
Two New Edible Lyophyllum Species from Tibetan Areas, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Morphological Studies
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Sequence Alignment and Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Specimen Number | Origin | ITS | Reference |
|---|---|---|---|---|
| Calocybe gambosa | HC78/64 | Switzerland | AF357027 | [7] |
| C. carnea | CBS552.50 | Switzerland | AF357028 | [7] |
| C. persicolor | HC80/99 | Switzerland | AF357026 | [7] |
| Hypsizygus marmoreus | V.1611 | Germany | AJ494834 | [11] |
| H. marmoreus | V.3133 | China | FJ609271 | [11] |
| H. marmoreus | HMW2 | Malaysia | HM561971 | [11] |
| H. marmoreus | HZND-1 | China | JX046028 | [11] |
| H. marmoreus | 1-1 | Korea | KF192813 | [11] |
| H. tessulatus | AFTOL-ID 1898 | USA | DQ917653 | [11] |
| H. tessulatus | L2 | China | FJ467372 | [11] |
| H. ulmarius | CBS 286.77 | Korea | AY265850 | [29] |
| H. ulmarius | DUKE-JM/HW | USA | EF421105 | [30] |
| L. ambustum | CBS452.87 | Switzerland | AF357057 | [7] |
| L. anthracophilum | HC79/132 | Switzerland | AF357055 | [7] |
| L. atratum | CBS709.87 | Switzerland | AF357053 | [7] |
| L. atrofuscum | HMJAU63461 | China | OP605493 | [16] |
| L. atrofuscum | HMJAU63456 * | China | OP605494 | [16] |
| L. caerulescens | HC80-140 | Switzerland | AF357052 | [7] |
| L. caerulescens | V.15759 | USA | JF908339 | [11] |
| L. crassifolium | V.5077 | Italy | JF908331 | [11] |
| L. decastes | dd08054 | China | FJ810160 | [11] |
| L. decastes | Ld418 | China | HM119485 | [11] |
| L. deqinense | YAASM6949 * | China | OQ418117 | [18] |
| L. deqinense | YAASM6948 | China | OQ418116 | [18] |
| L. deliberatum | V.15032 | Slovenia | JF908338 | [11] |
| L. favrei | BSI94cp2 | Switzerland | AF357035 | [7] |
| L. favrei | V.6334 | Italy | JF908333 | [11] |
| L. fumosum | SJ02/006 | Sweden | HM572539 | [11] |
| L. fumosum | LAS00/144 | Sweden | HM572541 | [11] |
