
Coping with Stress as a Prelude of Naturalization: Different Responses of Lagarosiphon Species to Water Trophy

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Target Species
2.2. Experimental Setup
2.3. Environmental Parameters
2.4. Experimental Treatments
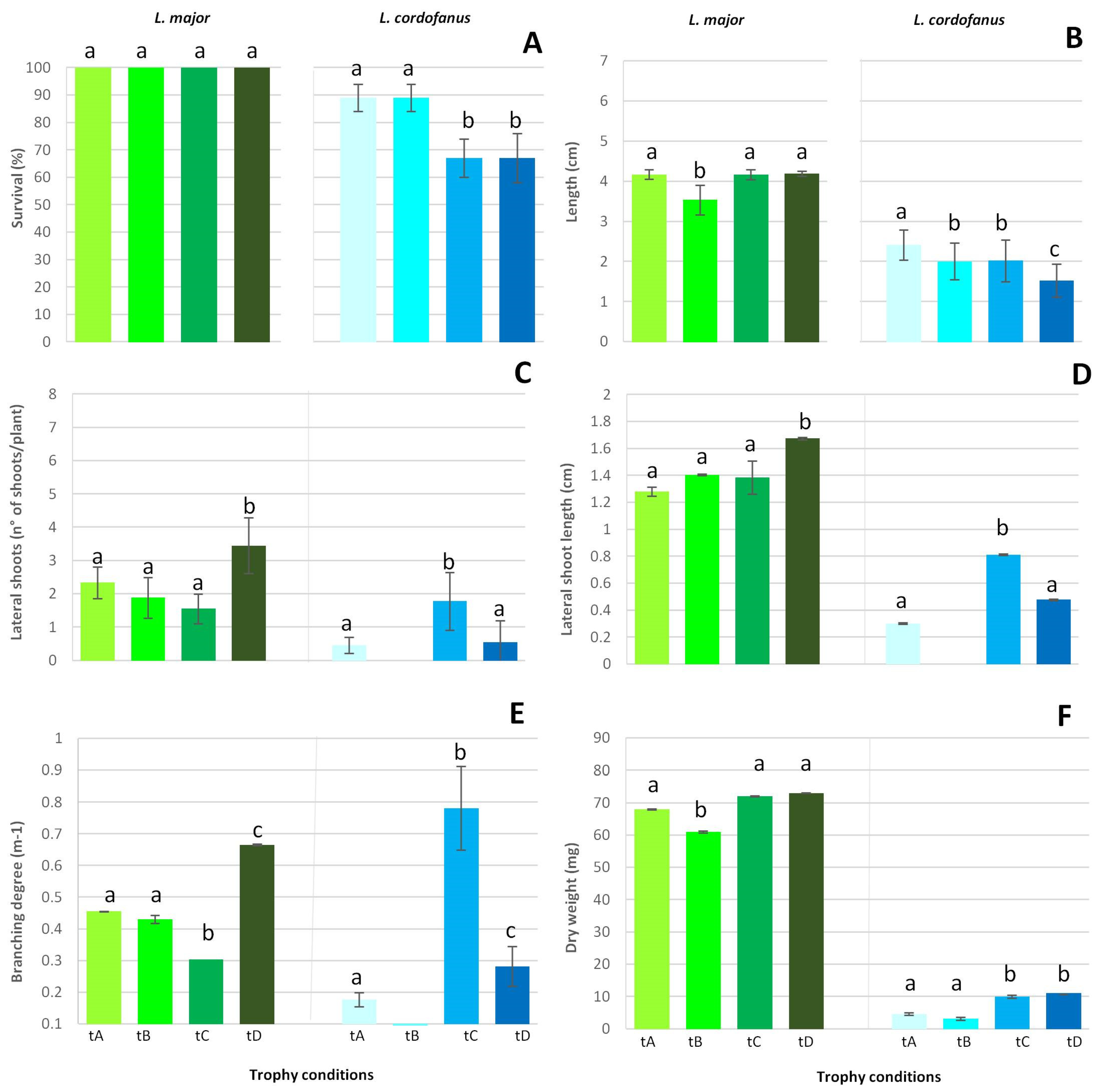
2.5. Functional Traits
- Plant survival percentage;
- Main shoot length (cm);
- Final dry weight (mg);
- Number of lateral shoots (n° of shoots per plant);
- Lateral shoot length (cm);
- Branching degree (m−1);
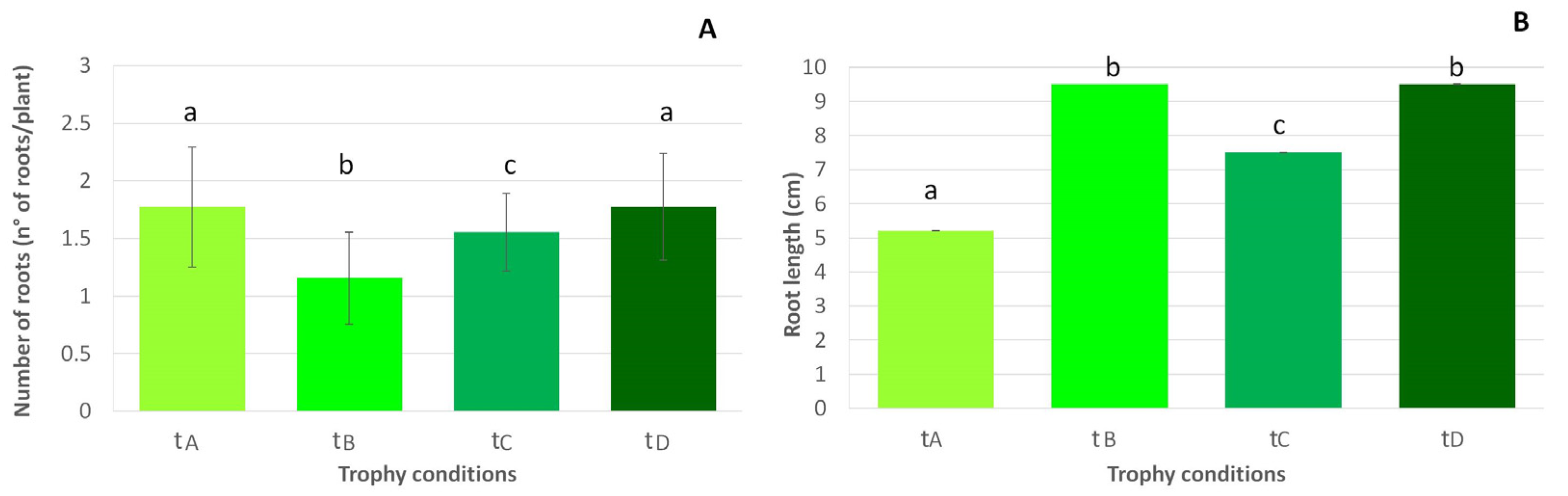
- Number of roots (n° of roots per plant);
- Root length (cm).
2.6. Periphyton and Microalgal Mucilage Cover
2.7. Statistical Analysis
3. Results
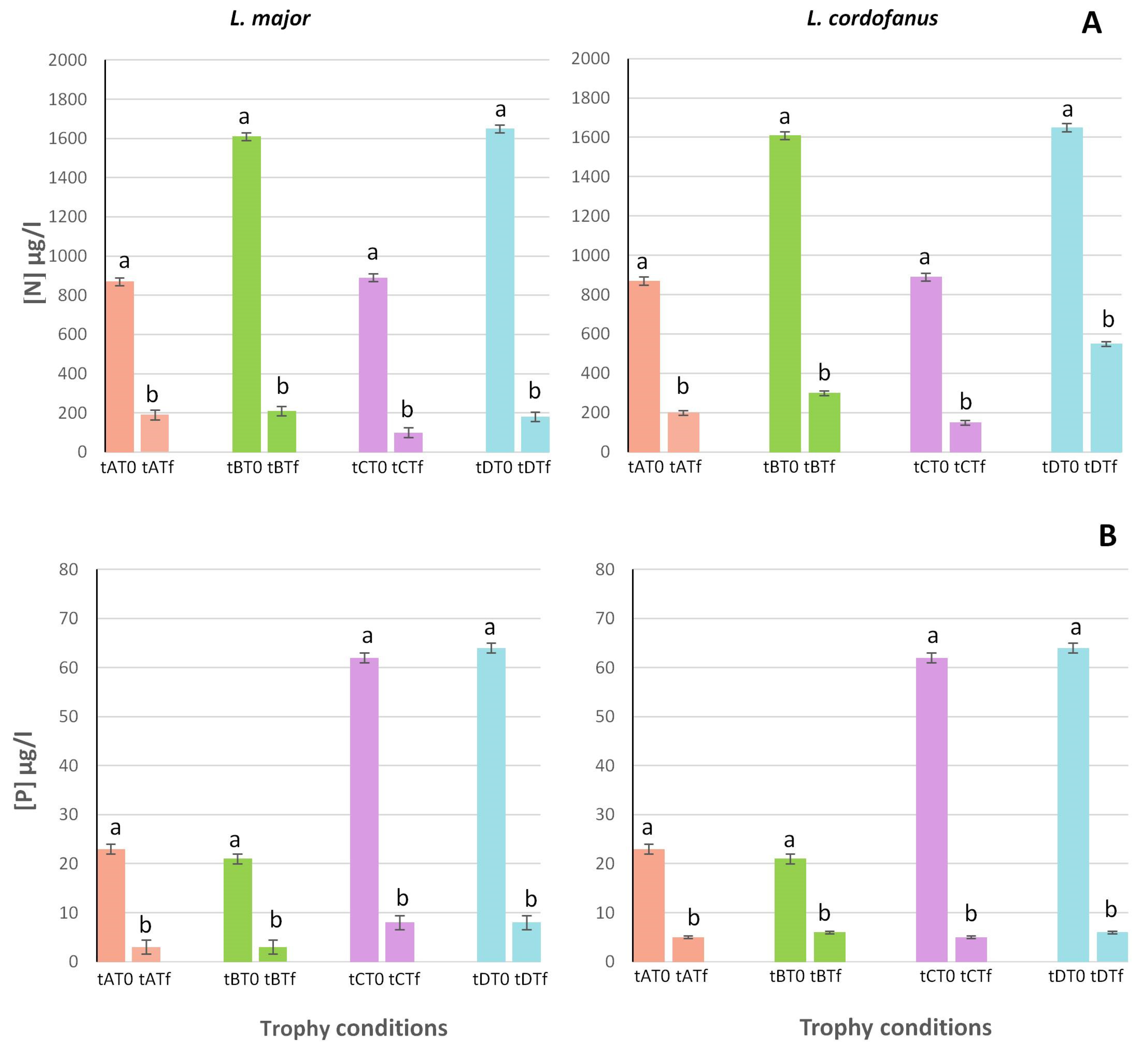
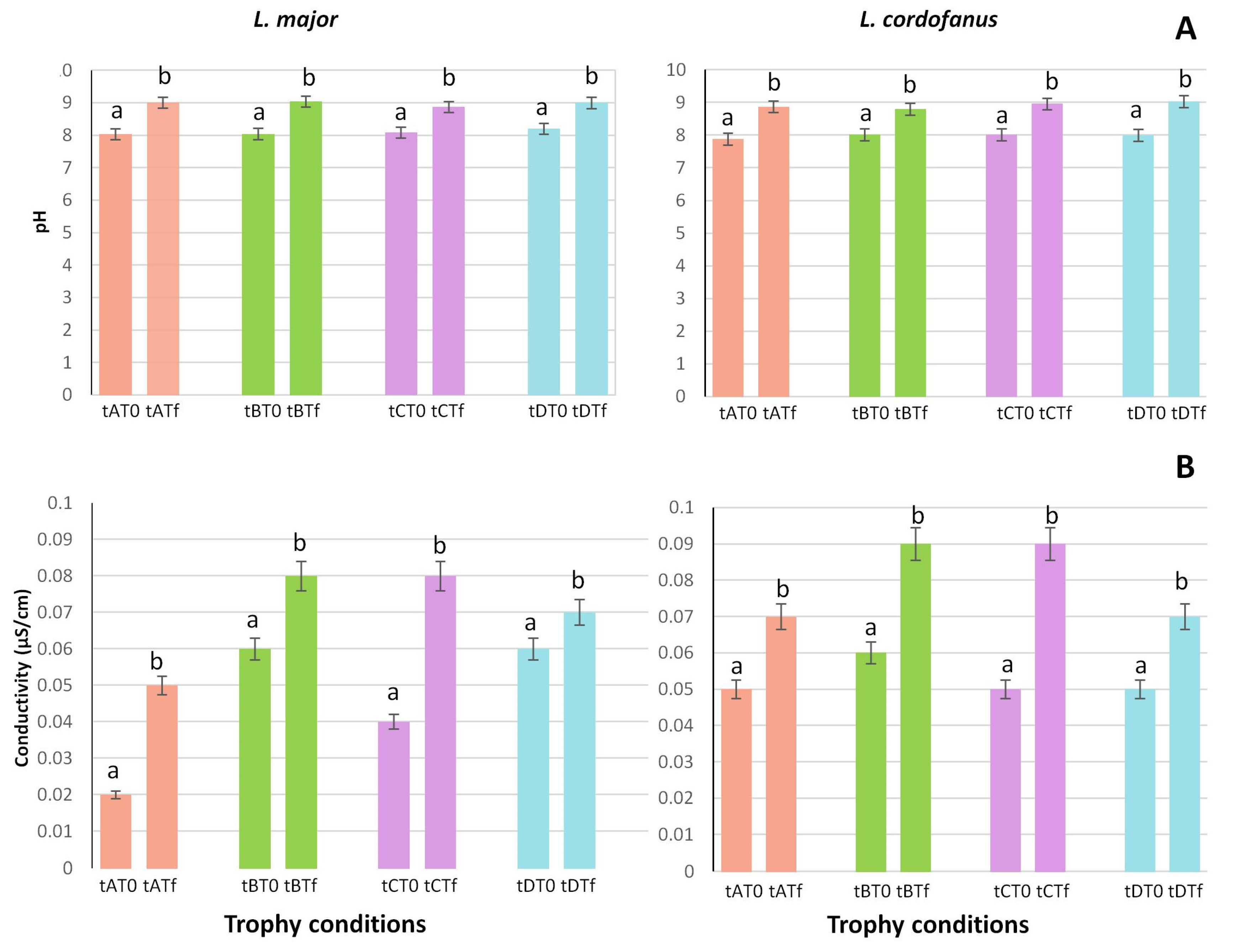
3.1. Environmental Parameters
3.2. Functional Traits
3.3. Periphyton and Microalgal Mucilage Cover
4. Discussion
4.1. Species Response to Nutrient Enrichment
4.2. Strategic Investment into Shoots and Roots
4.3. Factors Influencing the Growth and Survival
4.4. Change in Environmental Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turbelin, A.J.; Malamud, B.D.; Francis, R.A. Mapping the global state of invasive alien species: Patterns of invasion and policy responses. Glob. Ecol. Biogeogr. 2017, 26, 78–92. [Google Scholar] [CrossRef]
- Hulme, P.E.; Brundu, G.; Carboni, M.; Dehnen-Schmutz, K.; Dullinger, S.; Early, R.; Essl, F.; González-Moreno, P.; Groom, Q.J.; Kueffer, C.; et al. Integrating invasive species policies across ornamental horticulture supply chains to prevent plant invasions. J. Appl. Ecol. 2017, 55, 92–98. [Google Scholar] [CrossRef]
- Brunel, S. Pathway analysis: Aquatic plants imported in 10 EPPO countries. EPPO Bull. 2009, 39, 201–213. [Google Scholar] [CrossRef]
