
Diversity and Distribution of Helminths in Wild Ruminants of the Russian Arctic: Reindeer (Rangifer tarandus), Muskoxen (Ovibos moschatus), and Snow Sheep (Ovis nivicola)

 ,
,  , , , , , , , , , , , and
, , , , , , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Helminth Recovery
2.2. DNA Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Taylor, J.J.; Lawler, J.P.; Aronsson, M.; Barry, T.; Bjorkman, A.D.; Christensen, T.; Coulson, S.J.; Cuyler, C.; Ehrich, D.; Falk, K.; et al. Arctic terrestrial biodiversity status and trends: A synopsis of science supporting the CBMP State of Arctic Terrestrial Biodiversity Report. Ambio 2020, 49, 833–847. [Google Scholar] [CrossRef]
- Convey, P. Antarctic terrestrial biodiversity in a changing world. Polar Biol. 2011, 34, 1629–1641. [Google Scholar] [CrossRef]
- Carlson, C.J.; Burgio, K.R.; Dougherty, E.R.; Phillips, A.J.; Bueno, V.M.; Clements, C.F.; Castaldo, G.; Dallas, T.A.; Cizauskas, C.A.; Cumming, G.S.; et al. Parasite biodiversity faces extinction and redistribution in a changing climate. Sci. Adv. 2017, 3, e1602422. [Google Scholar] [CrossRef]
- Zykin, V.S.; Zykina, V.S.; Orlova, L.A.; Stratigrafiya, I. Osnovnye zakonomernosti izmeneniya prirodnoy sredy I klimata v pleistotsene I golotsene Zapadnoy Sibiri (Stratigraphy and the main regularities of changes in the natural environment and climate in the Pleistocene and Holocene of Western Siberia). Arkheologiya 2000, 1, 3–22. (In Russian) [Google Scholar]
- European Glacial Landscapes. Maximum Extent of Glaciations; Palacois, D.; Hughes, P.D.; Garcia-Ruiz, J.M.; Andres, N. (Eds.) Elsevier: Amsterdam, The Netherlands, 2021; p. 546. ISBN 978-0-12-823498-3. [Google Scholar]
- Holand, Ø.; Mizin, I.; Weladji, R.B. Reindeer Rangifer tarandus (Linnaeus, 1758). In Handbook of the Mammals of Europe; Hackländer, K., Zachos, F.E., Eds.; Springer: Cham, Switzerland, 2022; pp. 247–277. ISBN 978-3-030-24474-3. [Google Scholar] [CrossRef]
- Mizin, I.A.; Sipko, T.P.; Davydov, A.V.; Gruzdev, A.R. The wild reindeer (Rangifer tarandus: Cervidae, Mammalia) on the Arctic Islands of Russia: A Review. Nat. Conserv. Res. 2018, 3, 1–14. [Google Scholar] [CrossRef]
- Danilov, P.I.; Panchenko, D.V.; Tirronen, K.F. Severnyy Olen’ Vostochnoy Fennoskandii (Reindeer of Eastern Fennoscandia); KarNTS RAN: Petrozavodsk, Russia, 2020; p. 187. ISBN 978-5-9274-0879-5. (In Russian) [Google Scholar]
- Korolev, A.N.; Mamontov, V.N.; Kholodova, M.V.; Baranova, A.I.; Shadrin, D.M.; Poroshin, E.A.; Efimov, V.A.; Kochanov, S.K. Polymorphism of the mtDNA control region in Reindeer (Rangifer tarandus) from the mainland of the Northeastern part of European Russia. Biol. Bull. 2017, 44, 882–893. [Google Scholar] [CrossRef]
- Safronov, V.M. Ekologiyai Ispol’zovaniye Dikogo Severnogo Olenya v Yakutii (Ecology and Use of Wild Reindeer in Yakutia); Izdatel’stvo SO RAN: Yakutsk, Russia, 2005; p. 188. ISBN 5-463-00087-5. (In Russian) [Google Scholar]
- Schmidt, N.M.; Stelvig, M. Muskox Ovibos moschatus (Zimmermann, 1780). In Handbook of the Mammals of Europe; Hackländer, K., Zachos, F.E., Eds.; Springer: Cham, Switzerland, 2022; pp. 313–325. ISBN 978-3-030-24474-3. [Google Scholar]
- Cuyler, C.; Rowell, J.; Adamczewski, J.; Anderson, M.; Blake, J.; Bretten, T.; Brodeur, V.; Campbel, M.; Checkley, S.L.; Cluff, H.D.; et al. Muskox status, recent variation, and uncertain future. Ambio 2020, 49, 805–819. [Google Scholar] [CrossRef]
- Revin, T.V.; Sopin, L.V.; Zheleznov, N.K. Snezhnyy Baran. Morfologiya, Sistematika, Ekologiya, Okhrana (Snow Sheep. Morphology, Systematics, Ecology, Protection); Nauka: Novosibirsk, Russia, 1988; p. 193. ISBN 5-02-029417-9. (In Russian) [Google Scholar]
- Baskin, L.; Danell, K. Ecology of Ungulates: A Handbook of Species in Eastern Europe and Northern and Central Asia; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2003; pp. 378–385. ISBN 3-540-43804-1. [Google Scholar]
- Sipko, T.P.; Gruzdev, A.R.; Egorov, S.S.; Tikhonov, V.G. Analis protsessa introduktsii ovtsebyka na severe Evrazii (The Analysis of Muskox Introduction in Northern Asia). Zool. Zhurnal 2007, 86, 620–627. (In Russian) [Google Scholar]