| L. fumosum | V.16077 | Italy | JF908340 | [11] |
| L. fumosum | LfumNlf24 | Japan | JN983977 | [11] |
| L. fumosum | L2010512371 | China | JX966310 | [11] |
| L. fumosum | YAASM6215 | China | ON681708 | [17] |
| L. fumosum | YAASM6340 | China | ON681709 | [17] |
| L. gangraenosum | V.12332 | Italy | JF908335 | [11] |
| L. heimogu | L3026 * | China | KY434100 | This study |
| L. heimogu | L3033 | China | KY434101 | This study |
| L. heimogu | L3035 | China | KY434102 | This study |
| L. infumatum | V.10152 | Italy | JF908334 | [11] |
| L. leucophaeatum | Hae251.97 | Switzerland | AF357032 | [7] |
| L. littoralis | CA20091210 | Italy | JX280410 | [11] |
| L. loricatum | V.13175 | USA | JF908336 | [11] |
| L. loricatum | CA20090202.03 | Italy | JX280406 | [11] |
| L. loricatum | 01.12.09 | Italy | JX280407 | [11] |
| L. moncalvoanum | PDD 96328 * | New Zealand | NR_137615 | [2] |
| L. moncalvoanum | PDD 102581 | New Zealand | KJ461912 | [2] |
| L. ochraceum | BSI94.cp1 | Switzerland | AF357033 | [7] |
| L. ochraceum | V.537 | Italy | JF908329 | [11] |
| L. rhombisporum | L1762* | China | JX966307 | [11] |
| L. rhombisporum | L2082 | China | JX966308 | [11] |
| L. semitale | HC85/13 | Switzerland | AF357049 | [7] |
| L. semitale | EL187-09 | Sweden | HM572552 | [8] |
| L. shimeji | Olsen821006 | Sweden | HM572530 | [8] |
| L. shimeji | NZ4Q88 | New Zealand | JN983985 | [11] |
| L. shimeji | L2010512377 | China | JX966311 | [11] |
| L. sp. | PBM 2688 | USA | DQ182502 | [11] |
| L. sp. | SB102 | China | FJ687273 | [11] |
| L. sp. | Aase811014 | Sweden | HM572550 | [8] |
| L. sp. | TO-2011 | Italy | JF908337 | [11] |
| L. sp. | JN001 | China | FJ687270 | [11] |
| L. sp. | O73586 | Netherlands | GU234137 | [11] |
| L. sp. | Cultivar Jpn | Sweden | HM572551 | [8] |
| L. sp. | SL-2013 | China | JX966308 | [11] |
| L. sykosporum | IFO30978 | Switzerland | AF357050 | [7] |
| L. subalpinarum | HMJAU63447 * | China | OP605490 | [16] |
