
Impact of Global Warming on Kryal Fauna: Thermal Tolerance Response of Diamesa steinboecki (Goetghebuer, 1933; Chironomidae)

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Larvae and Acclimatization
2.2. Survival Following Heat Shock
2.3. RNA Extraction and Complementary DNA (cDNA) Synthesis
2.4. cDNA Amplification by Real-Time PCR (RT-PCR)
2.5. Statistical Analysis and Figure Design
3. Results
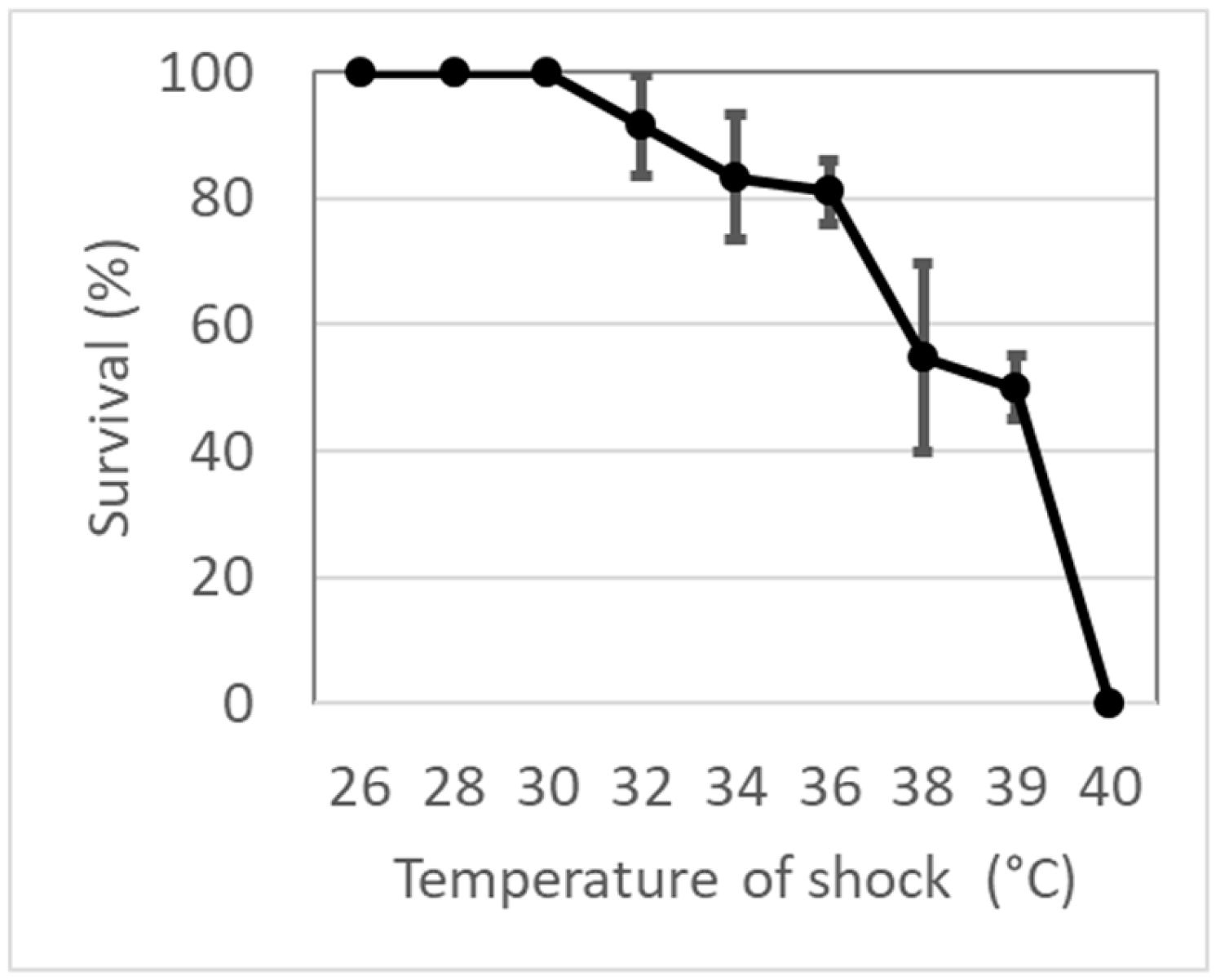
3.1. Survival and Lethal Temperature Determination
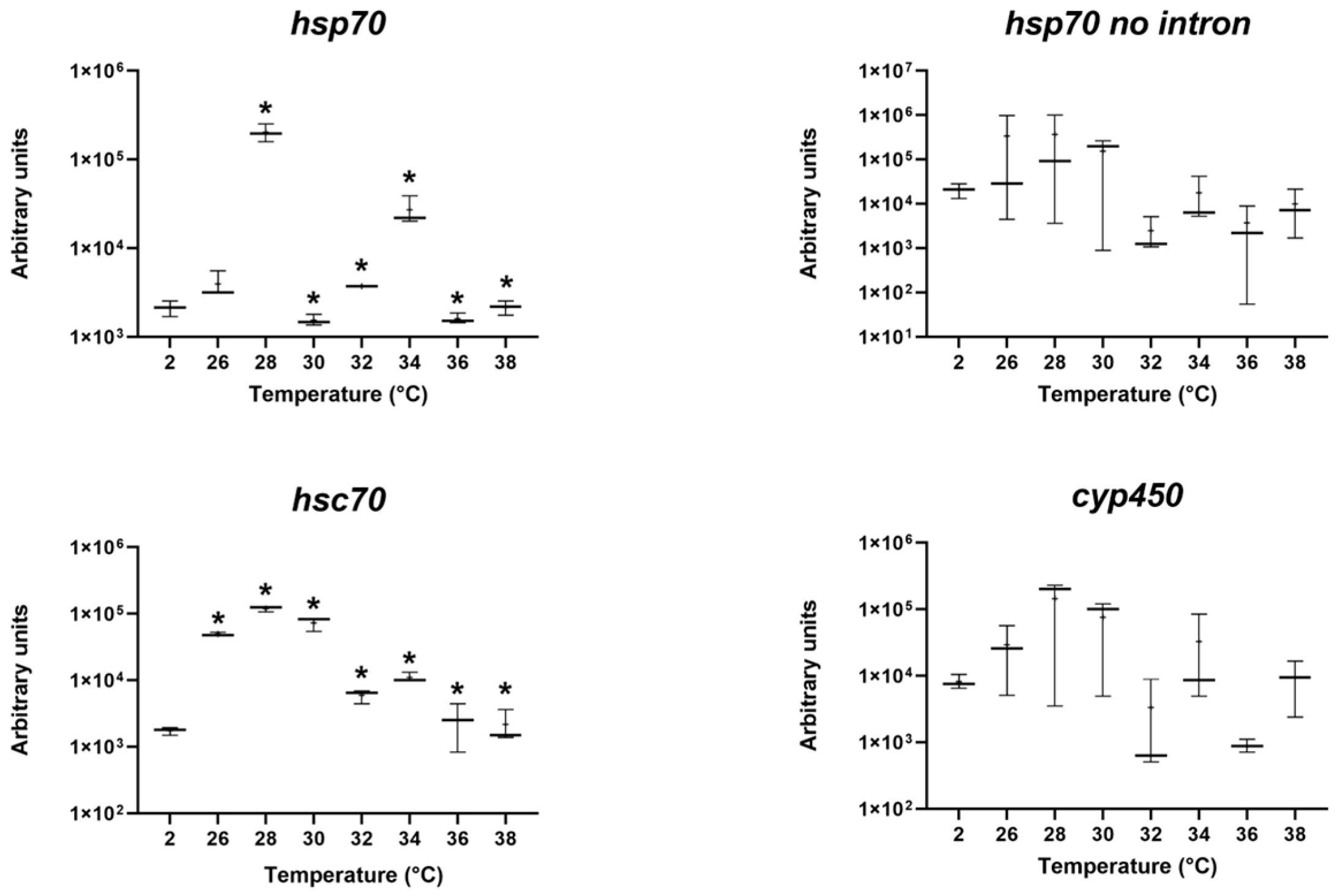
3.2. Gene Expression after Thermal Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giersch, J.J.; Jordan, S.; Luikart, G.; Jones, L.A.; Hauer, F.R.; Muhlfeld, C.C. Climate-induced range contraction of a rare alpine aquatic invertebrate. Freshw. Sci. 2015, 34, 53–65. [Google Scholar] [CrossRef]
- Jacobsen, D.; Dangles, O. Environmental harshness and global richness patterns in glacier-fed streams. Glob. Ecol. Biogeogr. 2012, 21, 647–656. [Google Scholar] [CrossRef]
- Birrell, J.H.; Shah, A.A.; Hotaling, S.; Giersch, J.J.; Williamson, C.E.; Jacobsen, D.; Woods, H.A. Insects in high-elevation streams: Life in extreme environments imperiled by climate change. Glob. Change Biol. 2020, 26, 6667–6684. [Google Scholar] [CrossRef] [PubMed]
- Lencioni, V.; Jousson, O.; Guella, G.; Bernabò, P. Cold adaptive potential of chironomids overwintering in a glacial stream. Physiol. Entomol. 2015, 40, 43–53. [Google Scholar] [CrossRef]
- Danks, H.V. How aquatic insects live in cold climates. BioOne 2007, 139, 443–471. [Google Scholar] [CrossRef]
- Lencioni, V. Survival strategies of freshwater insects in cold environments. J. Limnol. 2004, 63, 45–55. [Google Scholar] [CrossRef]
- Schütz, S.A.; Füreder, L. Unexpected patterns of chironomid larval size in an extreme environment: A highly glaciated, alpine stream. Hydrobiologia 2018, 820, 49–63. [Google Scholar] [CrossRef]
- Lencioni, V.; Bernabò, P.; Vanin, S.; Di Muro, P.; Beltramini, M. Respiration rate and oxy-regulatory capacity in cold stenothermal chironomids. J. Insect Physiol. 2008, 54, 1337–1342. [Google Scholar] [CrossRef]
- Lencioni, V.; Bernabò, P. Thermal survival limits of young and mature larvae of a cold stenothermal chironomid from the Alps (Diamesinae: Pseudodiamesa branickii [Nowicki, 1873]). Insect Sci. 2017, 24, 314–324. [Google Scholar] [CrossRef]
