
Stratigraphic and Paleoecological Significance of the Early/Middle Pleistocene Vertebrate Fauna of the Süttő 21 Site
 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
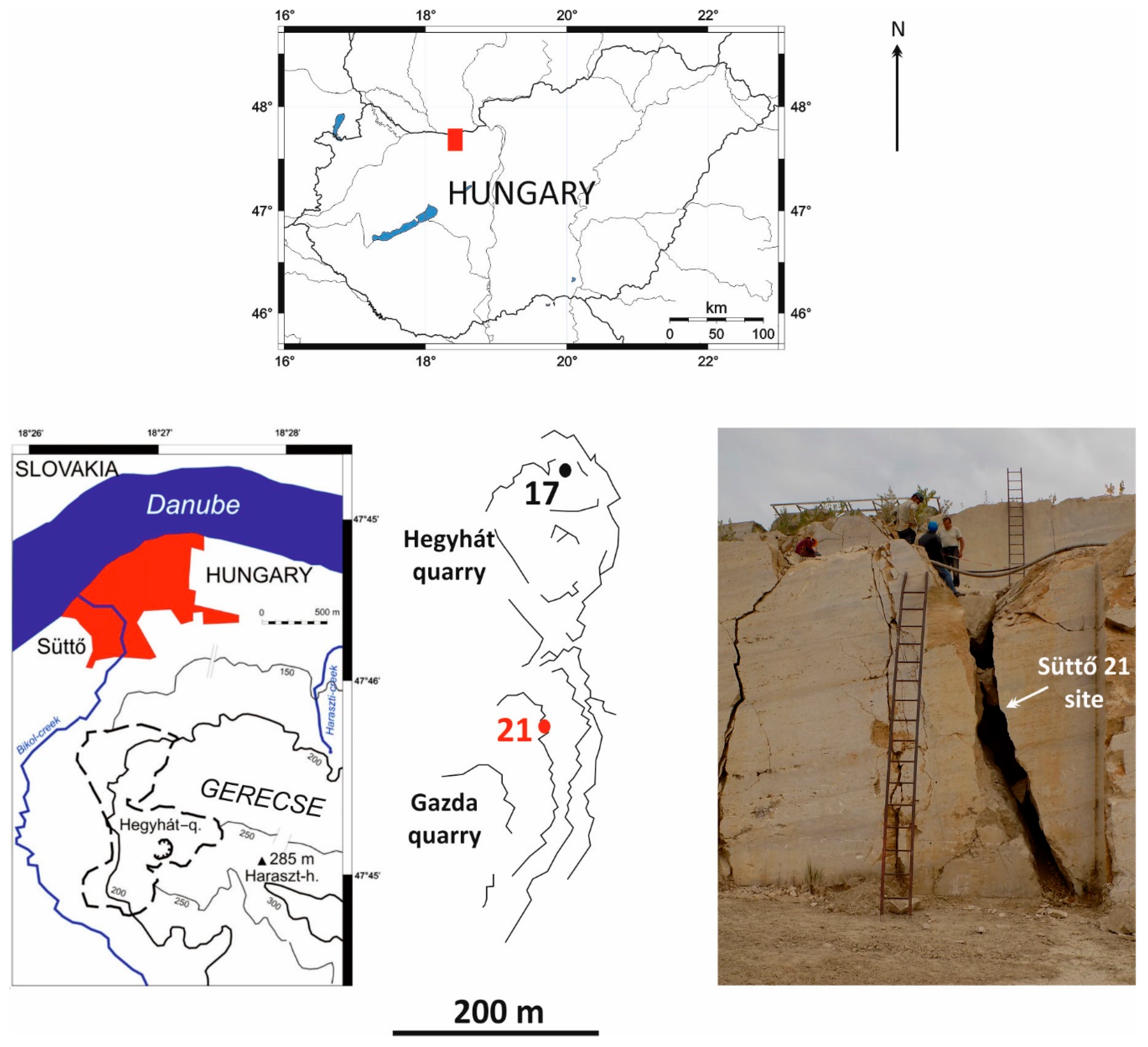
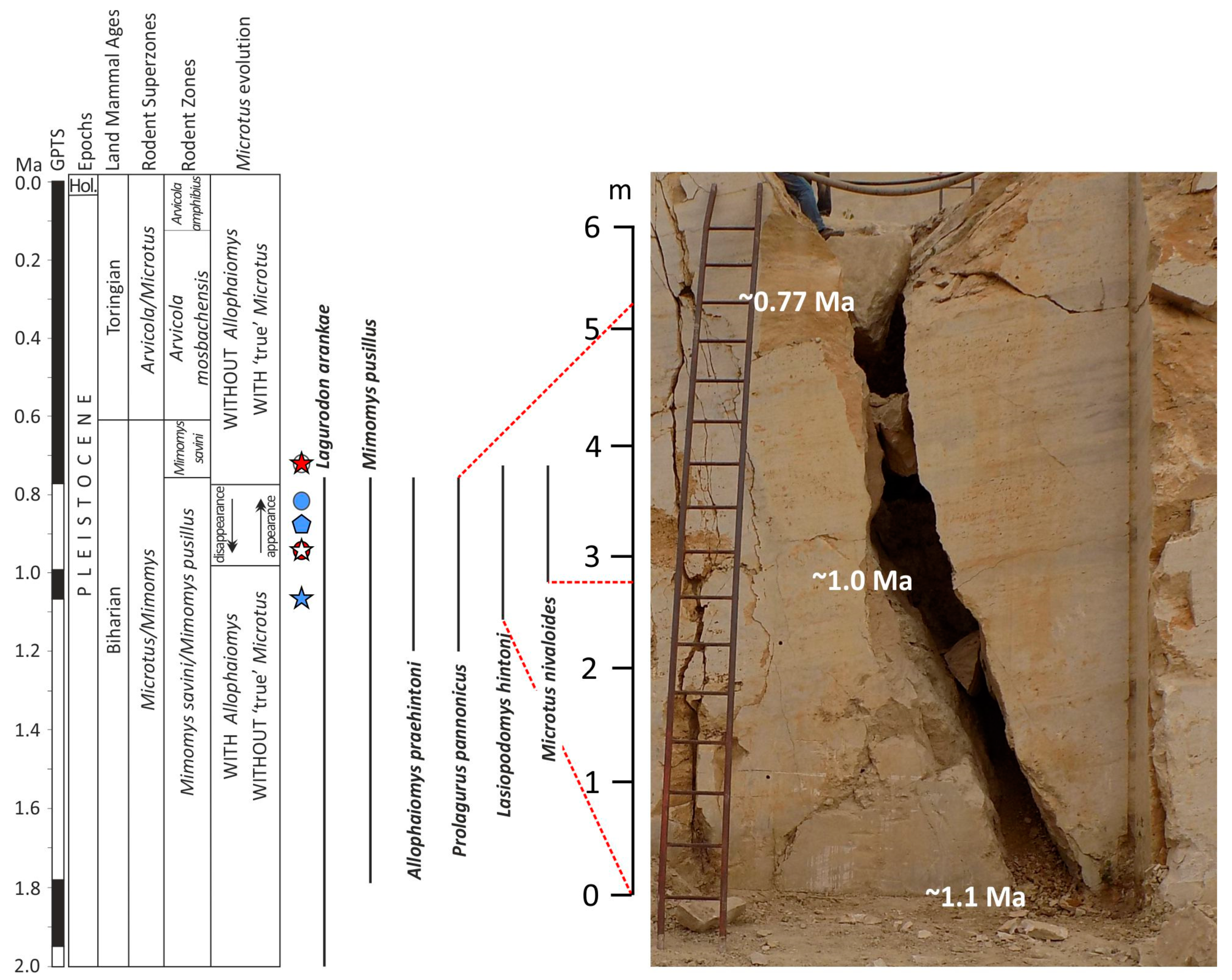
2.1. Süttő 21 Site
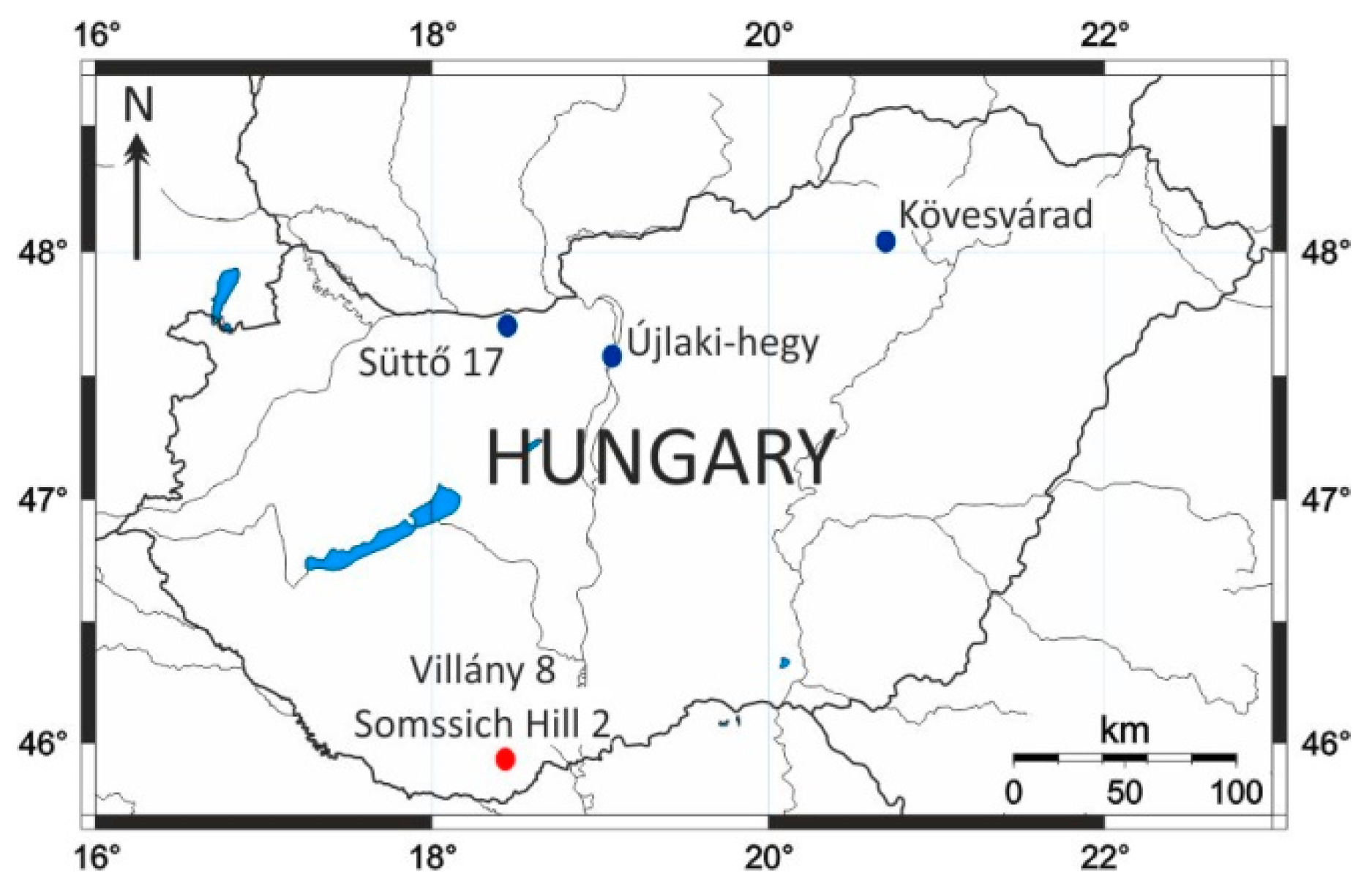
2.2. Sites of a Similar Age Included in the Study
2.2.1. Villány Hills (Southern Hungary)
2.2.2. Northern Hungary
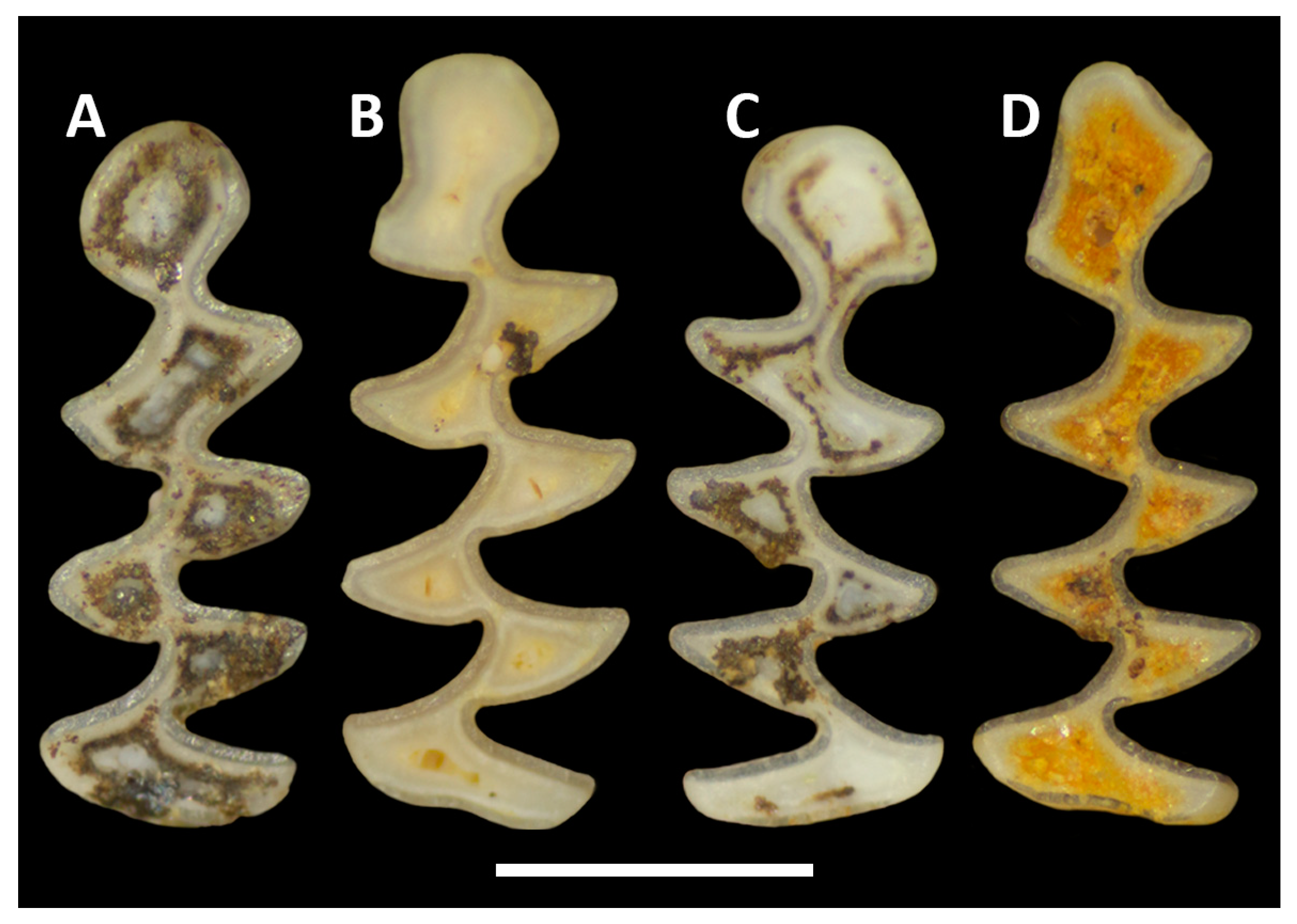
3. Biostratigraphic Results
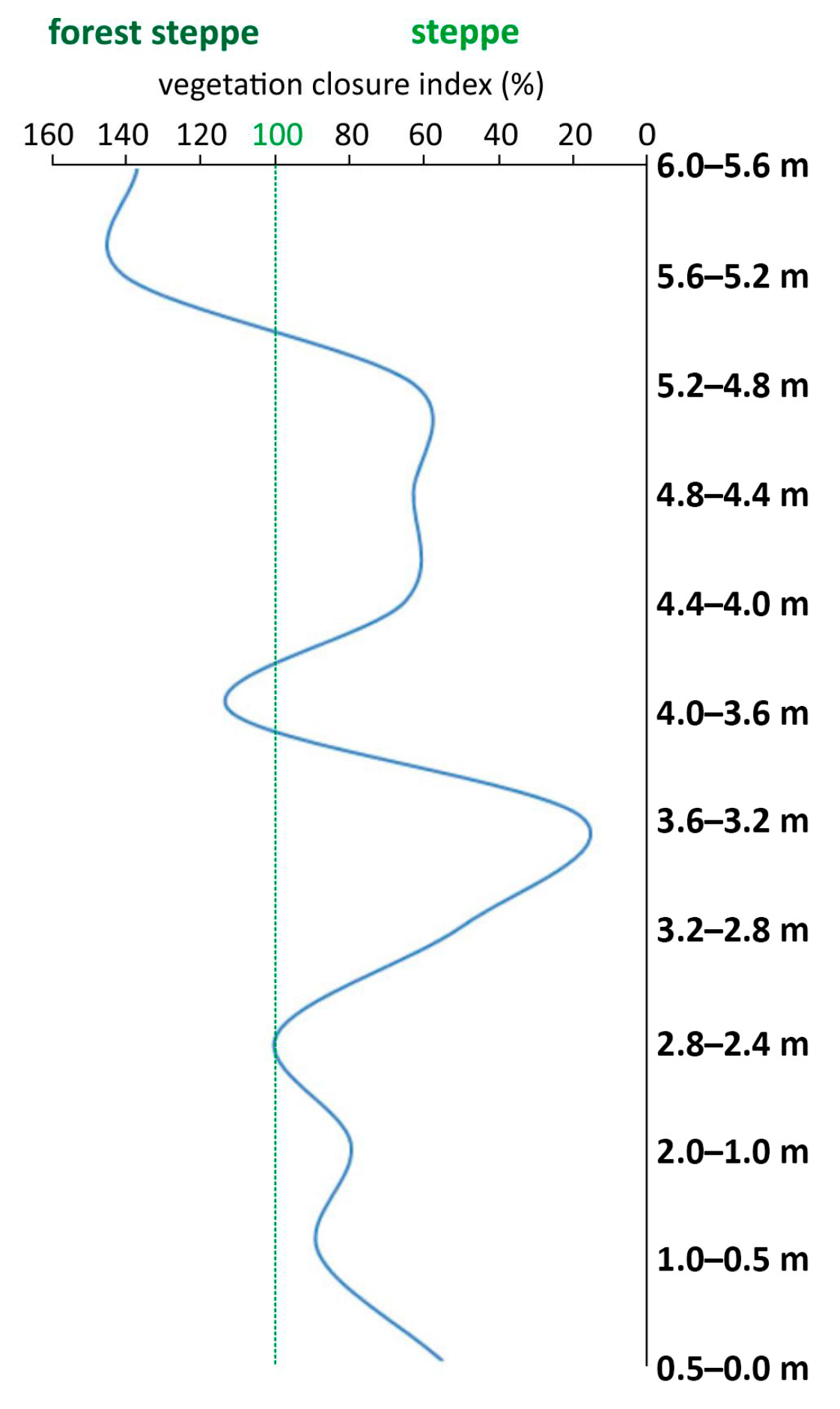
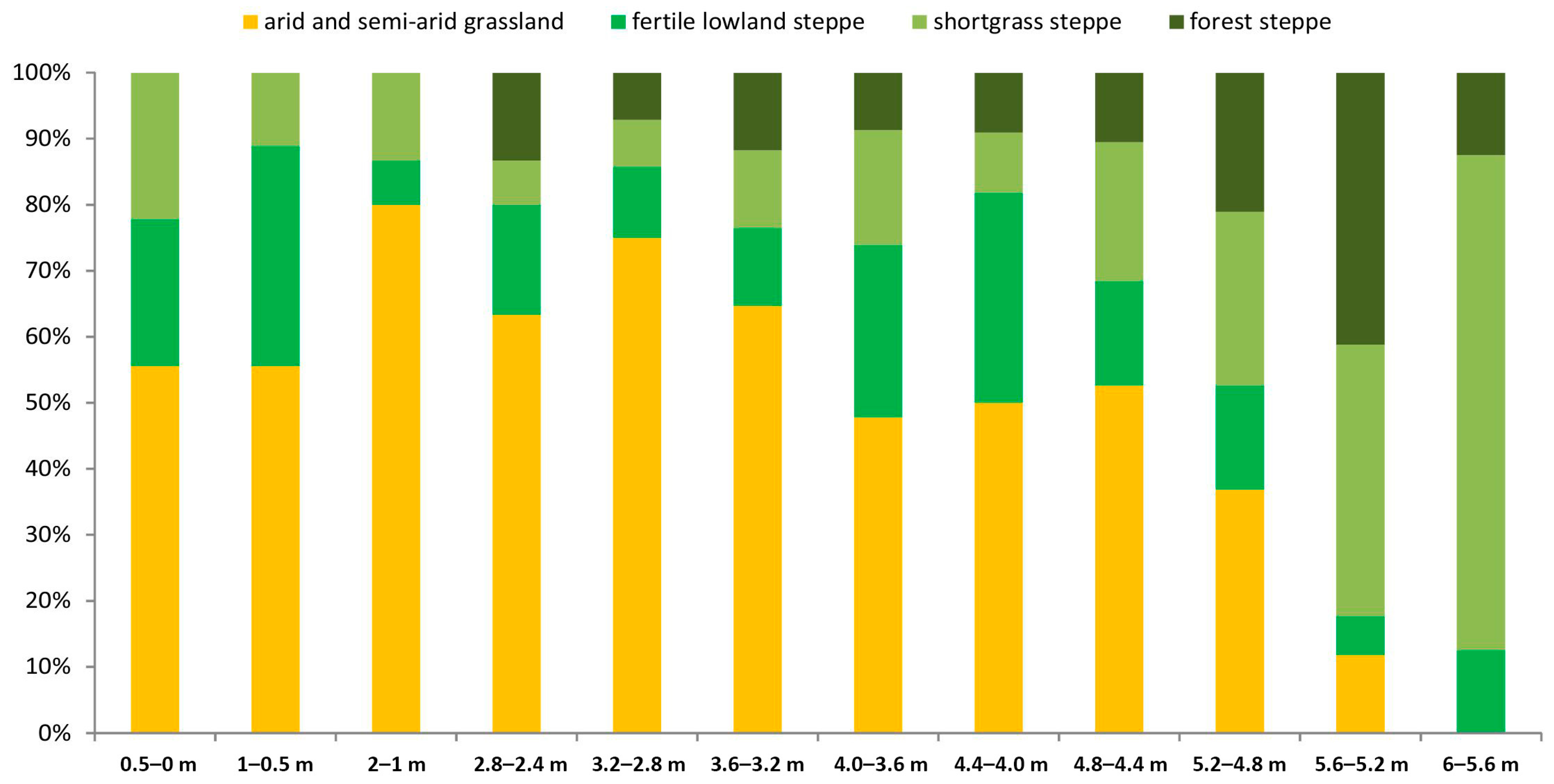
4. Paleoecological Evaluation
5. Comparison of the Süttő 21 Site with Sites of a Similar Age in Hungary
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kretzoi, M. A Negyedkor Taglalása Gerinces Faunák Alapján [An Analysis of the Quarter Age Based on Vertebrate Faunas]; A MTA Műszaki Tudományok Osztályának Alföldi Kongresszusa: Budapest, Hungary, 1953; pp. 89–99. [Google Scholar]
- Jánossy, D. Pleistocene Vertebrate Faunas of Hungary; Akadémiai Kiadó: Budapest, Hungary; Elsevier Sciences Publishers: Amsterdam, The Netherlands; Oxford, UK; New York, NY, USA; Tokyo, Japan, 1986; pp. 1–208. [Google Scholar]
- Pazonyi, P.; Kordos, L.; Magyari, E.; Marinova, E.; Fűköh, L.; Venczel, M. Pleistocene Vertebrate Faunas of the Süttő Travertine Complex. Quat. Int. 2014, 319, 50–63. [Google Scholar] [CrossRef] [Green Version]
- Ruszkiczay-Rüdiger, Z.; Csillag, G.; Fodor, L.; Braucher, R.; Novothny, Á.; Thamó-Bozsó, E.; Virág, A.; Pazonyi, P.; Timár, G.; ASTER Team. Integration of new and revised chronological data to constrain the terrace evolution of the Danube River (Gerecse Hills, Pannonian Basin). Quat. Geochronol. 2018, 48, 148–170. [Google Scholar] [CrossRef]
- Jánossy, D. Arvicolids from the Lower Pleistocene sites at Beremend 15 and Somssich-hegy 2, Hungary. In International Symposium Evolution, Phylogeny and Biostratigraphy of Arvicolids (Rodentia, Mammalia); Fejfar., O., Heinrich, W.-D., Eds.; Geological Survey of Czech Republic: Prague, Czech Republic, 1990; pp. 223–230. [Google Scholar]