| L. subalpinarum | HMJAU63453 | China | OP605491 | [16] |
| L. subdecastes | HMJAU63470 | China | OP605488 | [16] |
| L. subdecastes | HMJAU63467 * | China | OP605489 | [16] |
| L. turcicum | KATO-2971 * | Turkey | KJ158159 | [11] |
| L. yiqunyang | L4206 | China | KY434104 | This study |
| L. yiqunyang | L2989 * | China | KY434103 | This study |
| Tephrocybe boudier | BSI96/84 | USA | DQ825427 | [11] |
3. Result
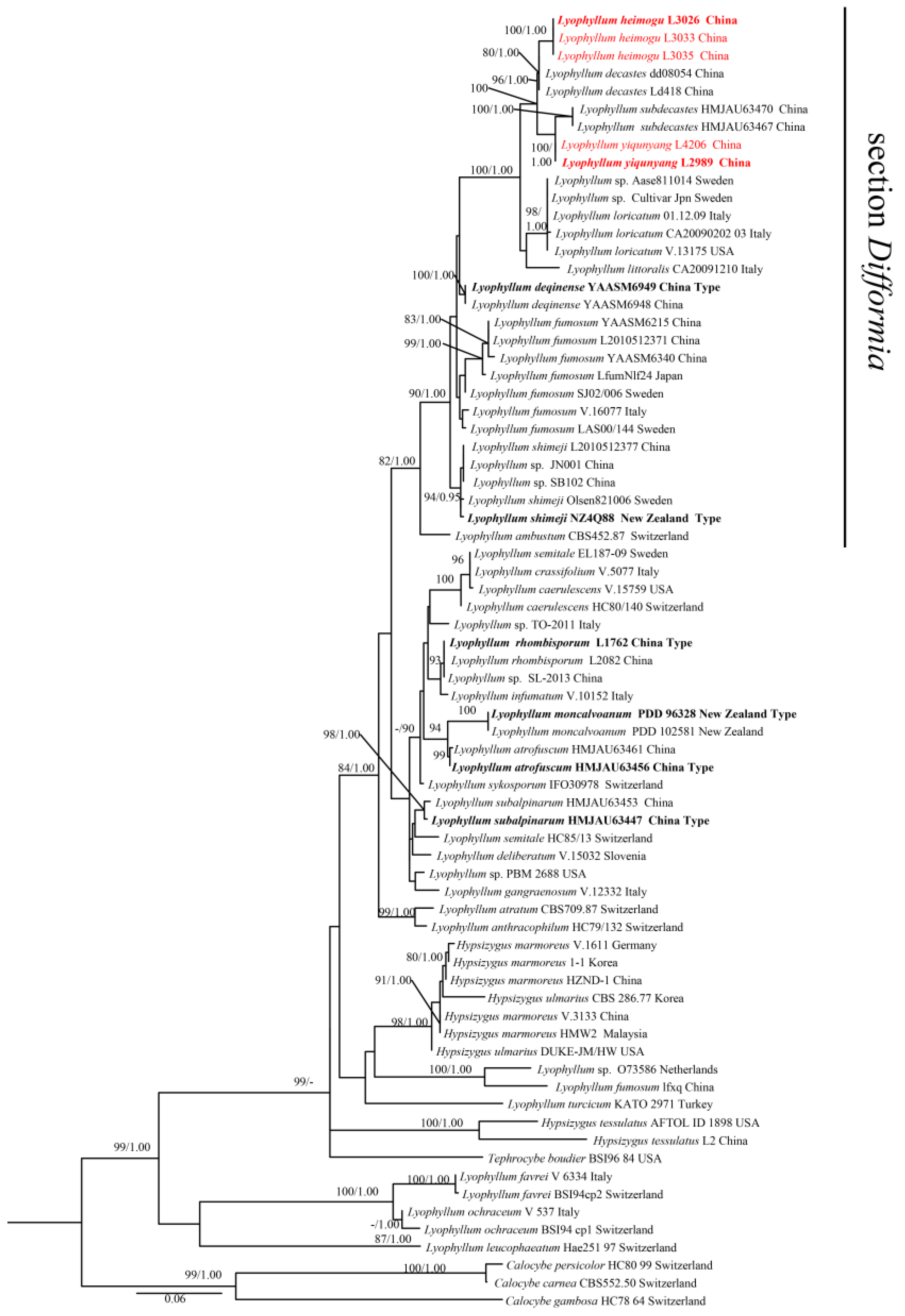
3.1. Phylogenetic Analysis
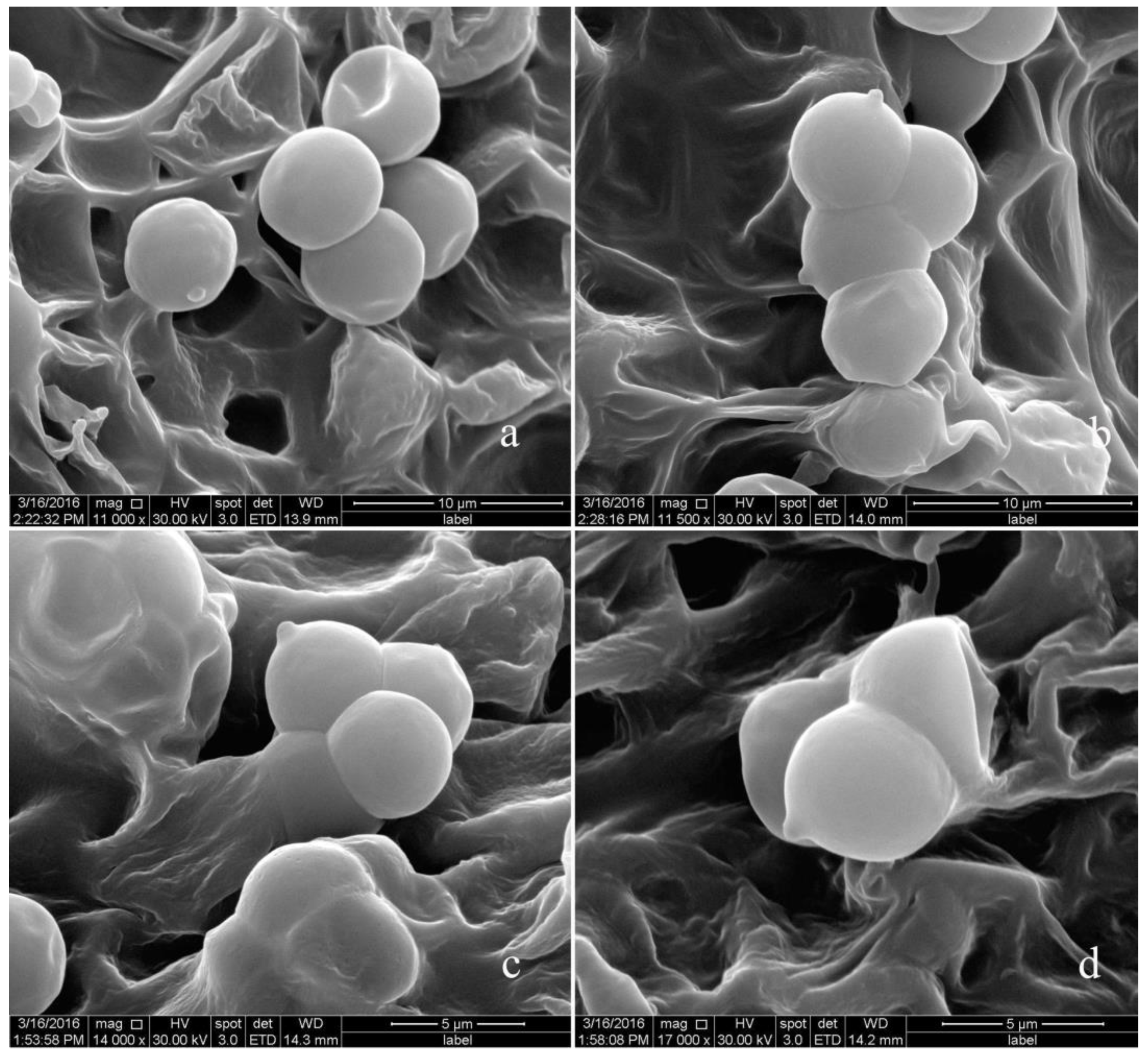
3.2. Taxonomy
4. Discussion
| Taxa | Pileus | Stipe | Spores | Basidia | Reference |
|---|---|---|---|---|---|
| L. decastes | Greyish-brown to yellowish-brown or brown, usually darker when young, 4–12 cm | Whitish-greyish, 5–10 cm | Broadly ellipsoid, 5–7 × 5–7 µm | - | [4,33,34] |
| L. fumosum | Dark to light grey, brown, 2–10 cm | Cream-coloured to brown, 2.5–10 cm | Globose to subglobose, 5.5–7 × 5–7 µm | 40–45 × 8–10 µm | [11] |
| L. heimogu | Dark grey to olive, 3.0–6.5 cm | Yellowish-brown, 2.5–7.0 × 0.5–2.0 cm | Globose to subglobose, 5.30–5.60 × 5.20–5.40 μm | 28.5–32.0 × 7.1–8.3 μm | Present study |
| L. subdecastes | Yellowish-brown, brown to greyish-red | Orange-white, reddish-grey to greyish-red 2.7–6.6 × 0.5–1.5 cm | Globose to subglobose, 3.9–5.0 × 3.7–5.0 μm | 36.7–50.6 × 8.4–10.9 μm | [16] |
| L. shimeiji | Dark grey to grey, brown, 2–8 cm | White, 3–8 cm | Globose to subglobose, 4.0–6.0 × 4.0–6.0 μm | - | [11] |
| L. loricatum | Reddish-brown to chestnut brown 3–12 cm | Cream to pale brownish, grey-brown when old, 3.5–9 × 0.7–1.5 cm | Globose to subglobose, 5.0–6.0 × 4.5–5.3 μm | 28.0–32.0 × 7.0–8.0 µm | [4] |
| L. littoralis | Grey to brownish-grey, 5–15 cm | Grey, 1.5–4 × 0.4–1.5 cm | Globose to subglobose, 4.5–5.5 × 4.5–5.5 μm | - | [36] |
