

Inference of Ploidy Level in 19th-Century Historical Herbarium Specimens Reveals the Identity of Five Acorus Species Described by Schott
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nomenclatural Analysis
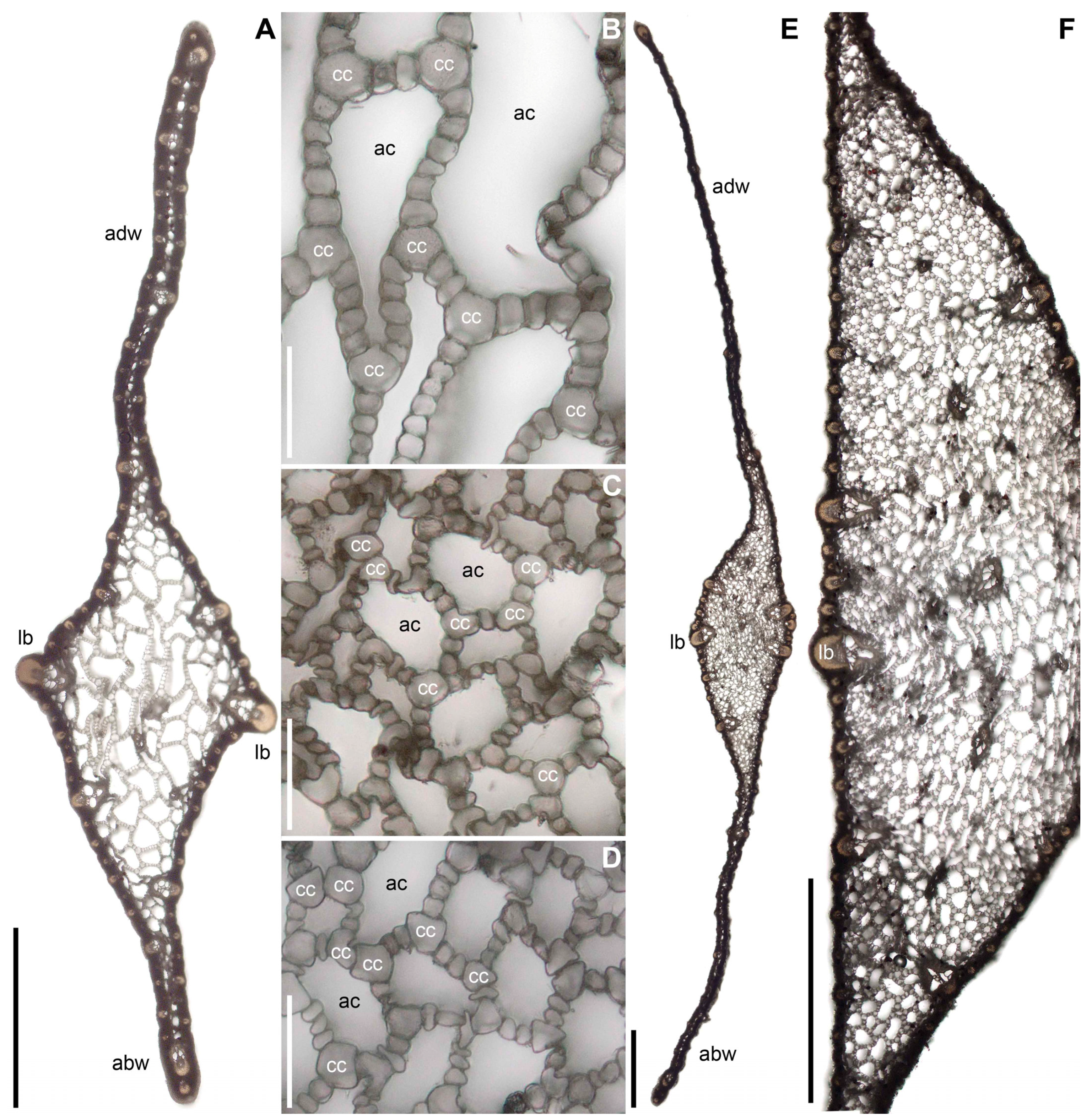
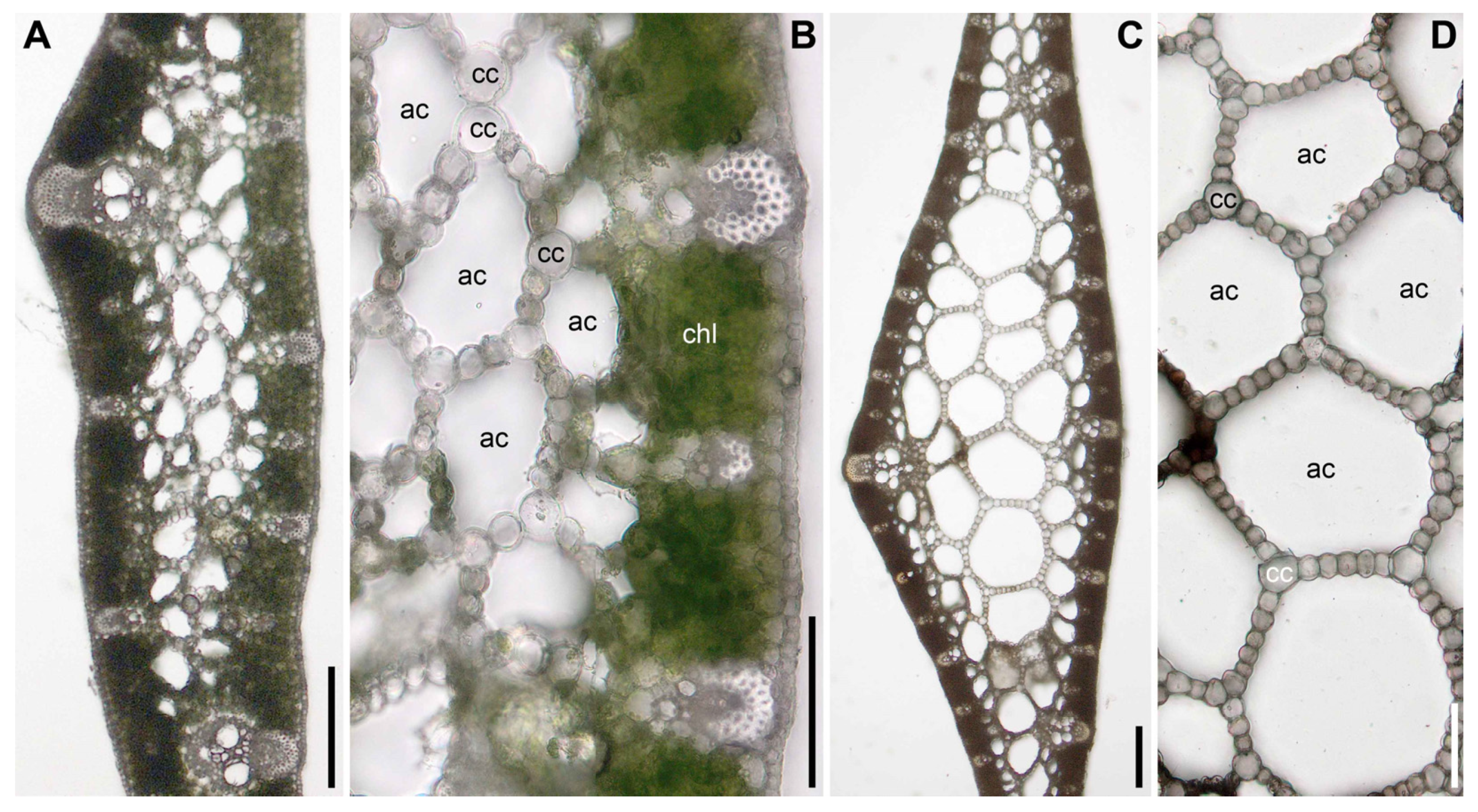
2.2. Ploidy Level
3. Results and Discussion
3.1. Acorus triqueter Is a Diploid from Siberia
3.2. Acorus tatarinowii Is a Tetraploid from N China
3.3. Acorus griffithii Is a Tetraploid from Bhutan
3.4. Acorus commutatus Is Another Name for A. griffithii
3.5. Acorus nilaghirensis Is a Tetraploid from the Peninsular India
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schott, H.W. Aroideen-Skizzen. Oesterr. Bot. Z. 1858, 8, 349–351. [Google Scholar] [CrossRef]
- Schott, H.W. Aroideen-Skizzen. Oesterr. Bot. Z. 1859, 9, 98–101. [Google Scholar] [CrossRef]
- Schott, H.G. Prodromus Systematis Aroidearum; Typis Congregationis Mechitharisticae: Vienna, Austria, 1860. [Google Scholar]
- Schott, H.G.; Araceae, H.W. Pars altera. Nova genera et species, adiectis quibusdam adnotationibus. Ann. Mus. Bot. Lugduno-Batavi 1864, 1, 278–286. [Google Scholar]
- Duvall, M.L.; Learn, G.H., Jr.; Eguiarte, L.E.; Clegg, M.T. Phylogenetic analysis of rbcL sequences identifies Acorus calamus as the primal extant monocotyledon. Proc. Natl. Acad. Sci. USA 1993, 90, 4641–4644. [Google Scholar] [CrossRef] [Green Version]
- Bogner, J.; Mayo, S.J. Acoraceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin, Germany, 1998; Volume 4, pp. 7–11. [Google Scholar]
- The Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Engler, A. Araceae. In Monographiae Phanerogamarum Prodromi Nunc Continuatio, Nunc Revisio; Candolle, A., de Candolle, C., Eds.; G. Masson: Paris, France, 1879; Volume 2. [Google Scholar]