- Pazonyi, P.; Virág, A.; Gere, K.; Botfalvai, G.; Sebe, K.; Szentesi, Z.; Mészáros, L.; Botka, D.; Gasparik, M.; Korecz, L. Sedimentological, Taphonomical and Palaeoecological Aspects of the Late Early Pleistocene Vertebrate Fauna from the Somssich Hill 2 Site (South Hungary). Comptes Rendus Palevol 2018, 17, 296–309. [Google Scholar] [CrossRef]
- Kordos, L. Revised Biostratigraphy of the Early Man Site at Vértesszőlős, Hungary. Cour. Forsch. 1994, 171, 225–236. [Google Scholar]
- Maul, L.C.; Parfitt, S.A. Micromammals from the 1995 Mammoth Excavation at West Runton, Norfolk, UK: Morphometric data, biostratigraphy and taxonomic reappraisal. Quat. Int. 2010, 228, 91–115. [Google Scholar] [CrossRef] [Green Version]
- Pazonyi, P.; Virág, A.; Podani, J.; Pálfy, J. Microtus (Microtus) nivaloides from the Somssich Hill 2 site (southern Hungary): An Early Pleistocene forerunner of modern ’true’ Microtus voles revealed by morphometric analyses. Quat. Int. 2018, 481, 61–74. [Google Scholar] [CrossRef]
- Kretzoi, M.; Pécsi, M. Pliocene and Quaternary chronostratigraphy and continental surface development of the Pannonian Basin. In Quaternary Studies in Hungary; Pécsi, M., Ed.; Geographical Research Institute Hungarian Academy of Sciences: Budapest, Hungary, 1982; pp. 11–42. [Google Scholar]
- Maul, L.C.; Markova, A.K. Similarity and Regional Differences in Quaternary Arvicolid Evolution in Central and Eastern Europe. Quat. Int. 2007, 160, 81–99. [Google Scholar] [CrossRef]
- Jánossy, D.; Topál, G. Allophaiomys-Fauna vom Újlaki-hegy (Budapest). Ann. Musei Hist.-Nat. Hung. 1990, 81, 17–26. [Google Scholar]
- Jánossy, D. Die altpleistozäne Wirbeltierfauna von Kövesvárad bei Répáshuta (Bükk-Gebirge). Ann. Musei Hist.-Nat. Hung. 1963, 55, 109–141. [Google Scholar]
- Markova, A.K. European Small Mammal Faunas during the End of the Early Pleistocene—The Beginning of the Middle Pleistocene. In Proceedings of the INQUA-SEQS 2014: The Quaternary of the Urals: Global Trends and Pan-European Quaternary Records, Ekaterinburg, Russia, 10–16 September 2014. [Google Scholar]