- Lencioni, V. Glacial influence and stream macroinvertebrate biodiversity under climate change: Lessons from the Southern Alps. Sci. Total. Environ. 2018, 622–623, 563–575. [Google Scholar] [CrossRef]
- Makarchenko, E.A.; Semenchenko, A.A.; Palatov, D.M. Taxonomy of Diamesa steinboecki group (Diptera: Chironomidae: Diamesinae), with description and DNA barcoding of new species. I. Subgroups steinboecki and longipes. Zootaxa 2022, 5125, 483–512. [Google Scholar] [CrossRef] [PubMed]
- Lencioni, V.; Stella, E.; Grazia, M.; Bellin, A. On the delay between water temperature and invertebrate community response to warming climate. Sci. Total. Environ. 2022, 837, 155759. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2022: Impacts, Adaptation and Vulnerability ; Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; p. 3056. [Google Scholar]
- Niedrist, G.H.; Füreder, L. Real-time warming of Alpine streams: (Re)defining invertebrates temperature preferences. River Res. 2021, 37, 283–293. [Google Scholar] [CrossRef]
- Webb, B.W.; Hannah, D.M.; Moore, R.D.; Brown, L.E.; Nobilis, F. Recent advances in stream and river temperature research. Hydrol. Process. 2008, 22, 902–918. [Google Scholar] [CrossRef]
- Brown, L.E.; Khamis, K.; Wilkes, M.; Blaen, P.; Brittain, J.E.; Carrivick, J.L.; Fell, S.; Friberg, N.; Füreder, L.; Gislason, G.M. Functional diversity and community assembly of river invertebrates show globally consistent responses to decreasing glacier cover. Nat. Ecol. Evol. 2018, 2, 325–333. [Google Scholar] [CrossRef]
- Milner, A.M.; Brittain, J.E.; Castella, E.; Petts, G.E. Trends of macroinvertebrate community structure in glacial fed streams in relation to environmental conditions: A synthesis. Freshw. Biol. 2001, 46, 1833–1848. [Google Scholar] [CrossRef]
- Caissie, D. The thermal regime of rivers: A review. Freshw. Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Dickson, N.E.; Carrivick, J.L.; Brown, L.E. Flow regulation alters alpine river thermal regimes. J. Hydrol. 2012, 464–465, 505–516. [Google Scholar] [CrossRef]
- Wilkes, M.A.; Carrivick, J.L.; Castella, E.; Ilg, C.; Cauvy-Fraunié, S.; Fell, S.; Füreder, L.; Huss, M.; James, W.; Lencioni, V.; et al. Glacier retreat reorganises river habitats leaving refugia for Alpine invertebrate biodiversity poorly protected. Nat. Ecol. Evol. 2023. [Google Scholar] [CrossRef]
- Bernabò, P.; Rebecchi, L.; Jousson, O.; Martinez-Guitarte, J.L.; Lencioni, V. Thermotolerance and hsp70 heat shock response in the cold-stenothermal chironomid Pseudodiamesa branickii (NE Italy). Cell Stress Chaperones 2011, 16, 403–410. [Google Scholar] [CrossRef]