- Caffrey, J.M.; Acevedo, S. Lagarosiphon major in Lough Corrib—Management options. In Fish Stocks and Their Environment; Moriary, C., Rossell, R., Gargan, P., Eds.; Institute of Fisheries Management: Westport, Ireland, 2008; pp. 85–97. [Google Scholar]
- Caffrey, J.M.; Millane, M.; Evers, S.; Moran, H.; Butler, M.A. Novel approach to aquatic weed control and habitat restoration using biodegradable jute matting. Aquat. Invasions 2010, 5, 123–129. [Google Scholar] [CrossRef]
- Baars, J.R.; Keenan, E.A.; O’Callaghan, P.; Caffrey, J.M. Changes to the invertebrate fauna of littoral habitats induced by the alien invasive species Lagarosiphon major (Hydrocharitaceae). In Research and Control Programme for Lagarosiphon major in Lough Corrib; Central Fisheries Board: Dublin, Ireland, 2009; p. 159. [Google Scholar]
- Bickel, T.O. Lagarosiphon major. In A Handbook of Global Freshwater Invasive Species; Francis, R.A., Ed.; Routledge: London, UK; New York, NY, USA, 2012. [Google Scholar]
- Matthews, J.; Beringen, R.; Collas, F.P.L.; Koopman, K.R.; Odé, B.; Pot, R.; Sparrius, L.B.; van Valkenburg, J.L.C.H.; Verbrugge, L.N.H.; Leuven, R.S.E.W. Knowledge Document for Risk Analysis of the Non-Native Curly Waterweed (Lagarosiphon major) in The Netherlands; Reports Environmental Science, 414; Department of Environmental Science: Nijmegen, The Netherlands, 2012. [Google Scholar]
- Redekop, P.; Hofstra, D.; Hussner, A. Elodea canadensis shows a higher dispersal capacity via fragmentation than Egeria densa and Lagarosiphon major. Aquat. Bot. 2016, 130, 45–49. [Google Scholar] [CrossRef]
- Turbelin, A.J.; Diagne, C.; Hudgins, E.J.; Moodley, D.; Kourantidou, M.; Novoa, A.; Haubrock, P.J.; Bernery, C.; Gozlan, R.E.; Francis, R.A.; et al. Introduction pathways of economically costly invasive alien species. Biol. Invasions 2022, 24, 2061–2079. [Google Scholar] [CrossRef]