- Engler, A. Araceae-Pothoideae. In Das Pflanzenreich, 21 (IV. 23B); Engler, A., Ed.; Engelmann: Leipzig, Germany, 1905; pp. 1–330. [Google Scholar]
- Röst, L.C.M. Biosystematic investigations with Acorus L. 4. Communication. A synthetic approach to the classification of the genus. Planta Med. 1979, 37, 289–307. [Google Scholar] [CrossRef]
- Li, H.; Zhu, G.; Bogner, J. Acoraceae. In Flora of China; Wu, Z., Raven, P., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MI, USA, 2010; Volume 2, pp. 1–2. [Google Scholar]
- Bogner, J. Acoraceae. In Flora Malesiana, 1st ed.; Noteboom, H.P., Ed.; Nationaal Herbarium Nederland: Leiden, The Netherlands, 2011; Volume 20, pp. 1–13. [Google Scholar]
- Boyce, P. Acoraceae. In Flora of Thailand; Santisuk, T., Larsen, K., Eds.; The Forest Herbarium, Department of National Parks, Wildlife and Plant Conservation: Bangkok, Thailand, 2012; Volume 11, pp. 322–325. [Google Scholar]
- Murata, J. Acoraceae. In Flora of Japan; Iwatsuki, K., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 2016; Volume IVb, p. 5. [Google Scholar]
- Linnaeus, C. Species Plantarum; L. Salvius: Holmia, Sweden, 1753. [Google Scholar]
- Aiton, W. Hortus Kewensis, or a Catalogue of the Plants Cultivated in the Royal Botanic Garden at Kew; George Nicol: London, UK, 1789; Volume 1. [Google Scholar]
- Löve, Á.; Löve, D. Drug content and polyploidy in Acorus. Proc. Genet. Soc. Can. 1957, 2, 14–17. [Google Scholar]
- Packer, J.G.; Ringius, G.S. The distribution and status of Acorus (Araceae) in Canada. Can. J. Bot. 1984, 62, 2248–2252. [Google Scholar] [CrossRef]