- Bernabò, P.; Viero, G.; Lencioni, V. A long noncoding RNA acts as a post-transcriptional regulator of heat shock protein (HSP70) synthesis in the cold hardy Diamesa tonsa under heat shock. PLoS ONE 2020, 15, e0227172. [Google Scholar] [CrossRef] [PubMed]
- Lencioni, V.; Bernabo, P.; Cesari, M.; Rebecchi, L. Thermal stress induces hsp70 proteins synthesis in larvae of the cold stream non-biting midge Diamesa cinerella Meigen. Arch. Insect Biochem. Physiol. 2013, 83, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Morrow, G.; Tanguay, R.M. Small heat shock protein expression and functions during development. Int. J. Biochem. Cell Biol. 2012, 44, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Kline, M.P.; Bimston, D.N.; Cotto, J.J. The heat-shock response: Regulation and function of heat-shock proteins and molecular chaperones. Essays Biochem. 1997, 32, 17–29. [Google Scholar]
- Muñiz-González, A.B.; Novo, M.; Martínez-Guitarte, J.L. Persistent pesticides: Effects of endosulfan at the molecular level on the aquatic invertebrate Chironomus riparius. Environ. Sci. Pollut. Res. 2021, 28, 31431–31446. [Google Scholar] [CrossRef]
- Nie, H.; Liu, L.; Wang, H.; Huo, Z.; Yan, X. Stress levels over time in Ruditapes philippinarum: The effects of hypoxia and cold stress on Hsp70 gene expression. Aquacult. Rep. 2018, 12, 1–4. [Google Scholar] [CrossRef]
- Xu, X.H.; Meng, X.; Gan, H.T.; Liu, T.H.; Yao, H.Y.; Zhu, X.Y.; Xu, J.T. Immune response, MT and HSP70 gene expression, and bioaccu-mulation induced by lead exposure of the marine crab, Charybdis japonica. Aquat. Toxicol. 2019, 210, 98–105. [Google Scholar] [CrossRef]
- Bernabò, P.; Gaglio, M.; Bellamoli, F.; Viero, G.; Lencioni, V. DNA damage and translational response during detoxification from copper exposure in a wild population of Chironomus riparius. Chemosphere 2017, 173, 235–244. [Google Scholar] [CrossRef]
- Lencioni, V.; Boschini, D.; Rebecchi, L. Expression of the 70 kDa Heat shock protein family in Alpine freshwater chironomids (Diptera, Chironomidae) under natural conditions. J. Limnol. 2009, 68, 251–256. [Google Scholar] [CrossRef]
- Benoit, J.B.; Lopez-Martinez, G.; Elnitsky, M.A.; Lee, R.E., Jr.; Denlinger, D.L. Dehydration-induced cross tolerance of Belgica antarctica larvae to cold and heat is facilitated by trehalose accumulation. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 152, 518–523. [Google Scholar] [CrossRef]