- Maul, L. Die Kleinsäugerreste (Insectivole, Lagomorpha, Rodentia) aus dem Unterpleistozän von Untermassfeld. In Das Pleistozän von Untermassfeld bei Meiningen (Thüringen), Monographien RGZM; Kahlke, R.-D., Ed.; Rudolf Habelt: Bonn, Germany, 2001; Volume 3, pp. 783–887. [Google Scholar]
- Maul, L.C. New Results on Micromammals from the Early Pleistocene Site of Untermassfeld. In The Pleistocene of Untermassfeld near Meiningen (Thüringen, Germany) Part 5; Kahlke, R.-D., Ed.; Verlag des Römisch-Germanischen Zentralmuseums: Bad Langensalza, Germany, 2022; pp. 1567–1626. [Google Scholar]
- Abramson, N.I.; Bodrov, S.Y.; Bondareva, O.V.; Genelt-Yanovskiy, E.A.; Petrova, T.V. A mitochondrial genome phylogeny of voles and lemmings (Rodentia: Arvicolinae): Evolutionary and taxonomic implications. PLoS ONE 2021, 16, e0248198. [Google Scholar] [CrossRef]
- Nadachowski, A. Comments on variation, evolution and phylogeny of Chionomys (Arvicolidae). In International Symposium Evolution, Phylogeny and Biostratigraphy of Arvicolids (Rodentia, Mammalia); Fejfar, O., Heinrich, W.-D., Eds.; Geological Survey: Prague, Czech Republic, 1990; pp. 353–368. [Google Scholar]
- Hír, J. A Comparative study on the dental morphology of the Early Pleistocene Cricetus praeglacialis SCHAUB, 1930 and recent Hungarian Cricetus cricetus L. Folia Hist. Nat. Musei Matra. 1997, 22, 51–79. [Google Scholar]
- Pradel, A. Morphology of the hamster Cricetus cricetus (Linnaeus, 1758) from Poland with some remarks on the evolution of this species. Acta Zool Crac. 1985, 29, 29–52. [Google Scholar]
- Fahlbusch, V. Cricetus Major Woldrich (Mammalia, Rodentia) aus der Mittelpleistozänen Spaltenfüllung Petersbuch 1; Bayerischen Staatssammlung der Paläontologie und Historische Geologie: Munich, Germany, 1976; Volume 16, pp. 71–81. [Google Scholar]
- Hír, J. Cricetus cricetus nanus Schaub, 1930 (Mammalia, Rodentia) finds from the Carpathian Basin. Ann. Hist.-Nat. Musei Natl. Hung. 1994, 86, 13–27. [Google Scholar]
- Hír, J. Cricetids from the Early Pleistocene vertebrate faunha of Somssich-hegy 2 (Southern Hungary, Villány Mountains). Ann. Hist.-Nat. Musei Natl. Hung. 1998, 90, 57–89. [Google Scholar]