- Löve, Á.; Löve, D.; Löve, Á. Chromosome Number Reports LXXXIX. Taxon 1985, 34, 729–730. [Google Scholar] [CrossRef]
- Petersen, G. Cytology and systematics of Araceae. Nord. J. Bot. 1989, 9, 119–166. [Google Scholar] [CrossRef]
- Thompson, S.A. Systematics and Biology of the Araceae and Acoraceae of Temperate North America. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 1995. [Google Scholar]
- Thompson, S.A. Acoraceae. In Flora of North America North of Mexico; Magnoliophyta: Alismatidae, Arecidae, Commelinidae (In Part), and Zingiberidae. Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2000; Volume 22, pp. 124–127. [Google Scholar]
- Larsen, K. Cytology of vascular plants III. A study of Thai aroids. Dansk Bot. Ark. 1969, 27, 39–59. [Google Scholar]
- Li, H. Acorus. In Flora Reipublicae Popularis Sinicae; Wu, C., Li, H., Eds.; Science Press: Beijing, China, 1979; Volume 13, pp. 4–9. (In Chinese) [Google Scholar]
- Wei, F.N.; Li, Y.K. A new spice, Acorus macrospadiceus from south China. Guihaia 1985, 5, 179–182. (In Chinese) [Google Scholar]
- Zhu, Z. Some new taxa of Acorus (Araceae) from Sichuan. Acta Bot. Bor.-Occ. Sinica 1985, 5, 118–121. (In Chinese) [Google Scholar]
- Ho, P.H. An Illustrated Flora of Vietnam; Nha Xuat Ban Tre: Ho Chi Minh, Vietnam, 2000; Volume 3, pp. 1–1020. (In Vietnamese) [Google Scholar]
- Zhang, W. Pharmacognostic Study of Acorus—From a Phylogenetic Perspective. Ph.D. Thesis, Hong Kong Baptist University, Hong Kong, China, 2005. [Google Scholar]
- Shu, H.; Zhang, S.; Lei, Q.; Zhou, J.; Ji, Y.; Luo, B.; Hong, L.; Li, F.; Liu, B.; Long, C. Ethnobotany of Acorus in China. Acta Soc. Bot. Polon. 2018, 87, 3585. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Shu, H.; Zhang, S.; Luo, B.; Gu, R.; Zhang, R.; Ji, Y.; Li, F.; Long, C. From folk taxonomy to species confirmation of Acorus (Acoraceae): Evidences based on phylogenetic and metabolomic analyses. Front. Plant Sci. 2020, 11, 965. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Remizowa, M.V.; Nuraliev, M.S.; Averyanov, L.V.; Sennikov, A.N. The first genome from the basal monocot family has been misnamed: Taxonomic identity of Acorus tatarinowii (Acoraceae), a source of numerous chemical compounds of pharmaceutical importance. Diversity 2023, 15, 176. [Google Scholar] [CrossRef]
- Wulff, H.D. Zur Zytologie, geographischen Verbreitung und Morphologie des Kalmus. Arch. Pharm. 1954, 287, 529–541. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2023. Available online: http://www.plantsoftheworldonline.org/ (accessed on 29 March 2023).
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code). In Proceedings of the Adopted by the Nineteenth International Botanical Congress, Shenzhen, China, 23–29 July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Wulff, H.D. Über die Ursache der Sterilität des Kalmus (Acorus calamus L). Planta 1940, 31, 478–491. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Skaptsov, M.V.; Vislobokov, N.A.; Smirnov, S.V.; Shmakov, A.I.; Remizowa, M.V. Morphological characterization of diploid and triploid Acorus calamus (Acoraceae) from southern Western Siberia, parthenocarpy in sterile plants and occurrence of aneuploidy. Bot. J. Linn. Soc. 2021, 195, 189–215. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Remizowa, M.V.; Skaptsov, M.V.; Yadav, S.R.; Sennikov, A.N. Back to Linnaeus: Proper botanical naming of the tetraploid Indian Acorus (Acoraceae), an important medicinal plant. Diversity. submitted.