- McGregor, P.G.; Gourlay, H. Assessing the Prospects for Biological Control of Lagarosiphon (Lagarosiphon major (Hydrocharitaceae)); Department of Conservation: Wellington, New Zealand, 2002; p. 14. [Google Scholar]
- Caffrey, J.; Millane, M.; Evers, S.; Moran, H. Management of Lagarosiphon major (Ridley) Moss in Lough Corrib—A review. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 2011; pp. 205–212. [Google Scholar]
- Lansdown, R.V. Lagarosiphon cordofanus. In The IUCN Red List of Threatened Species; IUCN Global Species Programme Red List Unit: Cambridge, UK, 2019; p. e.T185529A120203981. [Google Scholar]
- Migliorini, D.; Ghelardini, L.; Tondini, E.; Luchi, N.; Santini, A. The potential of symptomless potted plants for carrying invasive soilborne plant pathogens. Divers. Distrib. 2015, 21, 1218–1229. [Google Scholar] [CrossRef]
- Branquart, E.; Brundu, G.; Buholzer, S.; Chapman, D.; Ehret, P.; Fried, G.; Starfinger, U.; van Valkenburg, J.; Tanner, R. A prioritization process for invasive alien plant species incorporating the requirements of EU Regulation no. 1143/2014. EPPO Bull. 2016, 46, 603–617. [Google Scholar] [CrossRef]
- Roy, H.E.; Bacher, S.; Essl, F.; Adriaens, T.; Aldridge, D.C.; Bishop, J.D.D.; Blackburn, T.M.; Branquart, E.; Brodie, J.; Carboneras, C.; et al. Developing a list of invasive alien species likely to threaten biodiversity and ecosystems in the European Union. Glob. Chang. Biol. 2019, 25, 1032–1048. [Google Scholar] [CrossRef]
- Leung, B.; Roura-Pascual, N.; Bacher, S.; Heikkilä, J.; Brotons, L.; Burgman, M.A.; Dehnen-Schmutz, K.; Essl, F.; Hulme, P.E.; Richardson, D.M.; et al. TEASIng apart alien species risk assessments: A framework for best practices. Ecol. Lett. 2012, 15, 1475–1493. [Google Scholar] [CrossRef]