- Bellucci, L.; Sardella, R.; Rook, L. Large mammal biochronology framework in Europe at Jaramillo: The Epivillafranchian as a formal biochron. Quat. Int. 2015, 389, 84–89. [Google Scholar] [CrossRef]
- van der Made, J. First description of the large mammals from the locality of Penal, and updated faunal lists for the Atapuerca ungulates—Equus altidens, Bison and human dispersal into Western Europe. Quat. Int. 2013, 295, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Kahlke, R.-D. Late Early Pleistocene European large mammals and the concept of an Epivillafranchian biochron. Cour. Forsch.-Inst. Senckenberg 2007, 259, 265–278. [Google Scholar]
- Carisio, L.; Sacchi, L.; Seglie, D.; Sindaco, R. Habitat selection in the fossorial toad Pelobates fuscus insubricus (Amphibia: Pelobatidae): Does the soil affect species occurrence? Acta Herpetol. 2014, 9, 51–59. [Google Scholar] [CrossRef]
- Böhme, M.; Ilg, A.; Ossig, A.; Küchenhoff, H. New method to estimate paleo precipitation using fossil amphibians and reptiles and the middle and late Miocene precipitation gradients in Europe. Geology 2006, 34, 425–428. [Google Scholar] [CrossRef] [Green Version]
- Agasyan, A.; Avci, A.; Tuniyev, B.; Isailovic, J.C.; Lymberakis, P.; Andrén, C.; Cogalniceanu, D.; Wilkinson, J.; Ananjeva, N.; Üzüm, N.; et al. Telescopus fallax. IUCN Red List. Threat. Species 2009, e.T157258A5062870. [Google Scholar] [CrossRef]
- Haltenorth, T. British and European Mammals, Amphibians, and Reptiles; Irwin & Co., Ltd.: London, UK, 1979; p. 126. [Google Scholar]
- Kovar, R.; Brabec, M.; Vita, R.; Vodicka, R.; Bogdan, V. Habitat use of the Aesculapian snake, Zamenis longissimus at the northern extreme of its range in northwest Bohemia. Herpetol. Bull. 2016, 136, 1–9. [Google Scholar]
- Gál, E.; Hír, J.; Kessler, E.; Kókay, J.; Venczel, M. Középső-Miocén Ősmaradványok a Mátraszőlős, Rákóczi-Kapolna Alatti útbevágásból II. A Mátraszőlős 2. lelőhely [Middle Miocene Fossils from the Road Cut below the Rákóczi Chapel, Mátraszőlős, II. The Mátraszőlős 2. site]. Folia Hist. Nat. Musei Matra. 2000, 24, 39–75. [Google Scholar]