- Clark, M.; Fraser, K.; Burns, G.; Peck, L. The HSP70 heat shock response in the Antarctic fish Harpagifer antarcticus. Polar Biol. 2008, 31, 171–180. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Hayward, S.A.; Elnitsky, M.A.; Sandro, L.H.; Lee, R.E.; Denlinger, D.L. Continuous up-regulation of heat shock proteins in larvae, but not adults, of a polar insect. Proc. Natl. Acad. Sci. USA 2006, 103, 14223–14227. [Google Scholar] [CrossRef]
- Di Nica, V.; González, A.B.M.; Lencioni, V.; Villa, S. Behavioural and biochemical alterations by chlorpyrifos in aquatic insects: An emerging environmental concern for pristine Alpine habitats. Environ. Sci. Pollut. Res. 2020, 27, 30918–30926. [Google Scholar] [CrossRef] [PubMed]
- Muñiz-González, A.B.; Paoli, F.; Martínez-Guitarte, J.L.; Lencioni, V. Molecular biomarkers as tool for early warning by chlorpyrifos exposure on Alpine chironomids. Environ. Pollut. 2021, 290, 118061. [Google Scholar] [CrossRef] [PubMed]
- Rossaro, B.; Lencioni, V. A key to larvae of species belonging to the genus Diamesa from Alps and Apennines (Italy). Eur. J. Environ. Sci. 2015, 5, 62–79. [Google Scholar] [CrossRef]
- Holdway, D. Hydra Population Reproduction Toxicity Test Method. In Small-Scale Freshwater Toxicity Investigations, 1st ed.; Springer: Dordrecht, The Netherlands, 2005; pp. 395–411. [Google Scholar]
- Lencioni, V.; Bellamoli, F.; Bernabó, P.; Miari, F.; Scotti, A. Response of Diamesa spp. (Diptera: Chironomidae) from alpine streams to newly emergent contaminants and pesticides. J. Limnol. 2018, 77, 131–140. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 16–21. [Google Scholar] [CrossRef]
- Bouchard, R.W.; Carrillo, M.A.; Keils, S.A.; Ferrington, L.C. Freeze tolerance in larvae of the winter-active Diamesa mendotae Muttkowski (Diptera: Chironomidae): A contrast to adult strategy for survival at low temperatures. Hydrobiologia 2006, 568, 403–416. [Google Scholar] [CrossRef]
- Kim, B.M.; Rhee, J.S.; Lee, K.W.; Kim, M.J.; Shin, K.H.; Lee, S.J.; Lee, Y.M.; Lee, J.S. UV-B radiation-induced oxidative stress and p38 signaling pathway involvement in the benthic copepod Tigriopus japonicus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2015, 167, 15–23. [Google Scholar] [CrossRef]
- Muñiz-González, A.B. Ibuprofen as an emerging pollutant on non-target aquatic invertebrates: Effects on Chironomus riparius. Environ. Toxicol. Pharmacol. 2021, 81, 103537. [Google Scholar] [CrossRef] [PubMed]
