.; Carthey, A.J.R.; Mella, V.S.A.; McArthur, C.; Banks, P.B. A practical guide to avoid giving up on giving-up densities. Behav. Ecol. Sociobiol. 2013, 67, 1541–1553. [Google Scholar] [CrossRef]
- Visscher, D.R.; Unger, A.; Grobbelaar, H.; DeWitt, P.D. Bird foraging is influenced by both risk and connectivity in urban parks. J. Urban Ecol. 2018, 4, juy020. [Google Scholar] [CrossRef]
- Atkinson, P.W.; Fuller, R.J.; Vickery, J.A.; Conway, G.J.; Tallowin, J.R.B.; Smith, R.E.N.; Haysom, K.A.; Ings, T.C.; Asteraki, E.J.; Brown, V.K. Influence of agricultural management, sward structure and food resources on grassland field use by birds in lowland England. J. Appl. Ecol. 2005, 42, 932–942. [Google Scholar] [CrossRef]
- Robinson, S.K.; Holmes, R.T. Foraging behavior of forest birds: The relationships among search tactics, diet, and habitat structure. Ecology 1982, 63, 1918–1931. [Google Scholar] [CrossRef]
- Remsen, J.V., Jr.; Robinson, S.K. A classification scheme for foraging behavior of birds in terrestrial habitats. Stud. Avian Biol. 1990, 13, 144–160. [Google Scholar]
- Cueto, V.R.; Marone, L.; de Casenave, J.L. Seed preferences by birds: Effects of the design of feeding-preference experiments. J. Avian Biol. 2001, 32, 275–278. [Google Scholar] [CrossRef]
- Peterson, C.H.; Renaud, P.E. Analysis of feeding preference experiments. Oecologia 1989, 80, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Renner, S.C.; Baur, S.; Possler, A.; Winkler, J.; Kalko, E.K.V.; Bates, P.J.J.; Mello, M.A.R. Food preferences of winter bird communities in different forest types. PLoS ONE 2012, 7, e53121. [Google Scholar] [CrossRef] [PubMed]
- Plummer, K.E.; Bearhop, S.; Leech, D.I.; Chamberlain, D.E.; Blount, J.D. Effects of winter food provisioning on the phenotypes of breeding blue tits. Ecol. Evol. 2018, 8, 5059–5068. [Google Scholar] [CrossRef]
- Grzędzicka, E. Invasion of the giant hogweed and the Sosnowsky’s hogweed as a multidisciplinary problem with unknown future—A review. Earth 2022, 3, 287–312. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 February 2021).
- Pebesma, E.J.; Bivand, R.S. Classes and methods for spatial data in R. R News 2005, 5, 9–13. Available online: https://cran.r-project.org/doc/Rnews/ (accessed on 25 February 2021).
- Bivand, R.S.; Pebesma, E.; Gomez-Rubio, V. Applied Spatial Data Analysis with R, 2nd ed.; Springer: New York, NY, USA, 2013; Available online: https://asdar-book.org/ (accessed on 25 February 2021).
- Bivand, R.S.; Wong, D.W.S. Comparing implementations of global and local indicators of spatial association. TEST 2018, 27, 716–748. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Champely, S. PairedData: Paired Data Analysis, R Package version 1.1.1; University of Lyon: Lyon, France, 2018; Available online: https://CRAN.R-project.org/package=PairedData (accessed on 13 December 2023).
- Farine, D.R.; Lang, S.D.J. The early bird gets the worm: Foraging strategies of wild songbirds lead to the early discovery of food sources. Biol. Lett. 2013, 9, 20130578. [Google Scholar] [CrossRef] [PubMed]
- Hillemann, F.; Cole, E.F.; Keen, S.C.; Sheldon, B.C.; Farine, D.R. Diurnal variation in the production of vocal information about food supports a model of social adjustment in wild songbirds. Proc. R. Soc. B 2019, 286, 20182740. [Google Scholar] [CrossRef] [PubMed]
- Randler, C. Which species discovers novel food sources first? A camera trap study in a natural environment. Avian Res. 2021, 12, 31. [Google Scholar] [CrossRef]
- Didham, R.K.; Ewers, R.M.; Gemmell, N.J. Comment on “Avian extinction and mammalian introductions on oceanic islands”. Science 2005, 307, 1412. [Google Scholar] [CrossRef] [PubMed]
- Glushakova, A.M.; Kachalkin, A.V.; Chernov, I.Y. Soil yeast communities under the aggressive invasion of Sosnowsky’s hogweed (Heracleum sosnowskyi). Eurasian Soil Sci. 2015, 48, 201–207. [Google Scholar] [CrossRef]
- Renčo, M.; Baležentiené, L. An analysis of soil-free-living and plant-parasitic nematode communities in three habitats invaded by Heracleum sosnowskyi in central Lithuania. Biol. Invasions 2015, 17, 1025–1039. [Google Scholar] [CrossRef]
- Díaz Vélez, M.C.; Sérsic, A.N.; Traveset, A.; Paiaro, V. The role of frugivorous birds in fruit removal and seed germination of the invasive alien Cotoneaster franchetii in central Argentina. Austral Ecol. 2018, 43, 558–566. [Google Scholar] [CrossRef]
- Gallinat, A.S.; Primack, R.B.; Lloyd-Evans, T.L. Can invasive species replace native species as a resource for birds under climate change? A case study on bird–fruit interactions. Biol. Conserv. 2020, 241, 108268. [Google Scholar] [CrossRef]
- Perglová, J.; Pergl, J.; Pyšek, P. Flowering phenology and reproductive effort of the invasive alien plant Heracleum mantegazzianum. Preslia 2006, 78, 265–285. [Google Scholar]
Figure 1.