- Hír, J.; Kókay, J.; Venczel, M.; Gál, E.; Kessler, E. Előzetes Jelentés a Felsőtárkányi “Güdör-Kert” Őslénytani Lelőhelykomplex Újravizsgálatáról [Preliminary Report on the Re-Examination of the “Güdör Garden”Paleontological Site Complex in Felsőtárkány]. Folia Hist. Nat. Musei Matra. 2001, 25, 41–64. [Google Scholar]
- Miklas, P.M. Die Amphibienfauna (Amphibia: Caudata, Anura) der Obermiozanen Fundstelle Gotzendorf an der Lietha (sudliches Wiener Becken, Niederosterreich). Ann. Nat. Mus. Wien 2002, 103A, 161–211. [Google Scholar]
- Roček, Z. Late Miocene Amphibia from Rudabánya. Paleontogr. Ital. 2005, 90, 11–29. [Google Scholar]
- Venczel, M.; Hír, J. Lissamphibians and squamata reptiles from the early middle Miocene of Litke, Northern Hungary. Geobios 2015, 48, 491–504. [Google Scholar] [CrossRef]
- Bihari, Z.; Csorba, G.; Heltai, M. (Eds.) Magyarország Emlőseinek Atlasza [Atlas of the Mammals of Hungary]; Kossuth Kiadó: Budapest, Hungary, 2007; pp. 1–360. [Google Scholar]
- Zachos, F.E.; Hackländer, K. (Eds.) Mammals of Europe—Past, Present, and Future, Handbook of the Mammals of Europe; Springer International Publishing: New York, NY, USA, 2020; pp. 1–118. [Google Scholar]
- Botka, D.; Mészáros, L. Taxonomic and palaeoecological review of the Soricidae (Mammalia) fauna from the late Early Pleistocene Somssich Hill 2 locality (Villány Hills, Southern Hungary). Fragm. Palaeontol. Hung. 2018, 35, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Osipova, V.A.; Rzebik-Kowalska, B.; Zaitsev, M.V. Intraspecific variability and phylogenetic relationships of the Pleistocene shrew Sorex runtonensis (Soricidae). Acta Theriol. 2006, 51, 129–138. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Sampling Sites (Meter Distances in the Studied Fissure) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.4–0.0 | 0.8–0.4 | 1.2–0.8 | 1.6–1.2 | 2.0–1.6 | 2.4–2.0 | 2.8–2.4 | 3.2–2.8 | 3.6–3.2 | 4.0–3.6 | 4.4–4.0 | 4.8–4.4 | 5.2–4.8 | 5.6–5.2 | 6.0–5.6 | |
| Triturus cristatus | x | x | x | x | x | ||||||||||
| Lissotriton vulgaris | x | x | x | x | x | x | |||||||||