| L. yiqunyang | Olive-grey, 3.0–6.0 cm | White or light grey, 3.0–7.0 × 0.5–1.5 cm | Globose to subglobose, 6.1–6.3 × 6.0–6.2 μm | 31.0–42.0 × 7.8–9.0 μm | Present study |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dähncke, R.M.; Contu, M.; Vizzini, A. Two new species of Lyophyllum s.l. (Basidiomycota, Agaricomycetes) from La Palma (Canary Islands, Spain). Mycotaxon 2011, 115, 65–71. [Google Scholar] [CrossRef]
- Cooper, J.A. New species and combinations of some New Zealand agarics belonging to Clitopilus, Lyophyllum, Gerhardtia, Clitocybe, Hydnangium, Mycena, Rhodocollybia and Gerronema. Mycosphere 2014, 5, 263–288. [Google Scholar] [CrossRef]
- Bon, M. Flore Mycologique d’Europe. Les collybiomarasmïoides et Ressemblants; Doc Mycol Mémoire Hors-Série. n. 5; Association d’Ecologie et de Mycologie: Amiens, France, 1999. [Google Scholar]
- Breitenbach, J. Fungi of Switzerland. Volume 3, Boletes and Agarics 1; Verlag Mycologia: Luzern, Switzerland, 1991. [Google Scholar]
- Clémençon, H.; Smith, A.H. New species of Lyophyllum (Agaricales) from North America and a key to the known staining species. Mycotaxon 1983, 18, 379–437. [Google Scholar]
- Clémençon, H. Schwarzende Lyophyllum-Arten Europas. Z. Mykol. 1986, 52, 61–84. [Google Scholar]
- Hofstetter, V.; Clémençon, H.; Vilgalys, R.; Moncalvo, J.M. Phylogenetic analyses of the Lyophylleae (Agaricales, Basidiomycota) based on nuclear and mitochondrial rDNA sequences. Mycol. Res. 2002, 106, 1043–1059. [Google Scholar] [CrossRef]
- Larsson, E.; Sundberg, H. Lyophyllum shimeji, a species associated with lichen pine forest in northern Fennoscandia. Mycoscience 2011, 52, 289–295. [Google Scholar] [CrossRef]
- Vizzini, A.; Contu, M. Lyophyllum rosae-mariae sp. nov. (Basidiomycota, Agaricomycetes) from La Palma (Canary Islands, Spain). Mycosphere 2010, 1, 83–86. [Google Scholar]
- Yamada, A.; Ogura, T.; Ohmasa, M. Cultivation of mushrooms of edible ectomycorrhizal fungi associated with Pinus densiflora by in vitro mycorrhizal synthesis: I. Primordium and basidiocarp formation in open-pot culture. Mycorrhiza 2001, 11, 59–66. [Google Scholar] [CrossRef]