- Shi, T.; Huneau, C.; Zhang, Y.; Li, Y.; Chen, J.; Salse, J.; Wang, Q. The slow-evolving Acorus tatarinowii genome sheds light on ancestral monocot evolution. Nat. Plants 2022, 8, 764–777. [Google Scholar] [CrossRef]
- Wulff, H.D. Der Ölgehalt verschiedenchromosomiger Rassen vom Kalmus (Acorus Calamus L.). Zeitschr. Naturforsch. A 1946, 1, 600–603. [Google Scholar] [CrossRef]
- Wulff, H.D. Ölgehalt und Chromosomenzahl des nordamerikanischen Kalmus (Acorus calamus L.). Arch. Pharm. 1950, 283, 155–161. [Google Scholar] [CrossRef]
- Röst, L.C.M.; Bos, R. Biosystematic investigations with Acorus L. 3. Communication. Constituents of essential oils. Planta Med. 1979, 36, 350–361. [Google Scholar] [CrossRef]
- Mittal, N.; Varshney, V.K.; Song, B.H.; Ginwal, H.S. High levels of diversity in the phytochemistry, ploidy and genetics of the medicinal plant Acorus calamus L. Med. Aromat. Plants 2015, S1, 002. [Google Scholar] [CrossRef]
- Ahlawat, A.; Katoch, M.; Rama, G.; Ahuja, A. Genetic diversity in Acorus calamus L. as revealed by RAPD markers and its relationship with β-asarone content and ploidy level. Sci. Hortic. 2010, 124, 294–297. [Google Scholar] [CrossRef]
- Röst, L.C.M. Biosystematic investigations with Acorus L. (Araceae). 1. Communication. Cytotaxonomy. Proc. K Ned. Akad. Wet. 1978, 81, 428–441. [Google Scholar]
- Turczaninow, N. Flora Baicalensi-Dahurica seu Descriptio plantarum in regionibus cis- et transbaicalensibus atque in Dahuria sponte nascentium. Bull. Soc. Imp. Nat. Moscou 1854, 27, 53–130. [Google Scholar]
- Borodin, I. Collectors and Collections in the Flora of Siberia; Imperial Academy of Sciences: Saint-Petersburg, Russia, 1908. (In Russian) [Google Scholar]
- Mücke, M. Über den Bau und die Entwicklung der Früchte und über die Herkunft von Acorus calamus L. Bot. Zeit. 1908, 66, 1–23. [Google Scholar]
- Vinogradova, Y.K. Variability of Acorus calamus L. biological characteristics in natural and invasive populations. Byull. Glavn. Bot. Sada 2004, 187, 23–31. (In Russian) [Google Scholar]
- Chepinoga, V.V.; Gnutikov, A.A.; Enushchenko, I.V.; Chepinoga, A.V. Acorus calamus. In:Marhold, K., Ed. IAPT/IOPB Chromosome Data 6. Taxon 2008, 57, 1267, E1. [Google Scholar] [CrossRef]
- Krivenko, D.A.; Kotseruba, V.V.; Kazanovsky, S.G.; Verkhozina, A.V.; Chernova, O.D. Acorus calamus. In: Marhold, K., Ed IAPT/IOPB chromosome data 16. Taxon 2013, 62, 1357, E4. [Google Scholar]
- Probatova, N.S.; Gnutikov, A.A.; Rudyka, E.G.; Chepinoga, V.V. Chromosome numbers of some plant species from Baikal Siberia. Bot. Zhurn. 2008, 93, 162–181. [Google Scholar]
- Chepinoga, V.V. Chromosome Numbers of Plant Species from Baikal Siberia; Nauka: Novosibirsk, Russia, 2014. [Google Scholar]
- de Loureiro, J. Flora Cochinchinensis: Sistens Plantas in Regno Cochinchina Nascentes. Quibus Accedunt Aliæ Observatæ in Sinensi Imperio, Africa Orientali, Indiæque Locis Variis. Omnes Dispositæ Secundum Systema Sexuale Linnæanum; Ulyssipone: Lisboa, Portugal, 1790. [Google Scholar]
- Hooker, J.D. Works of the late William Griffith, Esq. F. L. S. & C. Posthumous papers, bequeathed to the Hon. East India Company, and printed by the order of the Government of Bengal. Lond. J. Bot. 1848, 7, 446–449. [Google Scholar]
- Hooker, J.D. Catalogue of the Plants Distributed at the Royal Gardens, Kew, under the Sanction of the Secretary of State for India, from the Herbaria of Griffith, Falconer, and Helfer; J.E. Taylor: London, UK, 1865. [Google Scholar]
- Lang, W.H. William Griffith, 1810–1845. In Makers of British Botany: A Collection of Biographies by Living Botanists; Oliver, F.W., Ed.; Cambridge University Press: Cambridge, UK, 1913; pp. 178–191. [Google Scholar]