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 52–66. [Google Scholar] [CrossRef]
- Essl, F.; Lenzner, B.; Bacher, S.; Bailey, S.; Capinha, C.; Daehler, C.; Dullinger, S.; Genovesi, P.; Hui, C.; Hulme, P.E.; et al. Drivers of future alien species impacts: An expert-based assessment. Glob. Chang. Biol. 2020, 26, 4880–4893. [Google Scholar] [CrossRef] [PubMed]
- Preston, D.L.; Hedman, H.D.; Johnson, P.T.J. Nutrient availability and invasive fish jointly drive community dynamics in an experimental aquatic system. Ecosphere 2018, 9, e02153. [Google Scholar] [CrossRef]
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef]
- Paerl, H.W. Controlling eutrophication along the freshwater–marine continuum: Dual nutrient (N and P) reductions are essential. Estuaries Coasts 2009, 32, 593–601. [Google Scholar] [CrossRef]
- Egertson, C.J.; Kopaska, J.A.; Downing, J.A. A Century of Change in Macrophyte Abundance and Composition in Response to Agricultural Eutrophication. Hydrobiologia 2004, 524, 145–156. [Google Scholar] [CrossRef]
- Phillips, G.L.; Eminson, D.; Moss, B. A mechanism to account for macrophyte decline in progressively eutrophicated freshwaters. Aquat. Bot. 1978, 4, 103–126. [Google Scholar] [CrossRef]
- Bolpagni, R. Towards global dominance of invasive alien plants in freshwater ecosystems: The dawn of the Exocene? Hydrobiologia 2021, 848, 2259–2279. [Google Scholar] [CrossRef]
- Rattray, M.R.; Howard-Williams, C.; Brown, J.M.A. Rates of early growth of propagules of Lagarosiphon major and Myriophyllum triphyllum in lakes of differing trophic status. N. Z. J. Mar. Freshw. Res. 1994, 28, 235–241. [Google Scholar] [CrossRef]