- Tartarotti, B.; Torres, J.J. Sublethal stress: Impact of solar UV radiation on protein synthesis in the copepod Acartia tonsa. J. Exp. Mar. Biol. Ecol. 2009, 375, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, R.D. Stress Protein Expression Kinetics. Annu. Rev. Biomed. Eng. 2006, 8, 403–424. [Google Scholar]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell. 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Diller, K.R.; Aggarwal, S.J. Kinetics study of endogenous heat shock protein 70 expression. J. Biomech. Eng. 2003, 125, 794–797. [Google Scholar] [CrossRef] [PubMed]
- Richard, I.M.; Sandy, D. Westerheide, Chapter 268—The Heat Shock Response and the Stress of Misfolded Proteins. In Handbook of Cell Signaling, 2nd ed.; Ralph, A.B., Edward, A.D., Eds.; Academic Press: Cambridge, MA, USA, 2010; pp. 2231–2239. ISBN 9780123741455. [Google Scholar]
- Kim, J.; Joo, H.J.; Kim, Y.H.; Ahn, S.; Chang, J.; Hwang, B.; Lee, H.; Lee, J. Systemic Analysis of Heat Shock Response Induced by Heat Shock and a Proteasome Inhibitor MG132. PLoS ONE 2011, 6, e20252. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chang, Y.W.; Bai, J. High temperature stress induces expression of CYP450 genes and contributes to insecticide tolerance in Liriomyza trifolii. Pestic. Biochem. Physiol. 2021, 174, 104826. [Google Scholar] [CrossRef]
- Ruan, H.Y.; Meng, J.Y.; Yang, C.L.; Zhou, L.; Zhang, C.Y. Identification of Six Small Heat Shock Protein Genes in Ostrinia furnacalis (Lepidoptera: Pyralidae) and Analysis of Their Expression Patterns in Response to Environmental Stressors. J. Insect Sci. 2022, 22, 7. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñiz-González, A.-B.; Martínez-Guitarte, J.-L.; Lencioni, V. Impact of Global Warming on Kryal Fauna: Thermal Tolerance Response of Diamesa steinboecki (Goetghebuer, 1933; Chironomidae). Diversity 2023, 15, 708. https://doi.org/10.3390/d15060708
Muñiz-González A-B, Martínez-Guitarte J-L, Lencioni V. Impact of Global Warming on Kryal Fauna: Thermal Tolerance Response of Diamesa steinboecki (Goetghebuer, 1933; Chironomidae). Diversity. 2023; 15(6):708. https://doi.org/10.3390/d15060708
Chicago/Turabian StyleMuñiz-González, Ana-Belén, José-Luis Martínez-Guitarte, and Valeria Lencioni. 2023. "Impact of Global Warming on Kryal Fauna: Thermal Tolerance Response of Diamesa steinboecki (Goetghebuer, 1933; Chironomidae)" Diversity 15, no. 6: 708. https://doi.org/10.3390/d15060708