
The Exceptional Presence of Megaloceros giganteus in North-Eastern Iberia and Its Palaeoecological Implications: The Case of Teixoneres Cave (Moià, Barcelona, Spain)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Teixoneres Cave
2.2. Methods
2.2.1. Linear Morphometry and Biogeography
2.2.2. Dental Wear Analysis
3. Results
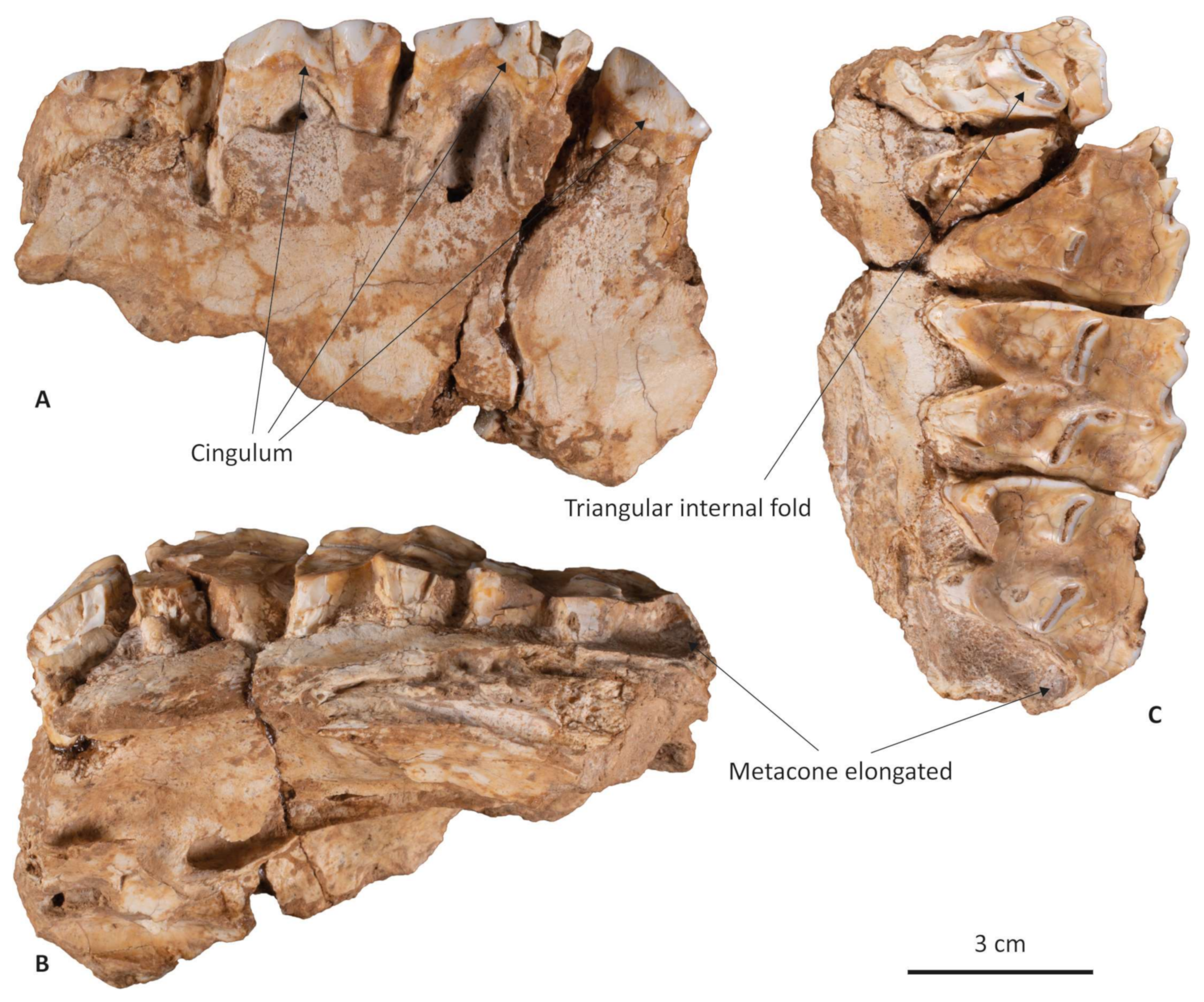
3.1. Morphometric Description of the Teixoneres Megaloceros Giganteus Maxillary
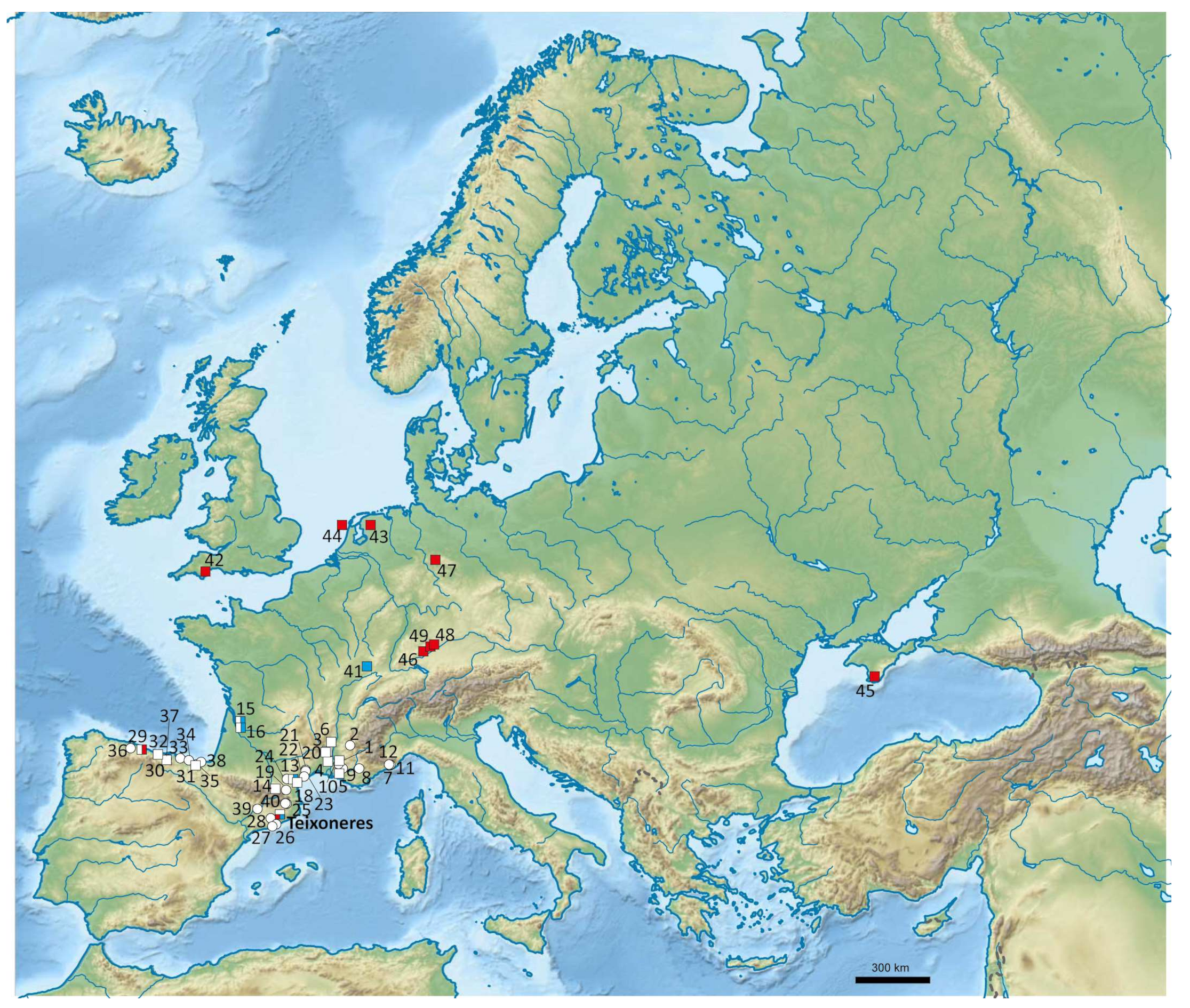
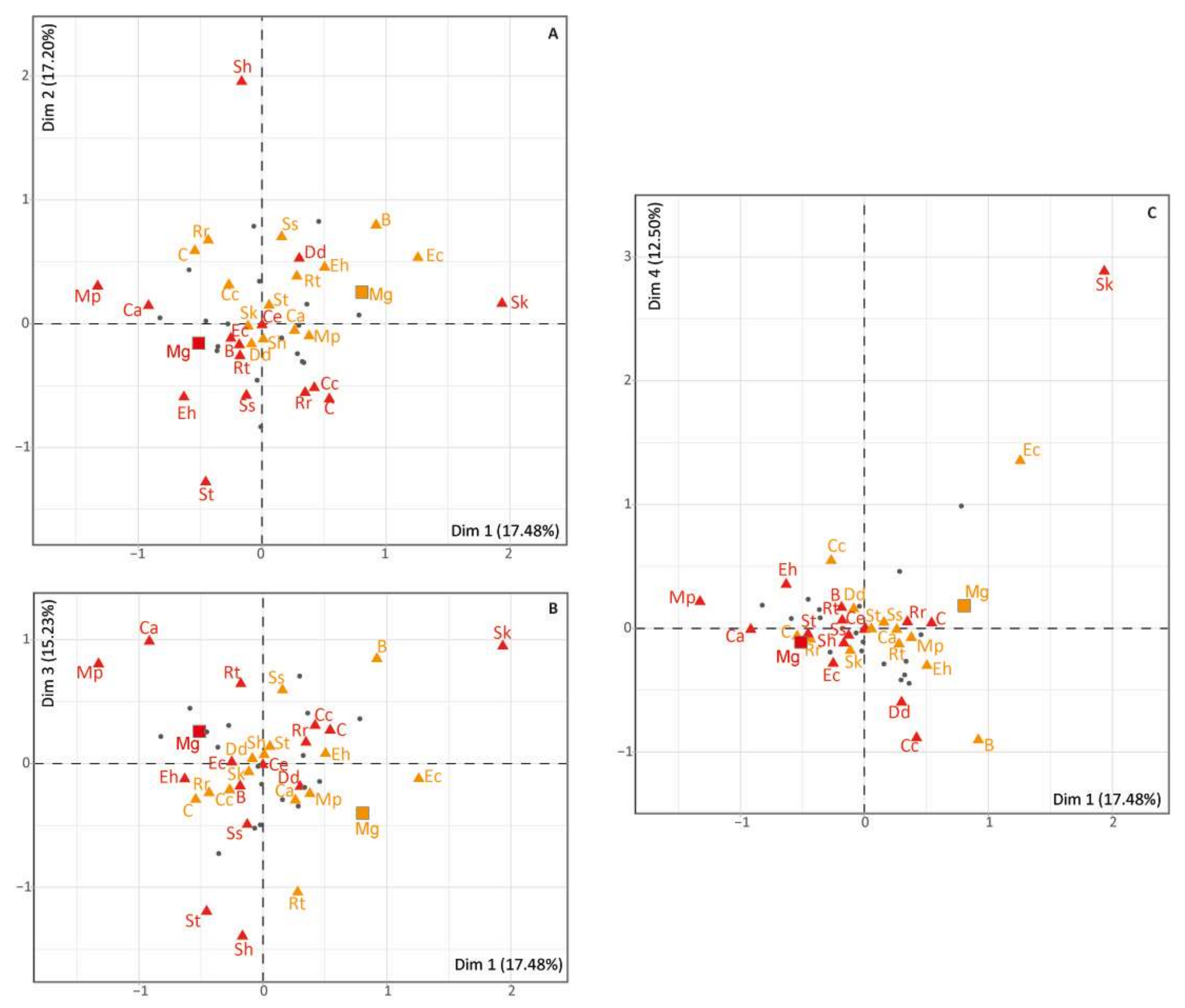
3.2. Co-Occurrence of Megaloceros with other Herbivores North and South of the Pyrenees
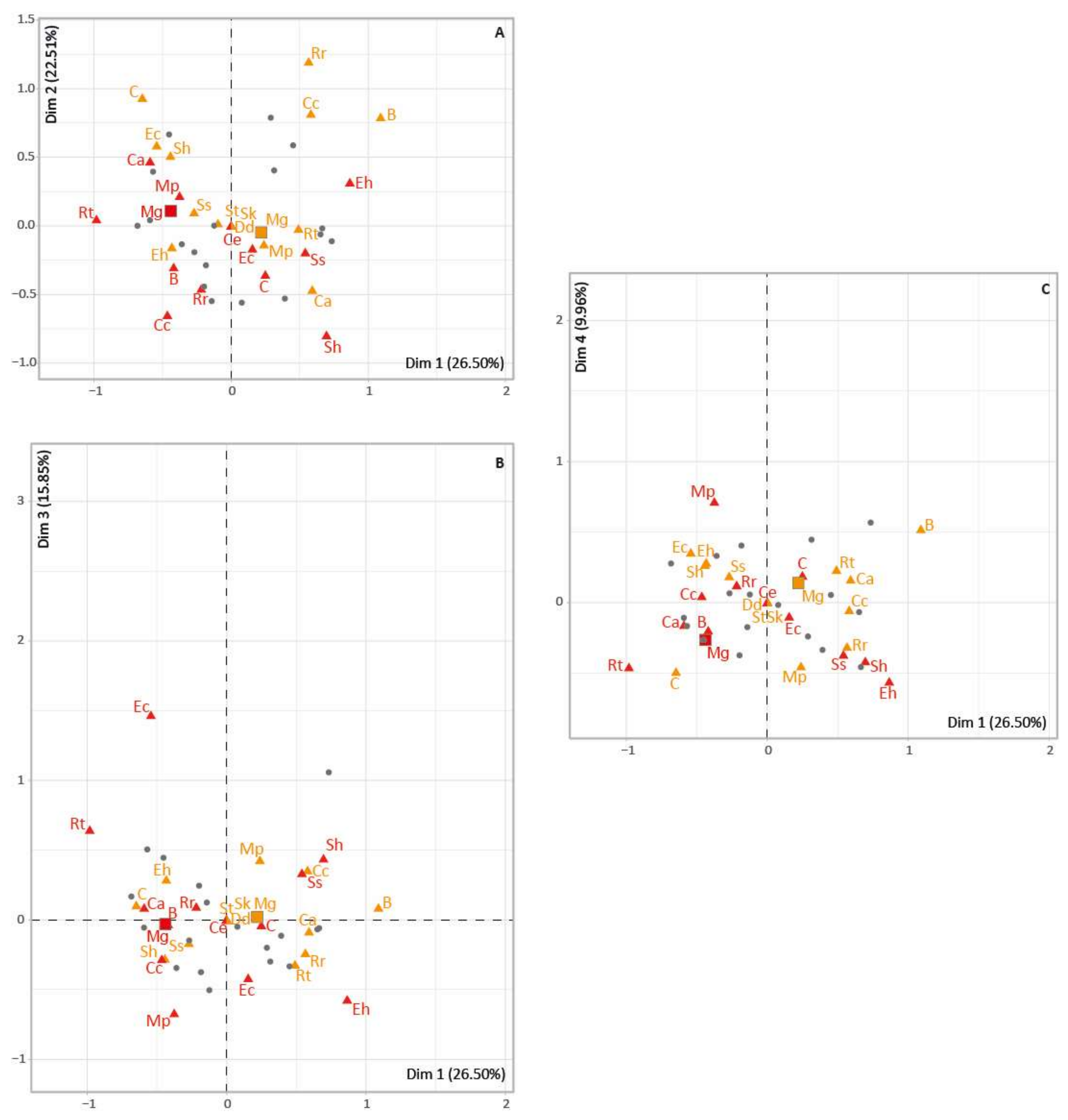
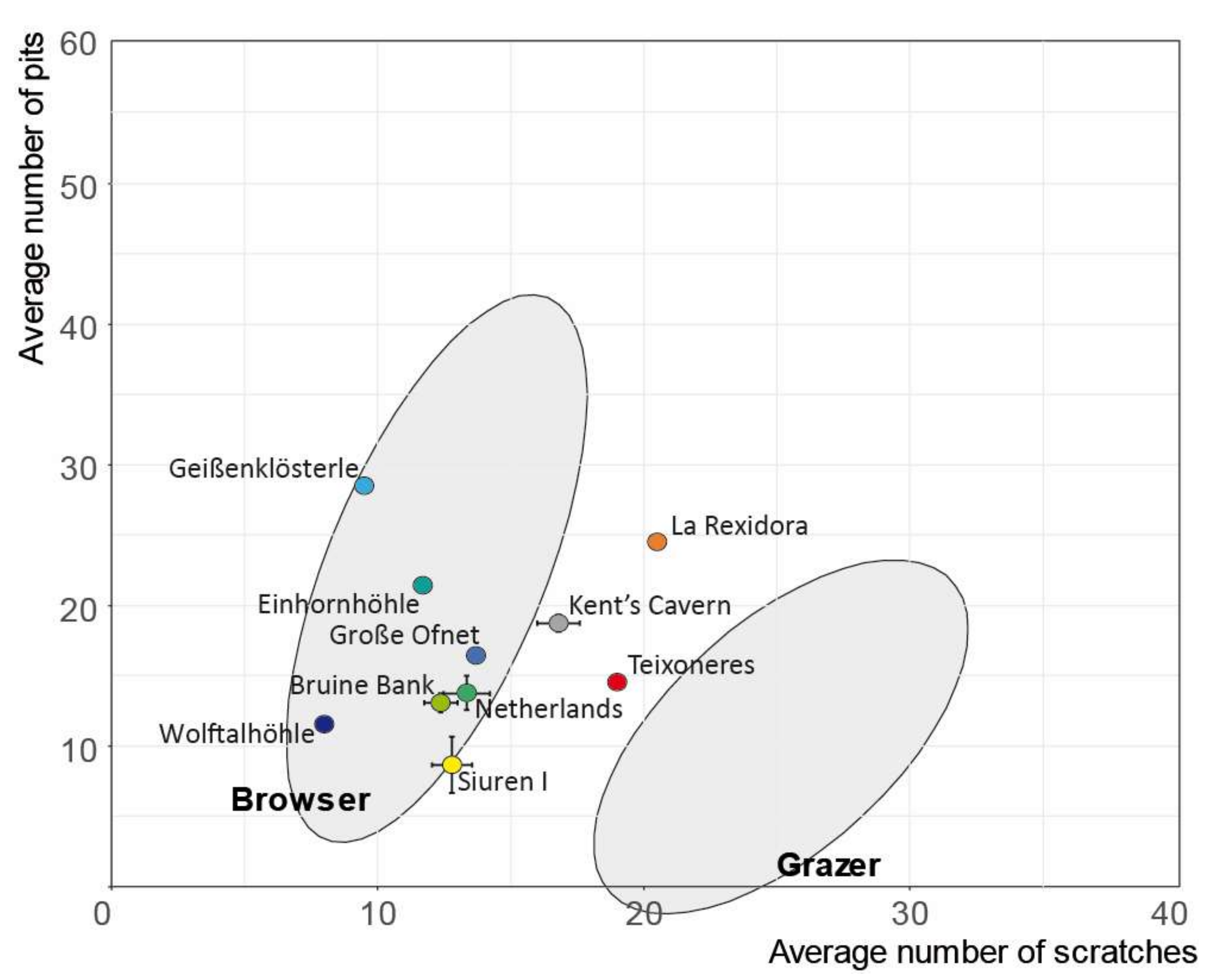
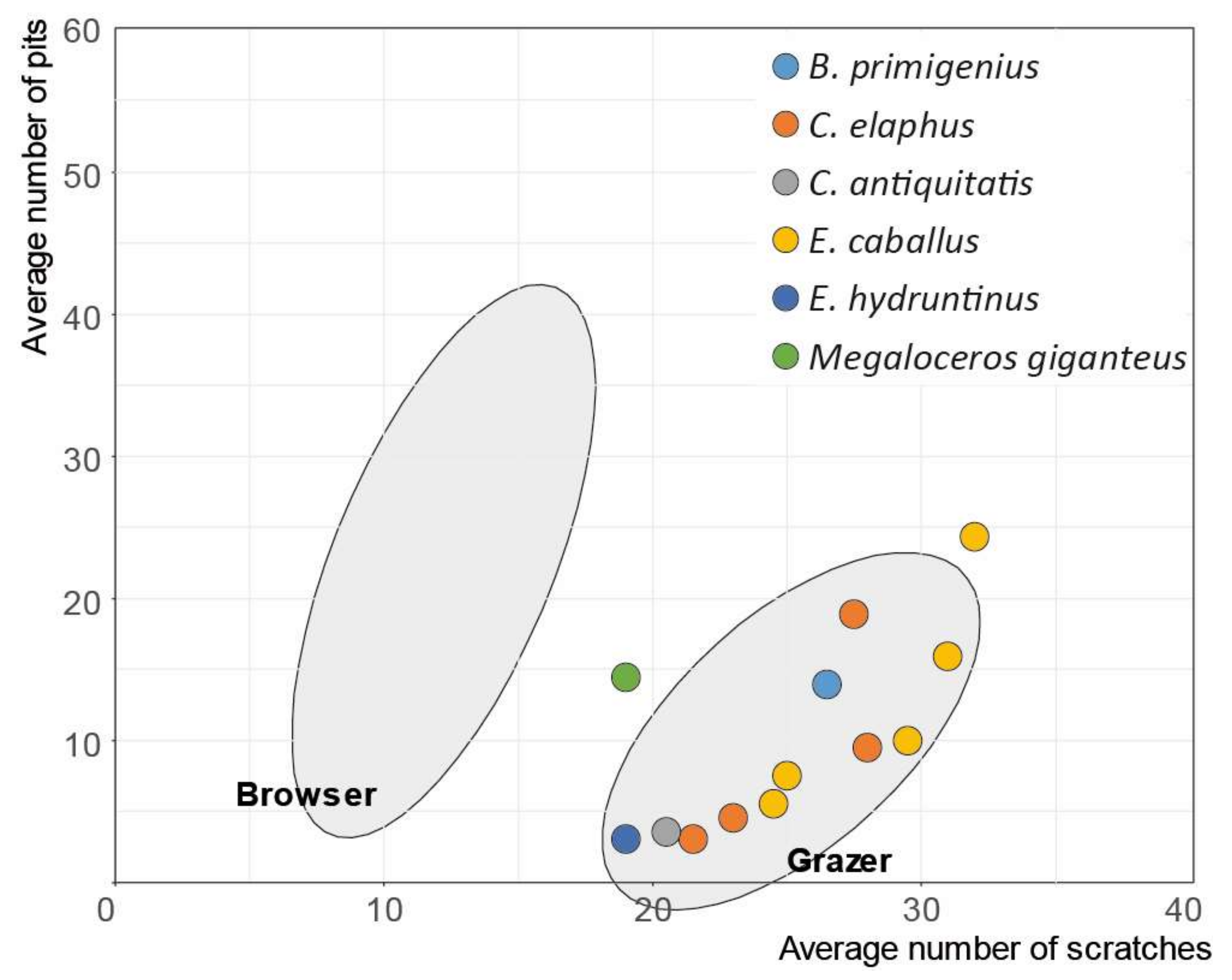
3.3. Megaloceros Dietary Habits
4. Discussion
4.1. Taxonomic Attribution of the Teixoneres Megaloceros Specimen
4.2. Did Megaloceros Avoid/Cohabit with the Same Herbivore Species North and South of the Pyrenees?
4.3. Megaloceros Dietary Niches from North to South
4.4. Synthesis: Which Conditions Favoured the Megaloceros Incursions into the Iberian Peninsula?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lister, A.M. The Evolution of the Giant Deer, Megaloceros Giganteus (Blumenbach). Zool. J. Linn. Soc. 1994, 112, 65–100. [Google Scholar] [CrossRef]
- Stuart, A.J.; Kosintsev, P.A.; Higham, T.F.G.; Lister, A.M. Pleistocene to Holocene Extinction Dynamics in Giant Deer and Woolly Mammoth. Nature 2004, 431, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Van der Plicht, J.; Molodin, V.I.; Kuzmin, Y.V.; Vasiliev, S.K.; Postnov, A.V.; Slavinsky, V.S. New Holocene Refugia of Giant Deer (Megaloceros Giganteus Blum.) in Siberia: Updated Extinction Patterns. Quat. Sci. Rev. 2015, 114, 182–188. [Google Scholar] [CrossRef]
- Lister, A.M.; Stuart, A.J. The Extinction of the Giant Deer Megaloceros Giganteus (Blumenbach): New Radiocarbon Evidence. Quat. Int. 2019, 500, 185–203. [Google Scholar] [CrossRef]
- Bouchud, J. Le Cervus megaceros Dans Le Sud et Le Sud-Ouest de La France. Isr. J. Zool. 1965, 14, 24–37. [Google Scholar] [CrossRef]
- Brochier, J.É. Evolution Des Climats et Des Paysages Vauclusiens Au Cours Du Würmien Récent et Du Postglaciaire En Vaucluse; Paléoécologie de l’Homme Fossile; CNRS Éditions: Paris, France, 1977. [Google Scholar]
- Crégut-Bonnoure, E.; Paccard, M. La faune à Saïga tatarica de Chinchon I (Saumanes, Vaucluse). Quaternaire 1997, 8, 391–407. [Google Scholar] [CrossRef]
- Magniez, P. Etude Paléontologique Des Artiodactyles de la Grotte Tournal (Bize-Minervois, Aude, France) Étude Taphonomique, Archéozoologique et Paléoécologique Des Grands Mammifères Dans Leur Cadre Biostratigraphique et Paléoenvironnemental. Thèse de Doctorat, Université de Perpignan, Perpignan, France, 2010. [Google Scholar]
- Athanassiou, A. The Fossil Record of Continental Fossil Deer (Mammalia: Artiodactyla: Cervidae) in Greece. In Fossil Vertebrates of Greece Vol. 2: Laurasiatherians, Artiodactyles, Perissodactyles, Carnivorans, and Island Endemics; Vlachos, E., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 205–247. ISBN 978-3-030-68442-6. [Google Scholar]
- Álvarez-Lao, D.J.; García, N. Geographical Distribution of Pleistocene Cold-Adapted Large Mammal Faunas in the Iberian Peninsula. Quat. Int. 2011, 233, 159–170. [Google Scholar] [CrossRef]
- Álvarez-Lao, D.J. The Jou Puerta Cave (Asturias, NW Spain): A MIS 3 Large Mammal Assemblage with Mixture of Cold and Temperate Elements. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 393, 1–19. [Google Scholar] [CrossRef]
- Koch, P.L.; Barnosky, A.D. Late Quaternary Extinctions: State of the Debate. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 215–250. [Google Scholar] [CrossRef] [Green Version]
- Lorenzen, E.D.; Nogués-Bravo, D.; Orlando, L.; Weinstock, J.; Binladen, J.; Marske, K.A.; Ugan, A.; Borregaard, M.K.; Gilbert, M.T.P.; Nielsen, R.; et al. Species-Specific Responses of Late Quaternary Megafauna to Climate and Humans. Nature 2011, 479, 359–364. [Google Scholar] [CrossRef] [Green Version]
- Bocherens, H. The Rise of the Anthroposphere since 50,000 Years: An Ecological Replacement of Megaherbivores by Humans in Terrestrial Ecosystems? Front. Ecol. Evol. 2018, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Rey-Iglesia, A.; Lister, A.M.; Campos, P.F.; Brace, S.; Mattiangeli, V.; Daly, K.G.; Teasdale, M.D.; Bradley, D.G.; Barnes, I.; Hansen, A.J. Exploring the Phylogeography and Population Dynamics of the Giant Deer (Megaloceros giganteus) Using Late Quaternary Mitogenomes. Proc. R. Soc. B Biol. Sci. 2021, 288, 20201864. [Google Scholar] [CrossRef]