- Martin, G.D.; Coetzee, J.A. Competition between two aquatic macrophytes, Lagarosiphon major (Ridley) Moss (Hydrocharitaceae) and Myriophyllum spicatum Linnaeus (Haloragaceae) as influenced by substrate sediment and nutrients. Aquat. Bot. 2014, 114, 1–11. [Google Scholar] [CrossRef]
- Cholo, F.; Foden, W. Lagarosiphon major. In The IUCN Red List of Threatened Species; IUCN Global Species Programme Red List Unit: Cambridge, UK, 2010; p. e.T185287A8382399. [Google Scholar]
- Riis, T.; Olesen, B.; Clayton, J.S.; Lambertini, C.; Brix, H.; Sorrell, B.K. Growth and morphology in relation to temperature and light availability during the establishment of three invasive aquatic plant species. Aquat. Bot. 2012, 102, 56–64. [Google Scholar] [CrossRef]
- Stiers, I.; Njambuya, J.; Triest, L. Competitive abilities of invasive Lagarosiphon major and native Ceratophyllum demersum in monocultures and mixed cultures in relation to experimental sediment dredging. Aquat. Bot. 2011, 95, 161–166. [Google Scholar] [CrossRef]
- Heidbüchel, P.; Hussner, A. Fragment type and water depth determine the regeneration and colonization success of submerged aquatic plants. Aquat. Sci. 2019, 81, 6. [Google Scholar] [CrossRef]
- Coughlan, N.E.; Armstrong, F.; Baker-Arney, C.; Crane, K.; Cuthbert, R.N.; Jansen, M.A.; Kregting, L.; Vong, G.Y.; Dick, J.T. Retention of viability by fragmented invasive Crassula helmsii, Elodea canadensis and Lagarosiphon major. River Res. Appl. 2022, 38, 1356–1361. [Google Scholar] [CrossRef]
- Hussner, A. Information on Measures and Related Costs in Relation to the Species Included on the Union List: Lagarosiphon major; Technical Note Prepared by IUCN for the European Commission; IUCN: Gland, Switzerland, 2019.