| Latonia gigantea | x | x | x | ||||||||||||
| Pelobates fuscus | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x |
| Bufo bufo | x | x | |||||||||||||
| Bufotes viridis | x | x | x | x | x | x | x | x | x | ||||||
| Hyla arborea | x | x | |||||||||||||
| Rana temporaria | x | x | x | x | x | x | x | x | |||||||
| Pelophylax esculentus Group | x | x | |||||||||||||
| Lacerta viridis | x | x | x | x | x | ||||||||||
| Anguis fragilis | x | x | x | x | x | x | x | x | x | ||||||
| Hierophis viridiflavus | x | x | x | x | x | ||||||||||
| Hierophis gemonensis | x | x | x | ||||||||||||
| Coronella austriaca | x | x | x | x | x | x | x | x | x | x | x | x | x | ||
| Elaphe cf. paralongissima | x | x | |||||||||||||
| Elaphe cf. quatuorlineata | x | ||||||||||||||
| Zamenis longissimus | x | x | x | x | x | x | x | x | x | x | |||||
| Natrix natrix | x | x | x | x | x | x | x | x | |||||||
| Natrix tesselata | x | x | x | x | x | x | x | x | x | x | |||||
| cf. Telescopus fallax | x | ||||||||||||||
| Vipera cf. ammodytes | x | ||||||||||||||
| Vipera ursinii | x | x | |||||||||||||
| Taxa | Sampling Sites (Meter Distances in the Studied Fissure) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.5–0.0 | 1.0–0.5 | 2.0–1.0 | 2.8–2.4 | 3.2–2.8 | 3.6–3.2 | 4.0–3.6 | 4.4–4.0 | 4.8–4.4 | 5.2–4.8 | 5.6–5.2 | 6.0–5.6 | |
| Erinaceidae gen. et sp. indet. (big-sized) | 1 | |||||||||||
| Talpa sp. indet. | 1 | 1 | ||||||||||
| Desmaninae gen. et sp. indet. | 1 | 1 | 1 | 1 | ||||||||
| Beremendia fissidens | 1 | 2 | 2 | 2 | 2 | 2 | ||||||
| Asoriculus gibberodon | 2 | 1 | ||||||||||
| Sorex (Drepanosorex) savini | 2 | 1 | 2 | 1 | ||||||||
| Sorex runtonensis | 3 | 4 | 2 | 1 | 2 | 1 | 2 | 1 | 1 | |||
| Sorex minutus | 4 | 1 | 2 | 1 | 1 | 1 | 1 | |||||
| Sorex sp. indet. (big-sized) | 1 | |||||||||||
| Spalax sp. | 1 | 1 | 1 | 2 | 1 | 2 | 2 | 2 | 2 | |||
| Cricetus cricetus ssp. | 1 | 2 | 1 | 4 | 3 | 1 | 4 | 2 | 2 | 1 | ||
| Allocricetus bursae | 1 | 1 | 1 | 1 | 2 | 5 | 1 | 2 | 1 | 1 | ||
| Apodemus cf. sylvaticus | 2 | 2 | 2 | 3 | 3 | 2 | 4 | 3 | 2 | 1 | 3 | 2 |
| Sciurus sp. | 1 | 4 | 1 | 2 | ||||||||