- Immel, A.; Drucker, D.G.; Bonazzi, M.; Jahnke, T.K.; Münzel, S.C.; Schuenemann, V.J.; Herbig, A.; Kind, C.-J.; Krause, J. Mitochondrial Genomes of Giant Deers Suggest Their Late Survival in Central Europe. Sci. Rep. 2015, 5, 10853. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Lao, D.J.; Rivals, F.; Sánchez-Hernández, C.; Blasco, R.; Rosell, J. Ungulates from Teixoneres Cave (Moià, Barcelona, Spain): Presence of Cold-Adapted Elements in NE Iberia during the MIS 3. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 466, 287–302. [Google Scholar] [CrossRef]
- Rosell, J.; Blasco, R.; Cebrià, A.; Chacón, M.G.; Menendez, L.; Morales, J.I.; Rodriguez-Hidalgo, A. Mossegades i Levallois: Les Noves Intervencionsa La Cova de Les Teixoneres (Moià, Bages). Trib D’arqueologia 2008, 29–43. [Google Scholar]
- Rosell, J.; López, R.B.; Rivals, F.; Artur, C.I.E.; Hidalgo, J.I.M.; Rodríguez, A.; Serrat, D.; Roura, E.C. Los ocupaciones en la Cova de les Teixoneres (Moià, Barcelona): Relaciones espaciales y grado de competencia entre hienas, osos y neandertales durante el Pleistoceno Superior. In Proceedings of the Actas de la 1a Reunión de Científicos Sobre Cubiles de Hiena (y Otros Grandes Carnívoros) en los Yacimientos Arqueológicos de la Península Ibérica; Arriaza, M.C., Pérez, E.B., Rosell, J., Eds.; Museo Arqueológico Regional: Madrid, Spain, 2010; pp. 392–402. [Google Scholar]
- Rosell, J.; Blasco, R.; Rivals, F.; Chacón, M.G.; Menéndez, L.; Morales, J.; Rodriguez-Hidalgo, A.; Cebria, A.; Carbonell, E.; Serrat, D. A Stop along the Way: The Role of Neanderthal Groups at Level III of Teixoneres Cave (Moià, Barcelona, Spain). Quaternaire 2010, 21, 139–154. [Google Scholar]
- Rosell, J.; Blasco, R.; Rivals, F.; Chacón, M.G.; Blain, H.-A.; Lopez Garcia, J.M.; Picin, A.; Camarós, E.; Rufà, A.; Sanchez-Hernandez, A.M.; et al. Cova Del Toll y Cova de Les Teixoneres (Moià, Barcelona). In Los Cazadores Recolectores del Pleistoceno y del Holoceno en Iberia y el Estrecho de Gibraltar; Sala, R., Carbonell, E., Bermudez de Castro, J.M., Arsuaga, J.L., Eds.; University of Burgos: Burgos, Spain, 2014; pp. 302–307. [Google Scholar]
- Zilio, L.; Hammond, H.; Karampaglidis, T.; Sánchez-Romero, L.; Blasco, R.; Rivals, F.; Rufà, A.; Picin, A.; Chacón, M.G.; Demuro, M.; et al. Examining Neanderthal and Carnivore Occupations of Teixoneres Cave (Moià, Barcelona, Spain) Using Archaeostratigraphic and Intra-Site Spatial Analysis. Sci. Rep. 2021, 11, 4339. [Google Scholar] [CrossRef]
- Talamo, S.; Blasco, R.; Rivals, F.; Picin, A.; Chacón, M.G.; Iriarte, E.; López-García, J.M.; Blain, H.-A.; Arilla, M.; Rufà, A.; et al. The Radiocarbon Approach to Neanderthals in a Carnivore Den Site: A Well-Defined Chronology for Teixoneres Cave (Moià, Barcelona, Spain). Radiocarbon 2016, 58, 247–265. [Google Scholar] [CrossRef]
- Ochando, J.; Carrión, J.S.; Blasco, R.; Rivals, F.; Rufà, A.; Demuro, M.; Arnold, L.J.; Amorós, G.; Munuera, M.; Fernández, S.; et al. Neanderthals in a Highly Diverse, Mediterranean-Eurosiberian Forest Ecotone: The Pleistocene Pollen Record of Teixoneres Cave, Northeastern Spain. Quat. Sci. Rev. 2020, 241, 106429. [Google Scholar] [CrossRef]
- Rufà, A.; Blasco, R.; Rivals, F.; Rosell, J. Who Eats Whom? Taphonomic Analysis of the Avian Record from the Middle Paleolithic Site of Teixoneres Cave (Moià, Barcelona, Spain). Quat. Int. 2016, 421, 103–115. [Google Scholar] [CrossRef]
- López-García, J.M.; Blain, H.-A.; Burjachs, F.; Ballesteros, A.; Allué, E.; Cuevas-Ruiz, G.E.; Rivals, F.; Blasco, R.; Morales, J.I.; Hidalgo, A.R.; et al. A Multidisciplinary Approach to Reconstructing the Chronology and Environment of Southwestern European Neanderthals: The Contribution of Teixoneres Cave (Moià, Barcelona, Spain). Quat. Sci. Rev. 2012, 43, 33–44. [Google Scholar] [CrossRef]
- Picin, A.; Chacón, M.G.; Gómez de Soler, B.; Blasco, R.; Rivals, F.; Rosell, J. Neanderthal Mobile Toolkit in Short-Term Occupations at Teixoneres Cave (Moia, Spain). J. Archaeol. Sci. Rep. 2020, 29, 102165. [Google Scholar] [CrossRef]