- Symoens, J.J.; Triest, L. Monograph of the african genus Lagarosiphon Harvey (Hydrocharitaceae). Bull. Du Jard. Bot. Natl. De Belg./Bull. Van De Natl. Plantentuin Van Belg. 1983, 53, 441–488. [Google Scholar] [CrossRef]
- Ngumbau, V.M.; Luke, Q.; Nyange, M.; Wanga, V.O.; Watuma, B.M.; Mbuni, Y.M.; Munyao, J.N.; Oulo, M.A.; Mkala, E.M.; Kipkoech, S.; et al. An annotated checklist of the coastal forests of Kenya, East Africa. PhytoKeys 2020, 147, 1–191. [Google Scholar] [CrossRef]
- Premazzi, G.; Chiaudani, G. Ecological Quality of Surface Waters: Quality Assessment Schemes for European Community Lakes; European Commission: Brussels, Luxembourg, 1992; Volume 14563.
- Monitoraggio PFAS in Lombardia. Available online: https://www.arpalombardia.it/Pages/Acque-Superficiali/Rapporti-Annuali.aspx (accessed on 20 February 2023).
- Scibona, A.; Nizzoli, D.; Hupfer, M.; Valerio, G.; Pilotti, M.; Viaroli, P. Decoupling of silica, nitrogen and phosphorus cycling in a meromictic subalpine lake (Lake Iseo, Italy). Biogeochemistry 2022, 159, 371–392. [Google Scholar] [CrossRef]
- Bolpagni, R.; Cerabolini, B.E.L. Habitat Acquatici in Lombardia: Aggiornamento delle Conoscenze e Proposte per un Monitoraggio Integrato; Università degli Studi dell’Insubria—Fondazione Lombardia per l’Ambiente, Osservatorio Regionale per la Biodiversità di Regione Lombardia: Milan, Italy, 2016. [Google Scholar]
- Bolpagni, R.; Azzella, M.M.; Agostinelli, C.; Beghi, A.; Bettoni, E.; Brusa, G.; De Molli, C.; Formenti, R.; Galimberti, F.; Cerabolini, B.E. Integrating the Water Framework Directive into the Habitats Directive: Analysis of distribution patterns of lacustrine EU habitats in lakes of Lombardy (northern Italy). J. Limnol. 2017, 76, 75–83. [Google Scholar] [CrossRef]
- Ciutti, F.; Cappelletti, C. Invasioni biologiche: Il caso del Lago di Garda. Biol. Ambient. 2017, 31, 59–164. [Google Scholar]
- Eugelink, A.H. Phosphorus uptake and active growth of E. canadensis Michx. and Elodea nuttallii (Planch.) St. John. Water Sci. Technol. 1998, 37, 59–65. [Google Scholar] [CrossRef]
- Barrat-Segretain, M.H. Growth of E. canadensis and Elodea nuttallii in mono-cultures and mixture under different light and nutrient conditions. Arch. Hydrobiol. 2004, 161, 133–144. [Google Scholar] [CrossRef]
- Zehnsdorf, A.; Hussner, A.; Eismann, F.; Rönicke, H.; Melzer, A. Management options of invasive Elodea nuttallii and Elodea canadensis. Limnologica 2015, 51, 110–117. [Google Scholar] [CrossRef]
- Thiébaut, G. Phosphorus and aquatic plants. In The Ecophysiology of Plant-Phosphorus Interactions; White, P.J., Hammond, J.P., Eds.; Springer Science & Business Media: Dordrecht, The Netherlands, 2008; Volume 7, pp. 31–49. [Google Scholar]
- Baldy, V.; Thiebaut, G.; Fernandez, C.; Sagova-Mareckova, M.; Korboulewsky, N.; Monnier, Y.; Perez, T.; Tremolieres, M. Experimental assessment of the water quality influence on the phosphorus uptake of an invasive aquatic plant: Biological responses throughout its phenological stage. PLoS ONE 2015, 10, e0118844. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. Interactions between Nitrogen and Phosphorus metabolism. In Annual Plant Reviews; Plaxton, W.C., Lambers, H., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; Volume 48. [Google Scholar]