| Spermophilus cf. primigenius | 1 | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 2 | 3 | 5 | 4 |
| Eliomys aff. quercinus | 1 | 1 | ||||||||||
| Muscardinus cf. dacicus | 1 | |||||||||||
| Sicista cf. praeloriger | 4 | 2 | 2 | 2 | 2 | 2 | 3 | 6 | 1 | |||
| Ochotona sp. | 1 | 1 | ||||||||||
| Pliomys episcopalis | 1 | 1 | ||||||||||
| Clethrionomys sp. | 1 | 4 | 2 | 1 | ||||||||
| Mimomys pusillus | 2 | 2 | 4 | 4 | 2 | 5 | 1 | 1 | 1 | |||
| Mimomys savini | 1 | 1 | 2 | 2 | 2 | 1 | ||||||
| Lagurodon arankae | 2 | 2 | 5 | 5 | 12 | 4 | 6 | 5 | 3 | 3 | ||
| Prolagurus pannonicus | 3 | 3 | 4 | 10 | 7 | 6 | 3 | 5 | 5 | 3 | 1 | |
| Allophaiomys praehintoni | 2 | 3 | 3 | 7 | 2 | 2 | 3 | 7 | 1 | |||
| Lasiopodomys hintoni | 1 | 2 | 7 | 11 | 5 | 8 | 3 | 2 | 2 | 1 | ||
| Microtus (M.) nivaloides | 1 | 3 | 1 | 4 | 5 | 14 | 6 | |||||
| Microtus (M.) nivalinus | 1 | |||||||||||
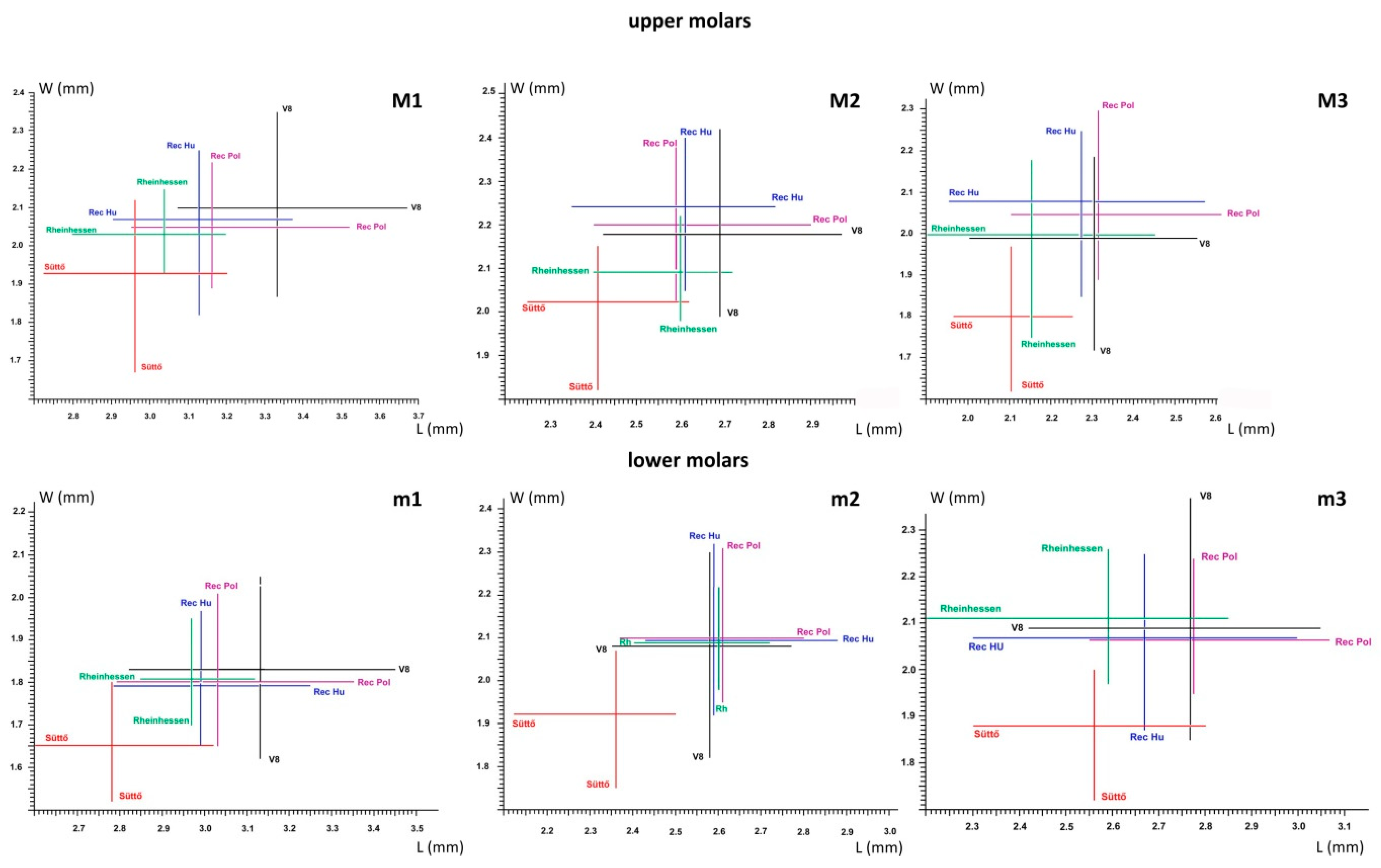
| Cricetus nanus Sites | Tooth Position | L (mm) | W (mm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | min | X | max | n | min | X | max | ||
| Somssich-hegy 2 | M1 | 6 | 2.75 | 2.87 | 2.97 | 6 | 1.75 | 1.84 | 1.92 |
| M2 | 4 | 1.93 | 2.21 | 2.37 | 4 | 1.64 | 1.81 | 1.95 | |
| M3 | 2 | 1.74 | 1.83 | 1.92 | 2 | 1.44 | 1.46 | 1.55 | |
| m1 | 12 | 2.54 | 2.70 | 2.85 | 12 | 1.45 | 1.58 | 1.70 | |
| m2 | 7 | 2.17 | 2.25 | 2.37 | 7 | 1.72 | 1.81 | 1.92 | |
| m3 | 7 | 2.00 | 2.26 | 2.60 | 7 | 1.47 | 1.70 | 1.90 | |
| Betfia 2 | M1 | 12 | 2.85 | 2.81 | 3.0 | 12 | 1.61 | 1.74 | 1.89 |
| M2 | 12 | 2.07 | 2.22 | 2.37 | 12 | 1.71 | 1.81 | 1.96 | |
| M3 | 7 | 1.75 | 1.87 | 1.96 | 7 | 1.55 | 1.62 | 1.68 | |
| m1 | 31 | 2.41 | 2.60 | 2.85 | 31 | 1.39 | 1.52 | 1.68 | |
| m2 | 46 | 1.97 | 2.12 | 2.27 | 46 | 1.54 | 1.71 | 1.88 | |
| m3 | 34 | 2.01 | 2.23 | 2.46 | 34 | 1.48 | 1.71 | 1.95 | |
| Osztramos 14 | M1 | 32 | 2.50 | 2.74 | 3.05 | 32 | 1.58 | 1.72 | 1.87 |
| M2 | 30 | 1.88 | 2.05 | 2.18 | 30 | 1.65 | 1.75 | 1.83 | |
| M3 | 22 | 1.71 | 1.84 | 1.96 | 22 | 1.40 | 1.58 | 1.68 | |
| m1 | 11 | 2.55 | 2.67 | 2.85 | 11 | 1.44 | 1.55 | 1.64 | |