- Sesli, E.; Vizzini, A.; Contu, M. Lyophyllum turcicum (Agaricomycetes: Lyophyllaceae), a new species from Turkey. Turk. J. Bot. 2015, 39, 512–519. [Google Scholar] [CrossRef]
- He, M.Q.; Zhao, R.L.; Kevin, D.H.; Dominik, B.; Martin, K.; Andrey, Y.; Eric, H.C.; Olivier, R.; Makoto, K.; Santiago, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef]
- Fanglan, D. A Summary of Chinese Fungi; Science Press: Beijing, China, 1979. [Google Scholar]
- Li, X.; Li, Y. Research progress on fungus of the genus Lyophyllum in China. J. Edible Fungi 2009, 16, 75–79. [Google Scholar]
- Wang, X.Q.; Zhou, D.Q.; Zhao, Y.C.; Zhang, X.L.; Li, L.; Li, S.H. Lyophyllum rhombisporum sp. nov. from China. Mycotaxon 2013, 123, 473–477. [Google Scholar] [CrossRef]
- Wei, S.W.; Lu, B.Y.; Wang, Y.; Dou, W.J.; Wang, Q.; Li, Y. Morphology and Phylogeny of Lyophylloid Mushrooms in China with Description of Four New Species. J. Fungi 2023, 9, 77. [Google Scholar] [CrossRef]
- Ma, Y.H.; Liu, P.; Zhao, Z.Y.; Chen, W.M.; Zhao, Y.C. Lyophyllum pallidofumosum sp. nov. (Lyophyllaceae, Agaricales), from southwestern China. Phytotaxa 2022, 576, 173–183. [Google Scholar] [CrossRef]
- Ma, Y.H.; Liu, P.; Zhao, Z.Y.; Chen, W.M.; Zhao, Y.C. Lyophyllum deqinense (Lyophyllaceae, Agaricales), a new species from southwestern China. Phytotaxa 2023, 598, 219–228. [Google Scholar] [CrossRef]
- Visnovsky, S.B.; Cummings, N.; Guerin-Laguette, A.; Wang, Y.; Yamada, A.; Kobayashi, H.; Kawai, M.; Pitman, A.R. Detection of the edible ectomycorrhizal fungus Lyophyllum shimeji colonising seedlings of cultivated conifer species in New Zealand. Mycorrhiza 2014, 24, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics science direct. PCR Protoc. 1990, 18, 315–322. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7, improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T. BioEdit v7. 2007. Available online: http://www.mbio.ncsu.edu/BioEdit/BioEdit.html (accessed on 10 June 2023).