- Fernández-García, M.; López-García, J.M.; Royer, A.; Lécuyer, C.; Rivals, F.; Rufà, A.; Blasco, R.; Rosell, J. New Insights in Neanderthal Palaeoecology Using Stable Oxygen Isotopes Preserved in Small Mammals as Palaeoclimatic Tracers in Teixoneres Cave (Moià, Northeastern Iberia). Archaeol Anthr. Sci. 2022, 14, 106. [Google Scholar] [CrossRef]
- Uzunidis, A.; Rufà, A.; Blasco, R.; Rosell, J.; Brugal, J.-P.; Texier, P.-J.; Rivals, F. Speciated Mechanism in Quaternary Cervids (Cervus and Capreolus) on Both Sides of the Pyrenees: A Multidisciplinary Approach—Scientific Reports. Sci. Rep. 2022, 12, 20200. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Brugal, J.-P. Apport à La Chronologie Du Würm Récent Dans Les Alpes Du Sud: La Faune à Mégacéros Du Pignon (Aspres-Sur-Büech, Hautes-Alpes). Préhistoire Anthropol. Méditerranéennes 1993, 2, 29–42. [Google Scholar]
- Gamberi, L.; Argant, A.; Argant, J.; Barth, P.; Boudadi-Maligne, M.; Boulbes, N.; Brugal, J.-P.; Caramelli, D.; Condémi, S.; Crégut-Bonnoure, E.; et al. L’aven de l’Arquet—Barjac (30) Étude d’un Aven Piège. Ardèche Archéologie 2011, 28, 3–10. [Google Scholar]
- Gerber, J.P. La Faune Des Grands Mammifères Du Würm Ancien Dans Le Sud-Est de La France. Ph.D. Thesis, Université de Provence, Marseille, France, 1973, unpublished. [Google Scholar]
- Bonifay, M.-F.; Lecourtois-Ducgoninaz, A. La Faune Des Niveaux Inférieurs de l’Abri Cornille à Istres (Bouches-Du-Rhône); Congrès Préhistorique de France: Provence, France, 1976; pp. 228–246. [Google Scholar]
- Daujeard, C. Exploitation Du Milieu Animal Par Les Néanderthaliens Dans Le Sud-Est de La France. Thèse de Doctorat, Université Lumière, Lyon, France, 2008. [Google Scholar]
- Brugal, J.-P.; Fourvel, J.-B.; Fosse, P. Premières Observations Sur Les Guildes de Carnivores de La Grotte de l’Observatoire (Principauté de Monaco). Bull. Du Musée D’anthropologie Préhistorique De Monaco 2017, 57, 35–49. [Google Scholar]
- Monchot, H. Les Chasseurs Tardigravettiens de La Baume de Goulon (Salernes, Var, France). Rev. De Paleobiol. 2008, 27, 409–427. [Google Scholar]
- Moussous, A. Approche Biostratigraphique et Evolution Des Paléoenvironnements Au Cours Du Pléistocène Supérieur, d’après l’étude Des Grands Mammifères Des Grottes de Balzi Rossi (Ligurie, Italie). Ph.D. Thesis, Perpignan University, Perpignan, France, 2014, unpublished. [Google Scholar]
- Prat, F. Sur Les Vestiges de Megaceros Découverts Dans Quelques Gisements Paléolithiques Du Sud-Ouest de La France. Thèse D’état, Bordeaux University, Bordeaux, France, 1968, unpublished. [Google Scholar]
- Guadelli, J.-L. Etude taphonomique du repaire d’hyènes de Camiac (Gironde, France). Eléments de comparaison entre un site naturel et un gisement préhistorique. Bull. De L’association Française Pour L’étude Du Quat. 1989, 2, 91–100. [Google Scholar] [CrossRef]
- Rusch, L. Comportements de Subsistance et Environnements Des Néandertaliens En Languedoc Au Pléistocène Supérieur: L’apport Des Grands Mammifères de l’archéoséquence Des Ramandils (Port-La-Nouvelle, Aude, France). Ph.D. Thesis, Perpignan University, Perpignan, France, 2020, unpublished. [Google Scholar]
- Saos, T.; Gregoire, S.; Moigne, A.-M.; Testu, A.; Boulbes, N.; Bachellerie, M.; Alladio, A.; Ortega-Cordellat, M.-I.; Veall, M.-A.; Deviese, T.; et al. Grotte de la Crouzade (Gruissan, Aude), Rapport de Fouille Programmée Triennale 2016–2018, Rapport Final 2018; Service Régional de l’Archéologie: Montpellier, France, 2018; p. 125. [Google Scholar]
- Fontana, L. Mobilité et Subsistance Au Magdalénien Dans Le Bassin de l’Aude. Bull. De La Société Préhistorique Française 1999, 96, 175–190. [Google Scholar] [CrossRef]
- Rillardon, M. Environnement et Subsistance Des Derniers Chasseurs-Cueilleurs Dans La Basse Vallée Du Rhône et Ses Marges Du Pléniglaciaire Supérieur (20 Ka BP) à l’optimum Climatique (8 Ka BP). These de Doctorat, Aix-Marseille 1, Aix-en Provence, France, 2010. [Google Scholar]
- Galobart, A.; Maroto, J.; Ros, X. Las Faunas Cuaternarias de Mamíferos de La Cuenca de Banyoles-Besalú (Girona). Rev. Española De Paleontol. Numero Extraordin. 1996, 11, 248–255. [Google Scholar] [CrossRef]
- Daura, J.; Sanz, M.; García, N.; Allué, E.; Vaquero, M.; Fierro, E.; Carrión, J.S.; López-García, J.M.; Blain, H.A.; Sánchez-Marco, A.; et al. Terrasses de La Riera Dels Canyars (Gavà, Barcelona): The Landscape of Heinrich Stadial 4 North of the “Ebro Frontier” and Implications for Modern Human Dispersal into Iberia. Quat. Sci. Rev. 2013, 60, 26–48. [Google Scholar] [CrossRef] [Green Version]