Productive pots hung on bushes and young trees on jute strings with examples of birds feeding on the seeds left inside (author of photographs: E. Grzędzicka).
Figure 1.
Productive pots hung on bushes and young trees on jute strings with examples of birds feeding on the seeds left inside (author of photographs: E. Grzędzicka).

Figure 2.
Comparison of the proportions of eaten and previously supplied seeds separately for hogweed and sunflower during the entire experiment and at all feeding points (i.e., feeders with results of using seeds, including those thrown on the ground under the feeder). The plot shows maximal error bar, i.e., food percent plus standard deviation.
Figure 2.
Comparison of the proportions of eaten and previously supplied seeds separately for hogweed and sunflower during the entire experiment and at all feeding points (i.e., feeders with results of using seeds, including those thrown on the ground under the feeder). The plot shows maximal error bar, i.e., food percent plus standard deviation.

Figure 3.
The examples of hogweed seeds found at feeding points with traces of damage from the beaks of passerine birds (author of photograph: E. Grzędzicka).
Figure 3.
The examples of hogweed seeds found at feeding points with traces of damage from the beaks of passerine birds (author of photograph: E. Grzędzicka).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of bird feeding in all n = 18 feeders, divided into parks and food type. The minimum, maximum, and mean percentages of the volume of seeds in 0.4 L of the pot regarding the food supplemented and then used (or left) by the birds were presented. All descriptive statistics were based on those pots where in a given park either a specific type of seed was supplemented or its use was then expected (pots with left seeds were included in the statistics).
Table 1.
Results of bird feeding in all n = 18 feeders, divided into parks and food type. The minimum, maximum, and mean percentages of the volume of seeds in 0.4 L of the pot regarding the food supplemented and then used (or left) by the birds were presented. All descriptive statistics were based on those pots where in a given park either a specific type of seed was supplemented or its use was then expected (pots with left seeds were included in the statistics).
| City Park in Kraków | Seed Type | Action with Food | Number of Episodes * with Action | Min.–Max. (% of 0.4 L) | Mean (% of 0.4 L) |
|---|---|---|---|---|---|
| Bednarski Park | Hogweed Sunflower | Supplied | 21 | 10–50 | 22.86 |
| Eaten vs. left Supplied Eaten vs. left | 15 vs. 6 70 69 vs. 1 | 0–10 50–100 0–100 | 7.14 94.59 81.08 | ||
| Jordan Park | Hogweed Sunflower | Supplied | 24 | 30–100 | 65.83 |
| Eaten vs. left Supplied Eaten vs. left | 4 vs. 20 56 55 vs. 1 | 0–40 10–100 0–100 | 11.67 72.58 68.71 |
* Episode—a single food supplement in a given feeder; there were seven such episodes at each feeder on different days.
Table 2.
Results of GLMM models comparing the proportions of food eaten and previously supplied for food types (hogweed, sunflower). In all cases, df = 1.
Table 2.
Results of GLMM models comparing the proportions of food eaten and previously supplied for food types (hogweed, sunflower). In all cases, df = 1.
| Food Type | Model Predictor | Estimate ± se | z-Value | p-Value |
|---|---|---|---|---|
| Hogweed | Intercept Action with food * | −4.306 ± 2.300 | −1.872 | 0.061 |
| 1.609 ± 0.079 | 20.252 | <0.001 | ||
| Sunflower | Intercept Action with food * | 4.138 ± 0.098 | 41.950 | <0.001 |
| 0.069 ± 0.019 | 3.640 | <0.001 |
* Action with food: supplied or eaten.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grzędzicka, E. Bird Feeder Explorers Are Not Attracted by the Seeds of Invasive Weeds in Winter. Diversity 2024, 16, 81. https://doi.org/10.3390/d16020081
AMA Style
Grzędzicka E. Bird Feeder Explorers Are Not Attracted by the Seeds of Invasive Weeds in Winter. Diversity. 2024; 16(2):81. https://doi.org/10.3390/d16020081
Chicago/Turabian StyleGrzędzicka, Emilia. 2024. "Bird Feeder Explorers Are Not Attracted by the Seeds of Invasive Weeds in Winter" Diversity 16, no. 2: 81. https://doi.org/10.3390/d16020081
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.