- Stamatakis, A. RAxML version 8, a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Nylander, J.A.A. MrModeltest, v. 2.2.; Program Distributed by the Author; Uppsala University, Department of Systematic Zoology: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.; Teslenko, M. MrBayes Version 3.2 Manual: Tutorials and Model Summaries. 2011. Available online: https://brahms.biology.rochester.edu/software.html (accessed on 10 June 2023).
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Ravikumar, G.; Kalaiselvi, M.; Gomathi, D.; Vidhya, B.; Devaki, K.; Uma, C. Effect of laccase from Hypsizygus ulmarius in decolorization of different dyes. J. Appl. Pharm. Sci. 2013, 3, 150–152. [Google Scholar] [CrossRef]
- Hofstetter, V.; Redhead, S.A.; Kauff, F.; Moncalvo, J.M.; Matheny, P.B.; Vilgalys, R. Taxonomic revision and examination of ecological transitions of the Lyophyllaceae (Basidiomycota, Agaricales) based on a multigene phylogeny. Cryptogam. Mycol. 2014, 35, 399–425. [Google Scholar] [CrossRef]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986. [Google Scholar]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Trudell, S.; Ammirati, J. Mushrooms of the Pacific Northwest. In Timber Press Field Guides; Timber Press: Portland, OR, USA, 2009; pp. 112–113. [Google Scholar]
- Davis, R.; Sommer, R.; John, A. Field Guide to Mushrooms of Western North America; University of California Press: Berkeley, CA, USA, 2012; p. 139. [Google Scholar]
- Miura, T.; Kubo, M.; Itoh, Y.; Iwamoto, N.; Kato, M.; Park, S.R.; Yuuichi, U.; Yukio, K.; Ikukatsu, S.; Suzuki, I. Antidiabetic activity of Lyophyllum decastes in genetically type 2 diabetic mice. Biol. Pharm. Bull. 2002, 2, 1234–1237. [Google Scholar] [CrossRef] [PubMed]
- Ballero, M.; Contu, M. A new species of Calocybe (Agaricales, Lyophylleae) from litoral pine woods of Sardinia (Italy). Mycotaxon 1990, 39, 473–476. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Tang, S.; He, J.; Zhou, D. Two New Edible Lyophyllum Species from Tibetan Areas, China. Diversity 2023, 15, 1027. https://doi.org/10.3390/d15091027
Li S, Tang S, He J, Zhou D. Two New Edible Lyophyllum Species from Tibetan Areas, China. Diversity. 2023; 15(9):1027. https://doi.org/10.3390/d15091027
Chicago/Turabian StyleLi, Shuhong, Songming Tang, Jun He, and Dequn Zhou. 2023. "Two New Edible Lyophyllum Species from Tibetan Areas, China" Diversity 15, no. 9: 1027. https://doi.org/10.3390/d15091027