- Daura, J.; Sanz, M.; Vaquero, M.; López-García, J.M.; Blain, H.-A.; Marco, A.S. Neanderthal Fossils, Mobile Toolkit and a Hyena Den: The Archaeological Assemblage of Lateral Gallery 1 in Cova Del Gegant (NE Iberian Peninsula). Quaternary 2022, 5, 12. [Google Scholar] [CrossRef]
- Caceres, I.; Rosell, J.; Huguet, R. Séquence d’utilisation de la biomasse animale dans le gisement de l’Abric Romani (Barcelone, Espagne). Quaternaire 1998, 9, 379–383. [Google Scholar] [CrossRef]
- Álvarez-Lao, D.J.; Ruiz-Zapata, M.B.; Gil-García, M.J.; Ballesteros, D.; Jiménez-Sánchez, M. Palaeoenvironmental Research at Rexidora Cave: New Evidence of Cold and Dry Conditions in NW Iberia during MIS 3. Quat. Int. 2015, 379, 35–46. [Google Scholar] [CrossRef]
- Rivals, F.; Álvarez-Lao, D.J. Ungulate Dietary Traits and Plasticity in Zones of Ecological Transition Inferred from Late Pleistocene Assemblages at Jou Puerta and Rexidora in the Cantabrian Region of Northern Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 499, 123–130. [Google Scholar] [CrossRef]
- Castaños Ugarte, P.M. El Castillo, Historia de Una Fauna Olvidada: La Fauna de Las Excavaciones Históricas (1910–1914) de la Cueva de “El Castillo” (Puente Viesgo, Cantabria); Consejería de Educación, Cultura y Deportes: Toledo, Spain, 2018; ISBN 978-84-697-8714-4. [Google Scholar]
- Rios-Garaizar, J.; Arrizabalaga, A.; Villaluenga, A. Haltes de chasse du Châtelperronien de la Péninsule Ibérique: Labeko Koba et Ekain (Pays Basque Péninsulaire). L’Anthropologie 2012, 116, 532–549. [Google Scholar] [CrossRef]
- Castaños Ugarte, P.M. Hallazgos de rinoceronte lanudo en Legintxiki (Etxauri, Navarra). Príncipe De Viana. Supl. De Cienc. 1996, 77–80. [Google Scholar]
- Castaños Ugarte, P.M.; Bereicua, X.M.; Bailon, S.; de la Fuente, J.C.; de Lafuente, X.S.; Hernando, O.S. Estudio de los vertebrados del yacimiento de Lezizako Koba (Kortezubi, Bizkaia). Kobie. Paleoantropol. 2009, 25–50. [Google Scholar]
- Altuna, J. Fauna de Mamíferos de Los Yacimientos Prehistóricos de Guipúzcoa, Con Catalogo de Los Mamíferos Cuaternarios Del Cantábrico y Del Pirineo Occidental. Munibe 1972, 24, 1–464. [Google Scholar]
- Altuna, J.; Mariezkurrena, K. Macromamíferos del yacimiento de Labeko Koba (Arrasate, País Vasco). Munibe 2000, 52, 107–181. [Google Scholar]
- Álvarez-Lao, D.J.; Méndez, M. Latitudinal Gradients and Indicator Species in Ungulate Paleoassemblages during the MIS 3 in W Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 449, 455–462. [Google Scholar] [CrossRef]
- Azanza, B.; Baldellou, V.; Cuchi, J.A.; López, P.; Montes, L.; Utrilla, P. Cronoestratigrafía de la cueva musteriense de Los Moros (Gabasa, Huesca). Cuatern. Y Geomorfol. 1988, 2, 1–12. [Google Scholar]
- Estévez-Escalera, J. La Fauna Del Pleistoceno Catalan. Ph.D. Thesis, Barcelona University, Barcelona, Spain, 1979, unpublished. [Google Scholar]
- Nedeljković, D. Lobanja Ženke Pleistocenskog Jelena Iz Korita Reke Save (Vojvodina, Srbija) [The Skull of Female Pleistocene Deer from the Riverbed of the Sava River (Vojvodina, Serbia)]. Actes Mus. Srem 2021, 11, 1–20. [Google Scholar]
- Fourvel, J.-B.; Fosse, P.; Fernandez, P.; Antoine, P.-O. Large Mammals of Fouvent-Saint-Andoche (Haute-Saône, France): A Glimpse into a Late Pleistocene Hyena Den. Geodiversitas 2015, 37, 237–266. [Google Scholar] [CrossRef]
- Rivals, F.; Lister, A.M. Dietary Flexibility and Niche Partitioning of Large Herbivores through the Pleistocene of Britain. Quat. Sci. Rev. 2016, 146, 116–133. [Google Scholar] [CrossRef]
- Rivals, F.; Mihlbachler, M.C.; Solounias, N.; Mol, D.; Semprebon, G.M.; de Vos, J.; Kalthoff, D.C. Palaeoecology of the Mammoth Steppe Fauna from the Late Pleistocene of the North Sea and Alaska: Separating Species Preferences from Geographic Influence in Paleoecological Dental Wear Analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 286, 42–54. [Google Scholar] [CrossRef]
- Ramírez-Pedraza, I.; Rivals, F.; Uthmeier, T.; Chabai, V. Palaeoenvironmental and Seasonal Context of the Late Middle and Early Upper Palaeolithic Occupations in Crimea: An Approach Using Dental Wear Patterns in Ungulates. Archaeol Anthr. Sci. 2020, 12, 268. [Google Scholar] [CrossRef]