| m2 | 13 | 2.00 | 2.11 | 2.20 | 13 | 1.61 | 1.69 | 1.82 | |
| m3 | 15 | 2.04 | 2.22 | 2.56 | 15 | 1.53 | 1.64 | 1.77 | |
| Osztramos 8 | M1 | 11 | 2.30 | 2.41 | 2.52 | 11 | 1.30 | 1.42 | 1.47 |
| M2 | 8 | 2.00 | 2.12 | 2.31 | 8 | 1.62 | 1.73 | 1.82 | |
| M3 | 3 | 1.64 | 1.74 | 1.81 | 3 | 1.39 | 1.41 | 1.44 | |
| m1 | 18 | 2.30 | 2.41 | 2.52 | 18 | 1.30 | 1.42 | 1.47 | |
| m2 | 17 | 1.88 | 2.09 | 2.17 | 17 | 1.47 | 1.60 | 1.72 | |
| m3 | 13 | 1.82 | 2.05 | 2.24 | 13 | 1.44 | 1.55 | 1.64 | |
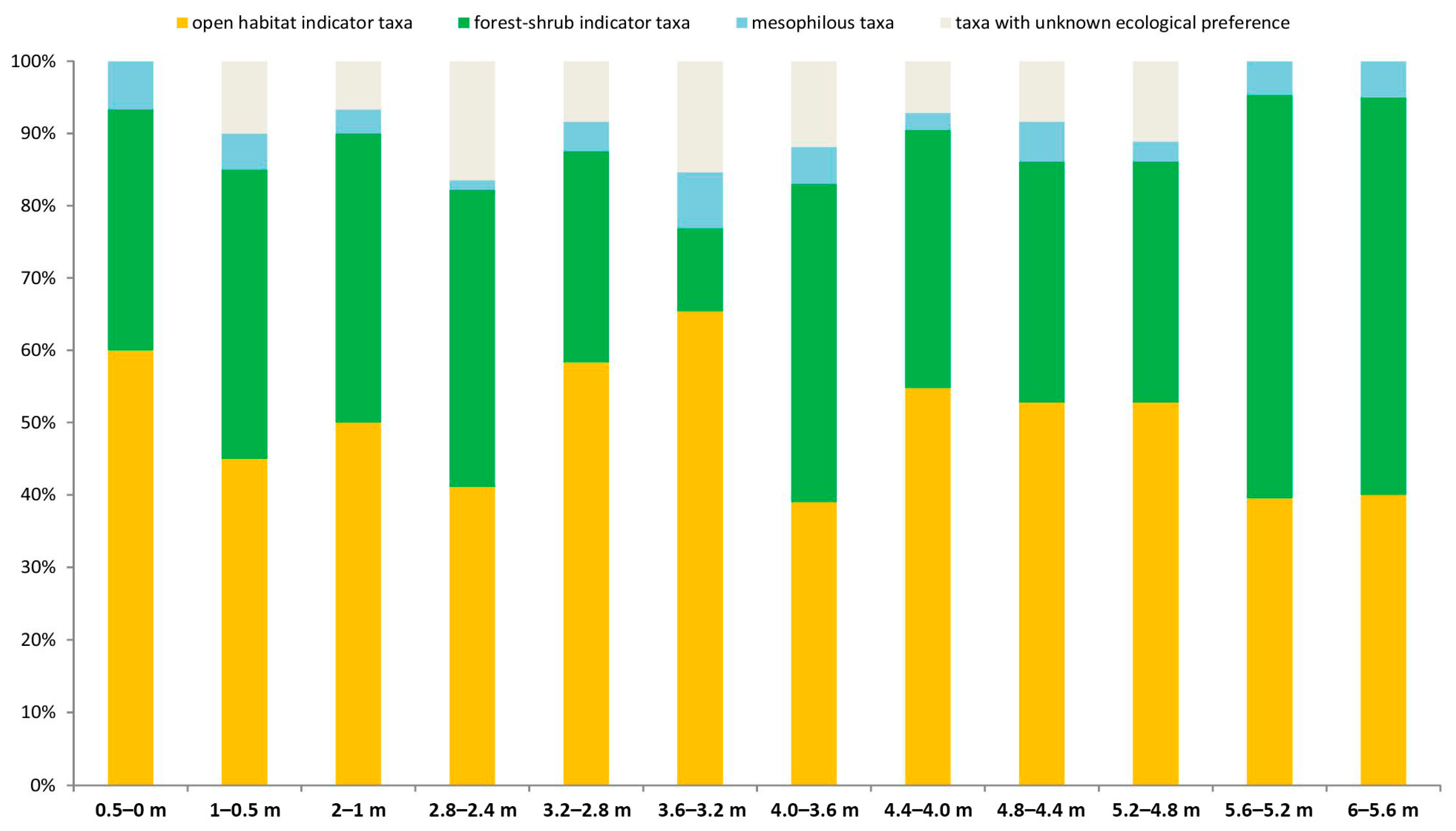
| Open Habitat Indicator Taxa | Forest-Shrub Indicator Taxa |
|---|---|
| Sorex runtonensis | Talpa sp. indet. |
| Spalax sp. | Asoriculus gibberodon |
| Cricetus cricetus ssp. | Sorex minutus |
| Allocricetus bursae | Sorex sp. indet. (big sized) |
| Spermophilus cf. primigenius | Apodemus cf. sylvaticus |
| Sicista cf. praeloriger | Sciurus sp. |
| Ochotona sp. | Eliomys aff. quercinus |
| Lagurodon arankae | Muscardinus cf. dacicus |
| Prolagurus pannonicus | Clethrionomys sp. |
| Allophaiomys praehintoni | |
| Lasiopodomys hintoni | |
| Microtus (M.) nivaloides | |
| Pliomys episcopalis | |
| mesophilous taxa | taxa with unknown ecological preference |
| Desmaninae gen et sp. indet. | Beremendia fissidens |
| Sorex (Drepanosorex) savini | Mimomys pusillus |
| Mimomys savini |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pazonyi, P.; Szentesi, Z.; Mészáros, L.; Hír, J.; Gasparik, M. Stratigraphic and Paleoecological Significance of the Early/Middle Pleistocene Vertebrate Fauna of the Süttő 21 Site. Diversity 2023, 15, 736. https://doi.org/10.3390/d15060736
Pazonyi P, Szentesi Z, Mészáros L, Hír J, Gasparik M. Stratigraphic and Paleoecological Significance of the Early/Middle Pleistocene Vertebrate Fauna of the Süttő 21 Site. Diversity. 2023; 15(6):736. https://doi.org/10.3390/d15060736
Chicago/Turabian StylePazonyi, Piroska, Zoltán Szentesi, Lukács Mészáros, János Hír, and Mihály Gasparik. 2023. "Stratigraphic and Paleoecological Significance of the Early/Middle Pleistocene Vertebrate Fauna of the Süttő 21 Site" Diversity 15, no. 6: 736. https://doi.org/10.3390/d15060736