- Rao, Q.; Su, H.; Deng, X.; Xia, W.; Wang, L.; Cui, W.; Ruan, L.; Chen, J.; Xie, P. Carbon, nitrogen, and phosphorus allocation strategy among organs in submerged macrophytes is altered by eutrophication. Front. Plant Sci. 2020, 11, 524450. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, H.Z.; Li, Y.; Shao, J.C.; Liang, X.M.; Jeppesen, E.; Wang, H.J. Effects of high nitrogen concentrations on the growth of submersed macrophytes at moderate phosphorus concentrations. Water Res. 2015, 83, 385–395. [Google Scholar] [CrossRef]
- Gonzalez Sagrario, M.A.; Jeppesen, E.; Gomà, J.; Søndergaard, M.; Jensen, J.P.; Lauridsen, T.; Landkildehus, F. Does high nitrogen loading prevent clear-water conditions in shallow lakes at moderately high phosphorus concentrations? Freshw. Biol. 2005, 50, 27–41. [Google Scholar] [CrossRef]
- Moss, B.; Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Liu, Z. Nitrogen, macrophytes, shallow lakes and nutrient limitation: Resolution of a current controversy? Hydrobiologia 2013, 710, 3–21. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Response of aquatic plants to abiotic factors: A review. Aquat. Sci. 2011, 73, 1–14. [Google Scholar] [CrossRef]
- Olsen, S.; Chan, F.; Li, W.; Zhao, S.; Søndergaard, M.; Jeppesen, E. Strong impact of nitrogen loading on submerged macrophytes and algae: A long-term mesocosm experiment in a shallow Chinese lake. Freshw. Biol. 2015, 60, 1525–1536. [Google Scholar] [CrossRef]
- Hussner, A.; Smith, R.; Mettler-Altmann, T.; Hill, M.P.; Coetzee, J. Simulated global increases in atmospheric CO2 alter the tissue composition, but not the growth of some submerged aquatic plant bicarbonate users growing in DIC rich waters. Aquat. Bot. 2019, 153, 44–50. [Google Scholar] [CrossRef]
- James, C.S.; Eaton, J.W.; Hardwick, K. Responses of three invasive aquatic macrophytes to nutrient enrichment do not explain their observed field displacements. Aquat. Bot. 2006, 84, 347–353. [Google Scholar] [CrossRef]
- Garbey, C.; Murphy, K.J.; Thiébaut, G.; Muller, S. Variation in P-content in aquatic plant tissues offers an efficient tool for determining plant growth strategies along a resource gradient. Freshw. Biol. 2004, 49, 346–356. [Google Scholar] [CrossRef]
- Wersal, R.M.; Madsen, J.D. Influences of water column nutrient loading on growth characteristics of the invasive aquatic macrophyte Myriophyllum aquaticum (Vell.). Verdc. Hydrobiol. 2011, 665, 93–105. [Google Scholar] [CrossRef]
- Madsen, T.V.; Cedergreen, N. Sources of nutrients to rooted submerged macrophytes growing in a nutrient-rich stream. Freshw. Biol. 2002, 47, 283–291. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Y.; Han, R.; Song, K.; Zhou, X.; Wang, G.; Wang, Q. Eutrophication triggers the shift of nutrient absorption pathway of submerged macrophytes: Implications for the phytoremediation of eutrophic waters. J. Environ. Manag. 2019, 239, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, K.; Heidbüchel, P.; Hussner, A. Effects of water nutrients on regeneration capacity of submerged aquatic plant fragments. Ann. Limnol.-Int. J. Lim. 2014, 50, 155–162. [Google Scholar] [CrossRef]
- Levi, P.S.; Riis, T.; Alnøe, A.B.; Peipoch, M.; Maetzke, K.; Bruus, C.; Baattrup-Pedersen, A. Macrophyte complexity controls nutrient uptake in lowland streams. Ecosystems 2015, 18, 914–931. [Google Scholar] [CrossRef]
- Heidbüchel, P.; Sachs, M.; Stanik, N.; Hussner, A. Species-specific fragmentation rate and colonization potential partly explain the successful spread of aquatic plants in lowland streams. Hydrobiologia 2019, 843, 107–123. [Google Scholar] [CrossRef]
- Dai, Y.; Wu, J.; Ma, X.; Zhong, F.; Cui, N.; Cheng, S. Increasing phytoplankton-available phosphorus and inhibition of macrophyte on phytoplankton bloom. Sci. Total Environ. 2017, 579, 871–880. [Google Scholar] [CrossRef]