- Rivals, F.; Drucker, D.G.; Starkovich, B.M.; Russo, G.; Leder, D.; Conard, N.J. Dental Wear Evidence for Browsing and Grazing Dietary Traits in the Giant Deer from the Late Pleistocene of Central Europe. MGfU 2021, 30, 95–113. [Google Scholar] [CrossRef]
- Lister, A.M.; Edwards, C.J.; Nock, D.A.W.; Bunce, M.; van Pijlen, I.A.; Bradley, D.G.; Thomas, M.G.; Barnes, I. The Phylogenetic Position of the ‘Giant Deer’ Megaloceros giganteus. Nature 2005, 438, 850–853. [Google Scholar] [CrossRef]
- Abdi, H.; Valentin, D. Multiple Correspondence Analysis. In Encyclopedia of Measurement and Statistics; Salkind, N.J., Ed.; SAGE Publications: Thousand Oaks, CA, USA, 2007; pp. 651–656. [Google Scholar]
- Arruda Silva, M.; Canezin Guideli, L.; Chaves Neto, A.; de Brum Passini, L.; Morales Kormann, A.C. Multiple Correspondence Analysis Applied to the Study of the Relationship between Traffic Crashes and Precipitation on a Highway in Brazil. Transportes 2020, 28, 196–211. [Google Scholar] [CrossRef]
- Grine, F.E. Dental Evidence for Dietary Differences in Australopithecus and Paranthropus: A Quantitative Analysis of Permanent Molar Microwear. J. Hum. Evol. 1986, 15, 783–822. [Google Scholar] [CrossRef]
- Teaford, M.F.; Oyen, O.J. In Vivo and in Vitro Turnover in Dental Microwear. Am. J. Phys. Anthropol. 1989, 80, 447–460. [Google Scholar] [CrossRef]
- Winkler, D.E.; Schulz-Kornas, E.; Kaiser, T.M.; Codron, D.; Leichliter, J.; Hummel, J.; Martin, L.F.; Clauss, M.; Tütken, T. The Turnover of Dental Microwear Texture: Testing the” Last Supper” Effect in Small Mammals in a Controlled Feeding Experiment. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 557, 109930. [Google Scholar] [CrossRef]
- Solounias, N.; Semprebon, G. Advances in the Reconstruction of Ungulate Ecomorphology with Application to Early Fossil Equids. Am. Mus. Novit. 2002, 3366, 49. [Google Scholar] [CrossRef]
- Semprebon, G.; Godfrey, L.R.; Solounias, N.; Sutherland, M.R.; Jungers, W.L. Can Low-Magnification Stereomicroscopy Reveal Diet? J. Hum. Evol. 2004, 47, 115–144. [Google Scholar] [CrossRef]
- Walker, A.; Hoeck, H.N.; Perez, L. Microwear of Mammalian Teeth as an Indicator of Diet. Science 1978, 201, 908–910. [Google Scholar] [CrossRef]
- Croitor, R. Giant Deer Megaloceros giganteus (Cervidae, Mammalia) from Late Pleistocene of Moldova. Oltenia. Stud. Si Comunicari. Stiintele Nat. 2008, 24, 262–266. [Google Scholar]
- Croitor, R.; Stefaniak, K.; Pawłowska, K.; Ridush, B.; Wojtal, P.; Stach, M. Giant Deer Megaloceros giganteus Blumenbach, 1799 (Cervidae, Mammalia) from Palaeolithic of Eastern Europe. Quat. Int. 2014, 326–327, 91–104. [Google Scholar] [CrossRef]
- Croitor, R. Taxonomy, Systematics and Evolution of Giant Deer Megaloceros giganteus (Blumenbach, 1799) (Cervidae, Mammalia) from the Pleistocene of Eurasia. Quaternary 2021, 4, 36. [Google Scholar] [CrossRef]
- Shpansky, A.V. The Giant Deer Megaloceros giganteus (Blum.) (Mammalia, Artiodactyla) from the Southeastern West Siberian Plain. Bjulleten’ Mosk. Obs. Ispyt. Prir. 2011, 86, 18–30. [Google Scholar]
- Shpansky, A.V. Skeleton of the Giant Deer Megaloceros giganteus giganteus (Blumenbach, 1803) (Mammalia, Artiodactyla) from the Irtysh Region near Pavlodar. Paleontol. J. 2014, 48, 534–550. [Google Scholar] [CrossRef]
- Vislobokova, I.A. Morphology, Taxonomy, and Phylogeny of Megacerines (Megacerini, Cervidae, Artiodactyla). Paleontol. J. 2013, 47, 833–950. [Google Scholar] [CrossRef]
- Scheglova, V.V. On Systematical Position and History of Development of Giant Deer. Rep. Acad. Sci. USSR 1950, 73, 813–816. [Google Scholar]
- Scheglova, V.V. On Giant Deer (Genus Megaloceros) from USSR. Proc. Belarusian State Univ. Geol. Ser. 1958, 43, 173–188. [Google Scholar]
- Paunović, G.; Bogićević, K.; Urošević, A. Intraspecific Differentiation and Sexual Dimorphism in Giant Deer (Megaloceros giganteus Blumenbach, 1799; Artiodactyla, Cervidae) Skulls Found in Serbia. Acta Zool. 2021, 102, 171–181. [Google Scholar] [CrossRef]
- Van der Made, J. The Evolution and Biogeography of the Pleistocene Giant Deer Megaloceros giganteus (Cervidae, Mammalia). In Late Neogene and Quarternary Biodiversity and Evolution: Regional Developments and Interregional Correlations; Kahlke, R.-D., Maul, L.C., Mazza, P.P.A., Eds.; Courier Forschungsinstitut Senckenberg: Frankfurt, Germany, 2006; pp. 117–129. [Google Scholar]