- Huxley, L. (Ed.) Life and Letters of Sir Joseph Dalton Hooker; J. Murray: London, UK, 1918; Volume 1. [Google Scholar]
- Griffith, W. Itinerary Notes of Plants Collected in the Khasyah and Bootan Mountains, 1837–1838, in Affghanisthan and Neighbouring Countries, 1839 to 1841; J.F. Bellamy: Calcutta, UK, 1848. [Google Scholar]
- Griffith, W. Journals of Travels in Assam, Burma, Bootan, Affghanistan and the Neighbouring Countries; Bishop’s College Press: Calcutta, UK, 1847. [Google Scholar]
- Hooker, J.D.; Thomson, T. Introductory Essay to the Flora Indica; W. Pamplin: London, UK, 1855. [Google Scholar]
- Anonymous. Intended distribution of the Herbaria of the East India Company. Bot. Gaz. 1849, 1, 112. [Google Scholar]
- Noltie, H.J. Flora of Bhutan Including a Record of Plants from Sikkim and Darjeeling; Royal Botanic Garden Edinburgh: Edinburgh, UK, 1994; Volume 3. [Google Scholar]
- Ogra, R.K.; Mohanpuria, P.; Sharma, U.K.; Sharma, M.; Sinha, A.K.; Ahuja, P.S. Indian calamus (Acorus calamus L.): Not a tetraploid. Curr. Sci. 2009, 97, 1644–1647. [Google Scholar]
- Griffith, W. Icones Plantarum Asiaticarum; C.A. Serrao: Calcutta, UK, 1851; Volume 3. [Google Scholar]
- Griffith, W. Notulae ad Plantas Asiaticas; C.A. Serrao: Calcutta, UK, 1851; Volume 3. [Google Scholar]
- Bliss, E.M. (Ed.) The Encyclopaedia of Missions; Funk & Wagnalls: New York, NY, USA, 1891; Volume 1. [Google Scholar]
- Stewart, R.R. Missionaries and clergymen as botanists in India and Pakistan. Taxon 1982, 31, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Fürnrohr, A.E. Getrocknete Pflanzensammlungen: Plantarum Indiae orientalis Sect. V. Flora 1854, 37, 187–189. [Google Scholar]
- Hohenacker, R.F. Verkäufiche Pflanzensammlungen. Oesterr. Bot. Z. 1859, 9, 1–4. [Google Scholar]
- Wörz, A. Der Esslinger Botanische Reiseverein 1825–1845: Eine Aktiengesellschaft zur Durchführung Naturkundlicher Sammelreisen; Logos Verlag: Berlin, Germany, 2016. [Google Scholar]
- Hohenacker, R.F. Anzeige: Sammlungen ostindischer Pflanzen aus Canara und den Nilgherries. Flora 1851, 34, 718–720. [Google Scholar]
- Hohenacker, R.F. (Ed.) Plantarum Indiae orientalis Sect. V. Cura Rev. Fr. Metz, Missionarii Evangelicae Societatis Basileensis, Pro Maximu Parte in Montibus Nilagiri Collectas, a DD. Bentham, Fenzl, Hochstetter, Meisner, Miquel, de Schlechtendal, C.H. Schultz Bip., Steudel aliisque Examinatas Edidit R.F. Hohenacker; Esslinger Botanische Reiseverein: Esslingen, Kingdom of Württemberg, 1853. [Google Scholar]
- British Museum. The History of the Collections Contained in the Natural History Departments of the British Museum; Trustees of the British Museum: London, UK, 1904; Volume 1. [Google Scholar]
- Lipsky, V.I. Herbarium. In Historical Account of the Imperial Botanical Garden in Saint-Petersburg for 1873–1898; Fischer von Waldheim, A.A., Ed.; Imperial Botanical Garden: Saint-Petersburg, Russia, 1899; pp. 36–160. (In Russian) [Google Scholar]
- Hohenacker, R.F. Anzeigen: Getrocknete Pflanzen aus Klein-Asien, Sibirien und Ost-Indien. Flora 1847, 30, 622. [Google Scholar]
- Riedl, H. Heinrich Wilhelm Schott (1794–1865). Taxon 1965, 14, 209–213. [Google Scholar] [CrossRef]
- Wang, H.; Li, W.L.; Gu, Z.J.; Chen, Y.Y. Cytological study on Acorus L. in southwestern China, with some cytogeographical notes on A. calamus. Acta Bot. Sinica 2001, 43, 354–358. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen ID | Schott’s Species Name | Nomencla- Tural Status | Number of Staining/Nonstaining Pollen Grains Counted (% of Fertile) | Fruits with Seeds | Number of air Canals Per 0.62 mm2 of the Cross Section of Leaf Blade (Mean of Two Fields of View) | Mean Cell Number in 20 Septa in Cross Section (Corner Cells not Counted) | Inferred Ploidy Level |