- Søndergaard, M.; Lauridsen, T.L.; Johansson, L.S.; Jeppesen, E. Nitrogen or phosphorus limitation in lakes and its impact on phytoplankton biomass and submerged macrophyte cover. Hydrobiologia 2017, 795, 35–48. [Google Scholar] [CrossRef]
- Qin, B.Q.; Gao, G.; Zhu, G.W.; Zhang, Y.L.; Song, Y.Z.; Tang, X.M.; Xu, H.; Deng, J.M. Lake eutrophication and its ecosystem response. Chin. Sci. Bull. 2013, 58, 961–970. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, H.; Zhang, J.; Yu, J.; Xie, P.; Chen, J. Physiological differences between free-floating and periphytic filamentous algae, and specific submerged macrophytes induce proliferation of filamentous algae: A novel implication for lake restoration. Chemosphere 2020, 239, 124702. [Google Scholar] [CrossRef] [PubMed]
- Hussner, A.; Heidbüchel, P.; Coetzee, J.; Gross, E.M. From introduction to nuisance growth: A review of traits of alien aquatic plants which contribute to their invasiveness. Hydrobiologia 2021, 848, 2119–2151. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Naturalization of introduced plants: Ecological drivers of biogeographical patterns. New Phytol. 2012, 196, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Zenni, R.D.; Nuñez, M.A. The elephant in the room: The role of failed invasions in understanding invasion biology. Oikos 2013, 122, 801–815. [Google Scholar] [CrossRef]
- Pedersen, O.; Colmer, T.D.; Sand-Jensen, K. Underwater photosynthesis of submerged plants–recent advances and methods. Front. Plant Sci. 2013, 4, 140. [Google Scholar] [CrossRef]
- Hussner, A.; Mettler-Altmann, T.; Weber, A.P.; Sand-Jensen, K. Acclimation of photosynthesis to supersaturated CO2 in aquatic plant bicarbonate users. Freshw. Biol. 2016, 61, 1720–1732. [Google Scholar] [CrossRef]
- Prins, H.B.A.; Elzenga, J.T.M. Bicarbonate utilization: Function and mechanism. Aquat. Bot. 1989, 34, 59–83. [Google Scholar] [CrossRef]
- Underwood, A. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Gerald, B. A brief review of independent, dependent and one sample t-test. Int. J. Appl. Math. Theor. Phys. 2018, 4, 50–54. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total N (µg/L) | Total P (µg/L) | Level of Trophy for Total N and Total P |
|---|---|---|---|
| A | 870 | 21 | Oligo-/mesotrophy |
| B | 1610 | 21 | Hyper-/mesotrophy |
| C | 890 | 62 | Oligo-/eutrophy |
| D | 1680 | 64 | Hyper-/eutrophy |
| Treatment | Periphyton and Microalgal Mucilage Cover | |
|---|---|---|
| L. cordofanus | L. major | |
| A | 30% | 22% |
| B | 48% | 45% |
| C | 60% | 50% |
| D | 65% | 60% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montagnani, C.; Caronni, S.; Quaglini, L.A.; Sebesta, N.; Gentili, R.; Citterio, S. Coping with Stress as a Prelude of Naturalization: Different Responses of Lagarosiphon Species to Water Trophy. Diversity 2023, 15, 693. https://doi.org/10.3390/d15050693
Montagnani C, Caronni S, Quaglini LA, Sebesta N, Gentili R, Citterio S. Coping with Stress as a Prelude of Naturalization: Different Responses of Lagarosiphon Species to Water Trophy. Diversity. 2023; 15(5):693. https://doi.org/10.3390/d15050693
Chicago/Turabian StyleMontagnani, Chiara, Sarah Caronni, Lara Assunta Quaglini, Nicole Sebesta, Rodolfo Gentili, and Sandra Citterio. 2023. "Coping with Stress as a Prelude of Naturalization: Different Responses of Lagarosiphon Species to Water Trophy" Diversity 15, no. 5: 693. https://doi.org/10.3390/d15050693