- Kahlke, R.-D. The History of the Origin, Evolution and Dispersal of the Late Pleistocene Mammuthus-Coelodonta Faunal Complex in Eurasia (Large Mammals); Mammoth Site of Hot Springs: Hot Springs, SD, USA, 1999. [Google Scholar]
- Kahlke, R.-D. The Origin of Eurasian Mammoth Faunas (Mammuthus-Coelodonta Faunal Complex). Quat. Sci. Rev. 2014, 96, 32–49. [Google Scholar] [CrossRef]
- Guthrie, R.D. Mammals of the Mammoth Steppe as Paleoecological Indicators. In Paleoecology of Beringia; Hopkins, D.M., Chambers, F.M., Matthews, J.V., Schweger, C.E., Young, S.B., Eds.; Academic Press: Cambridge, MA, USA, 1982; pp. 307–326. [Google Scholar]
- Rivals, F.; Ziegler, R. High-Resolution Paleoenvironmental Context for Human Occupations during the Middle Pleistocene in Europe (MIS 11, Germany). Quat. Sci. Rev. 2018, 188, 136–142. [Google Scholar] [CrossRef]
- Britton, K.; Jimenez, E.-L.; Le Corre, M.; Pederzani, S.; Daujeard, C.; Jaouen, K.; Vettese, D.; Tütken, T.; Hublin, J.-J.; Moncel, M.-H. Multi-Isotope Zooarchaeological Investigations at Abri Du Maras: The Paleoecological and Paleoenvironmental Context of Neanderthal Subsistence Strategies in the Rhône Valley during MIS 3. J. Hum. Evol. 2023, 174, 103292. [Google Scholar] [CrossRef]
- Van Geel, B.; Sevink, J.; Mol, D.; Langeveld, B.W.; van der Ham, R.W.J.M.; van der Kraan, C.J.M.; van der Plicht, J.; Haile, J.S.; Rey-Iglesia, A.; Lorenzen, E.D. Giant Deer (Megaloceros Giganteus) Diet from Mid-Weichselian Deposits under the Present North Sea Inferred from Molar-Embedded Botanical Remains. J. Quat. Sci. 2018, 33, 924–933. [Google Scholar] [CrossRef]
- Chritz, K.L.; Dyke, G.J.; Zazzo, A.; Lister, A.M.; Monaghan, N.T.; Sigwart, J.D. Palaeobiology of an Extinct Ice Age Mammal: Stable Isotope and Cementum Analysis of Giant Deer Teeth. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 282, 133–144. [Google Scholar] [CrossRef]
- Delpech, F. Les Faunes du Paléolithique Supérieur Dans le Sud-Ouest de la France; Cahiers du Quaternaire; Editions du CNRS: Paris, France, 1983; ISBN 978-2-222-03258-8. [Google Scholar]
- Gastearena, K.M. Hallazgos de macromamíferos poco frecuentes en yacimientos arqueológicos y paleontológicos del Pleistoceno de la región cantábrica. Kobie. Paleoantropol. 2011, 30, 83–110. [Google Scholar]
- Bradshaw, R.H.W.; Hannon, G.E.; Lister, A.M. A Long-Term Perspective on Ungulate–Vegetation Interactions. For. Ecol. Manag. 2003, 181, 267–280. [Google Scholar] [CrossRef]
- Álvarez-Lao, D.J.; García, N. Southern Dispersal and Palaeoecological Implications of Woolly Rhinoceros (Coelodonta Antiquitatis): Review of the Iberian Occurrences. Quat. Sci. Rev. 2011, 30, 2002–2017. [Google Scholar] [CrossRef]
- Daujeard, C.; Vettese, D.; Britton, K.; Béarez, P.; Boulbes, N.; Crégut-Bonnoure, E.; Desclaux, E.; Lateur, N.; Pike-Tay, A.; Rivals, F.; et al. Neanderthal Selective Hunting of Reindeer? The Case Study of Abri Du Maras (South-Eastern France). Archaeol Anthr. Sci. 2019, 11, 985–1011. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Location | Date | Analysis | References |
|---|---|---|---|---|
| Chinchon I l. 13 | France | MIS 2 | Biogeography | [7] |
| Le Pignon | France | MIS 2 | Biogeography | [31] |
| L’Arquet | France | MIS 3 | Biogeography | [32] |
| La Calmette | France | MIS 4-3 | Biogeography | [33] |
| Cornille | France | MIS 2 | Biogeography | [34] |
| Barasses l. 2-3 | France | MIS 3 | Biogeography | [35] |
| Observatoire l.4 | France | MIS 4 | Biogeography | [36] |
| Baume Goulon | France | MIS 2 | Biogeography | [37] |
| Baume des Peyrards l. c-d | France | MIS 4 | Biogeography | [35] |
| Cavillon Foyer II | France | MIS 4 | Biogeography | [38] |
| Enfants coupe 7 | France | MIS 3 | Biogeography | [38] |
| Prince foyer B | France | MIS 4 | Biogeography | [38] |
| Tournal ens. II | France | MIS 3 | Biogeography, LM | [8] |
| Tournal ens. IV | France | MIS 2 | Biogeography, LM | [8] |
| Roc Traücat | France | MIS 4-2 | Biogeography | [39] |
| Pair non pair | France | MIS 3 | Biogeography, LM | [5,39] |
| Camiac | France | MIS 3 | Biogeography, LM | [4,40] |
| La Crouzade C.7 | France | MIS 3 | Biogeography | [41,42] |
| La Crouzade C.8 | France | MIS 3 | Biogeography | [41,42] |
| Belvis l. 7 | France | MIS 3 | Biogeography | [43] |