|---|---|---|---|---|---|---|---|
| LE 01082852 | A. triqueter | lectotype | n/a | present | 9.75 | 5.4 | diploid |
| LE 01082856 | A. triqueter | isolectotype | n/a | present | 22.5 | 4.5 | diploid |
| LE 01182327 | A. tatarinowii | isotype | 101/3 (97%) | absent | 107.25 | 1.6 | tetraploid |
| LE 01182325 | A. tatarinowii | isotype | 112/1 (99%) | absent | 134.5 | 1.9 | tetraploid |
| LE 01082877 | A. griffithii | isotype | 50/1 (98%) | absent | n/a | 1.2 | tetraploid |
| LE 01082855 | A. nilaghirensis | isolectotype | 14/1 (93%) | absent | n/a | 1.2 | tetraploid |
| Herbarium Collection (See Labels in Figure 4) | c1 | c2 | c3 |
|---|---|---|---|
| Griffith’s collection number | 519 | 1254 | 1254 |
| E.I.C. distribution number | 2626 | no distribution | no distribution |
| Kew distribution number | 5941 | 5941 | 5941 |
| herbarium specimens | K 000883788, K 000883697 (pro parte), BM 000958483, LE 01082877, P 02137244 | K 002467072 (pro parte), P 02137232 (pro parte) | K 000883697 (pro parte), K 002467072 (pro parte), P 02137232 (pro parte) |
| collection date | 1–4 February 1838 | 15–21 September 1837 | 12 November 1837 |
| present-day country | Bhutan | Bangladesh | India |
| locality as printed on distribution labels | East Himalaya | East Bengal | East Bengal |
| locality as written on distribution labels | Bhotan | Khasia | Khasia |
| locality as in Griffith’s travel diaries | Benka | Sylhet | Nurtung |
| locality in present-day terminology | Tashigang District: Tashigang Town | Sylhet Division: from Habiganj Town to Surma River west of Chhatak Town | Meghalaya State: West Jaintia Hills District, Nartiang Town |
| estimated coordinates | 27.339° N, 91.547° E | between 24.4° N, 91.4° E and 25° N, 91.62° E | 25.566° N, 92.21° E |
| estimated elevation | 700 m | 50 m | 1200 m |
| phenological phase | male stage of anthesis | postanthetic | postanthetic (one inflorescence abortive?) |
| characteristic features | plants less than 40 cm tall, leaves less than 1 cm wide, young leaves present, spadix < 5 cm long | plants c. 50 cm tall, leaves less than 1 cm wide, distal parts of leaves longitudinally split during herborization, young leaves absent, spadix < 5 cm long | plants >70 cm tall, leaves more than 1 cm wide, foliage appears to be senescent, spadix > 5 cm long |
| inferred ploidy level | tetraploid | pending destructive sampling | pending destructive sampling |
| nomenclatural status | type collection of Acorus griffithii | no type status | no type status |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokoloff, D.D.; Remizowa, M.V.; Severova, E.E.; Sennikov, A.N. Inference of Ploidy Level in 19th-Century Historical Herbarium Specimens Reveals the Identity of Five Acorus Species Described by Schott. Diversity 2023, 15, 766. https://doi.org/10.3390/d15060766
Sokoloff DD, Remizowa MV, Severova EE, Sennikov AN. Inference of Ploidy Level in 19th-Century Historical Herbarium Specimens Reveals the Identity of Five Acorus Species Described by Schott. Diversity. 2023; 15(6):766. https://doi.org/10.3390/d15060766
Chicago/Turabian StyleSokoloff, Dmitry D., Margarita V. Remizowa, Elena E. Severova, and Alexander N. Sennikov. 2023. "Inference of Ploidy Level in 19th-Century Historical Herbarium Specimens Reveals the Identity of Five Acorus Species Described by Schott" Diversity 15, no. 6: 766. https://doi.org/10.3390/d15060766