| Salpêtrière l. 3 | France | MIS 2 | Biogeography | [44] |
| Salpêtre de Pompignan | France | MIS 4-3 | Biogeography | [33] |
| L’Hortus | France | MIS 3 | Biogeography | [33] |
| Baumasse d’Antonègue | France | MIS 3 | Biogeography | [33] |
| Gazel l. 7-8 | France | MIS 2 | Biogeography | [43] |
| Teixoneres | Spain | MIS 3 | Biogeography, LM, DW | This work |
| Arbreda l. H | Spain | MIS 3 | Biogeography | [45] |
| Canyars | Spain | MIS 4 | Biogeography | [46] |
| Cova del Gegant l. 1 | Spain | MIS 3 | Biogeography | [47] |
| Abric Romaní | Spain | MIS 3 | Biogeography | [48] |
| La Rexidora | Spain | MIS 3 | Biogeography, DW | [49,50] |
| El Castillo l. 20 | Spain | MIS 3 | Biogeography | [4,51] |
| El Castillo l. 14 | Spain | MIS 3 | Biogeography | [4,51] |
| Labeko Koba l. IX superior | Spain | MIS 3 | Biogeography | [4,52] |
| Jou Puerta | Spain | MIS 3 | Biogeography | [11,50] |
| Covacho de Arenillas l. II | Spain | MIS 3 | Biogeography | [53] |
| Lezika | Spain | Unknown | Biogeography | [54] |
| Lezetxiki l. IIIa | Spain | MIS 2 | Biogeography | [55] |
| Las Caldas l. VII | Spain | MIS 2 | Biogeography | [56] |
| Morin | Spain | MIS 3 | Biogeography | [57] |
| Urtiagako Leizea | Spain | Unknown | Biogeography | [57] |
| Moros de Gabasa | Spain | MIS 3 | Biogeography | [58] |
| Ermitons | Spain | MIS 3 | Biogeography | [59] |
| Italy | Italy | Unknown | LM | Raven, 1935 in [60] |
| Fouvent | France | MIS 3 | LM | [61] |
| Kent’s Cavern | England | MIS 3 | DW | [62] |
| Netherlands | Netherlands | MIS 3 | DW | [63] |
| Bruine Bank | North Sea | MIS 3 | DW | [63] |
| Siuren I | Crimea | MIS 3 | DW | [64] |
| Geißenklösterle | Germany | MIS 3 | DW | [65] |
| Einhornhöhle | Germany | MIS 3 | DW | [65] |
| Große Ofnet | Germany | MIS 3 | DW | [65] |
| Wolftalhöhle | Germany | MIS 3 | DW | [65] |
| Site | P4_L | P4_w | M1_L | M1_w | M2_L | M2_w | M3_L | M3_w | M1-M3_L | |
|---|---|---|---|---|---|---|---|---|---|---|
| Teixoneres_IIa | n | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| m | 17.4 | 22.3 | 31.77 | 24.7 | 30.9 | 27.51 | 27.54 | 77 | ||
| Tournal ens. II | n | 3 | 3 | 2 | 2 | 3 | 3 | 3 | 3 | |
| m | 19.1 | 24.2 | 25 | 30.3 | 28.2 | 31.7 | 30.2 | 29.5 | ||
| min | 19 | 24 | 25 | 30 | 28 | 31.5 | 30 | 29.5 | ||
| max | 19.2 | 24.5 | 25 | 30.5 | 28.5 | 32 | 30.5 | 29.5 | ||
| Pair-non-Pair | n | 12 | 12 | 2 | 2 | 5 | 5 | 9 | 9 | |
| m | 18.7 | 26.1 | 26.3 | 34.5 | 28.6 | 33.9 | 29.8 | 30.5 | ||
| s | 0.6 | 2 | 0.7 | 0.9 | 1.8 | 1.4 | ||||
| min | 18 | 22 | 26 | 34.5 | 28 | 33 | 27.5 | 28 | ||
| max | 19.5 | 27 | 26.5 | 34.5 | 29 | 35 | 32.5 | 32.5 | ||
| Camiac | n | 1 | 2 | 2 | 1 | |||||
| m | 19.5 | 25.8 | 27.7 | 32.8 | ||||||
| min | 25 | 27.3 | ||||||||
| max | 26.5 | 27.7 | ||||||||
| Italy | n | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| m | 19 | 24 | 28.5 | 28 | 31 | 30 | 29 | 27 | 84.5 | |
| Fouvent | n | 1 | 1 | 4 | 4 | 2 | 2 | |||
| m | 28 | 33 | 28 | 29.4 | 34 | 27.3 | ||||
| min | 26 | 27 | 33 | 25 | ||||||
| max | 31 | 32 | 35 | 29.5 | ||||||
| Teixoneres_IIa and b | n | 2 | 2 | 3 | 3 | 2 | 2 | 3 | 2 | |
| Cervus elaphus | m | 14.61 | 18.43 | 18.99 | 21.43 | 22.22 | 24.08 | 22.32 | 24.81 | |
| s | 0.65 | 1.32 | 2.32 | 4.04 | 2.32 | 1.78 | 1.05 | 1.04 | ||
| min | 14.15 | 17.5 | 16.46 | 16.8 | 20.58 | 22.82 | 21.5 | 24.07 | ||
| max | 15.07 | 19.36 | 21 | 24.25 | 23.86 | 25.34 | 23.5 | 25.54 |
| Site | Mp | Sh | Sk | Ca | B | Eh | Ec | Ce | Dd | Cc | Rt | Ss | C | Rr | St | Mg |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chinchon I l. 13 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 |
| Le Pignon | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| L’Arquet | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 |
| La Calmette | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| Cornille | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 |
| Barasses l. 2-3 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 |
| Observatoire l.4 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 |
| Baume Goulon | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Baume des Peyrards l. c-d | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 |
| Cavillon Foyer II | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Enfants coupe 7 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 |
| Prince foyer B | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 |
| Tournal ens. II | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 |
| Tournal ens. IV | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 |
| Roc Traücat | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Pair non pair | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 |
| Camiac | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| La Crouzade C.7 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| La Crouzade C.8 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 |
| Belvis l. 7 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 |
| Salpêtrière l. 3 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Salpêtre de Pompignan | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 |
| L’Hortus | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Baumasse d’Antonègue | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Gazel l. 7-8 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 |
| Teixoneres IIa | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Arbreda l. H | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Canyars | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Cova del Gegant l. 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Abric Romaní | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| Rexidora | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| El Castillo l. 20 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 |
| El Castillo l. 14 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 |
| Labeko Koba l. IX superior | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 |
| Jou Puerta | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 |
| Covacho de Arenillas l. II | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 |
| Lezika | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| Lezetxiki l. IIIa | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 |
| Las Caldas l. VII | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Morin | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 |
| Urtiagako Leizea | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 |
| Moros de Gabasa | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 |
| Ermitons | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 |
| Site | Microwear | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| n | NP | NS | %LP | %G | SWS | %HC | %XS | ||
| Teixoneres unit IIa | |||||||||
| Megaloceros giganteus | 1 | 14.5 | 19 | 0 | 0 | 0 | 0 | 0 | |
| Bos primigenius | 1 | 14 | 16.5 | 0 | 0 | 0 | 0 | 100 | |
| Cervus elaphus | m | 4 | 9 | 25 | 0 | 0 | 0.25 | 0 | 100 |
| sd | 7.22 | 3.24 | 0.5 | ||||||
| Coelodonta antiquitatis | 1 | 3.5 | 20.5 | 0 | 0 | 0 | 0 | 100 | |
| Equus caballus | m | 5 | 12.7 | 28.4 | 0 | 0 | 0 | 0 | 100 |
| sd | 7.69 | 3.45 | 0 | ||||||
| Equus hydruntinus | 1 | 3 | 19 | 0 | 0 | 1 | 0 | 0 | |
| Rexidora | 1 | 24.5 | 20.5 | 0 | 0 | 1 | 0 | ||
| Kent’s Cavern | m | 18 | 18.7 | 16.8 | 55.6 | 27.8 | 1.1 | 33.3 | |
| sd | 0.6 | 0.8 | |||||||
| Siuren I | m | 4 | 8.6 | 12.8 | 73.3 | 0 | 1.1 | 0 | |
| sd | 2.01 | 0.75 | |||||||
| Netherlands | 12 | 13.9 | 13.4 | 75 | 37.5 | 1.4 | 43.8 | ||
| Bruine bank | 28 | 13.6 | 13 | 78.6 | 39.3 | 1.4 | 53.6 | ||
| Geißenklösterle | 1 | 28.5 | 9.5 | 2 | |||||
| Große Ofnet | 10 | 16.4 | 13.7 | 100 | 20 | 1 | 30 | ||
| Einhornhöhle | 3 | 21.4 | 11.7 | 100 | 0 | 1.2 | 0 | ||
| Wolftalhöhle | 1 | 11.5 | 8 | 2 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uzunidis, A.; Rivals, F.; Rufà, A.; Blasco, R.; Rosell, J. The Exceptional Presence of Megaloceros giganteus in North-Eastern Iberia and Its Palaeoecological Implications: The Case of Teixoneres Cave (Moià, Barcelona, Spain). Diversity 2023, 15, 299. https://doi.org/10.3390/d15020299
Uzunidis A, Rivals F, Rufà A, Blasco R, Rosell J. The Exceptional Presence of Megaloceros giganteus in North-Eastern Iberia and Its Palaeoecological Implications: The Case of Teixoneres Cave (Moià, Barcelona, Spain). Diversity. 2023; 15(2):299. https://doi.org/10.3390/d15020299
Chicago/Turabian StyleUzunidis, Antigone, Florent Rivals, Anna Rufà, Ruth Blasco, and Jordi Rosell. 2023. "The Exceptional Presence of Megaloceros giganteus in North-Eastern Iberia and Its Palaeoecological Implications: The Case of Teixoneres Cave (Moià, Barcelona, Spain)" Diversity 15, no. 2: 299. https://doi.org/10.3390/d15020299