- Gordeeva, N.V.; Sipko, T.P.; Gruzdev, A.R. Izmenchivost’ mikrosatellitnoy DNK v populyatsiyakh ovtsebykov (Ovibos moschatus), akklimatizirovannykh v Rossii (Microsatellite DNA Variability in the Populations of Muskoxen (Ovibos moschatus) Transplanted into the Russian North). Genetika 2009, 45, 932–940. (In Russian) [Google Scholar]
- Argunov, A.V. Alien Species of the Mammalian Fauna in Yakutia. Russ. J. Biol. Invasions. 2018, 9, 313–326. [Google Scholar] [CrossRef]
- Poulin, R.; Presswell, B.; Bennett, J.; de Angeli Dutra, D.; Salloum, P.M. Biases in parasite biodiversity research: Why some helminth species attract more research than others. Int. J. Parasitol. Parasites Wildl. 2023, 21, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Mizkewitsch, V.Y. Gel’minty Severnogo Olenya i Vyzyvayemyye Imi Zabolevaniya (Reindeer Helminths and the Diseases They Cause); Kolos: Leningrad, Russia, 1967; 308p. (In Russian) [Google Scholar]
- Emelyanova, A.; Savolainen, A.; Oksanen, A.; Nieminen, P.; Loginova, O.; Abass, K.; Rautio, A. Research on Selected Wildlife Infections in the Circumpolar Arctic—A Bibliometric Review. Int. J. Environ. Res. Public Health 2022, 19, 11260. [Google Scholar] [CrossRef]
- Latypov, D.G.; Timerbayeva, R.R.; Kirillov, E.G. Parazitologiya i Invazionnyye Bolezni Zhvachnykh Zhivotnykh (Parasitology and Invasive Diseases of Ruminants); Lan: St. Petersburg, Russia, 2019; pp. 59–67. ISBN 978-5-8114-3561-6. (In Russian) [Google Scholar]
- Verocai, G.G.; Chaudhry, U.N.; Lejeune, M. Diagnostic Methods for Detecting Internal Parasites of Livestock. Vet. Clin. Food. Anim. 2020, 36, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Holterman, M.; van der Wurff, A.; van den Elsen, S.; van Megen, H.; Bongers, T.; Holovachov, O.; Bakker, J.; Helder, J. Phylum-Wide Analysis of SSU rDNA Reveals Deep Phylogenetic Relationships among Nematodes and Accelerated Evolution toward Crown Clades. Mol. Biol. Evol. 2006, 23, 1792–1800. [Google Scholar] [CrossRef]
- Vrain, T.C.; Wakarchuk, D.A.; Levesque, A.C.; Hamilton, R.I. Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fundam. Appl. Nematol. 1992, 15, 563–573. [Google Scholar]
- Gasser, R.B.; Chilton, N.B.; Hoste, H.; Beveridge, I. Rapid sequencing of rDNA from single worms and eggs of parasitic helminths. Nucleic Acids Res. 1993, 21, 2525–2526. [Google Scholar] [CrossRef] [PubMed]
- Nadler, S.A.; Hudspeth, D.S.S. Phylogeny of the Ascaridoidea (Nematoda: Ascaridida) based on three genes and morphology: Hypotheses of structural and sequence evolution. J. Parasitol. 2000, 86, 380–393. [Google Scholar] [CrossRef]
- Thomas, W.K.; Vida, J.T.; Frisse, L.M.; Mundo, M.; Baldwin, J.G. DNA sequences from formalin-fixed nematodes: Integrating molecular and morphological approaches to taxonomy. J. Nematol. 1997, 29, 250–254. [Google Scholar]
- Kanzaki, N.; Futai, K. A PCR primer set for determination of phylogenetic relationships of Bursaphelenchus species within the xylophilus group. Nematology 2002, 4, 35–41. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2004, 7, 203–214. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef]
- Loginova, O.A.; Kolpashchikov, L.A.; Spiridonov, S.E. First report of Orthostrongylus sp. (Nematoda: Protostrongylidae) in wild reindeer (Rangifer tarandus) from the Taimyr, Russia: Nearctic parasites in a Palearctic host. Parasitol. Res. 2023, 122, 685–689. [Google Scholar] [CrossRef]
- Kutz, S.J.; Ducrocq, J.; Verocai, G.G.; Hoar, B.M.; Colwell, D.D.; Beckmen, K.B.; Polley, L.; Elkin, B.T.; Hoberg, E.P. Parasites in ungulates of Arctic North America and Greenland: A view of contemporary diversity, ecology, and impact in a world under change. Adv. Parasitol. 2012, 79, 99–252. [Google Scholar] [CrossRef] [PubMed]
- Kutz, S.J.; Laaksonen, S.; Asbakk, K.; Nilssen, A.C. Parasitic Infections and Diseases. In Reindeer and Caribou. Health and Disease; Tryland, M., Kutz., S.J., Eds.; CRC Press (Taylor and Francis Group): Boca Raton, FL, USA; London, UK; New York, NY, USA, 2019; pp. 177–235. ISBN 978-1-4822-5068-8. [Google Scholar]
- Eduardo, S.L. The taxonomy of the family Paramphistomatidae Fischoeder, 1901 with special references to the morphology of species occurring in ruminants. II. Revision of the genus Paramphistomum Fischoeder, 1901. Syst. Parasitol. 1982, 4, 189–238. [Google Scholar] [CrossRef]
- Skrjabin, K.I. Trematody Zhivotnykh i Cheloveka. Osnovy Trematodologii (Trematodes of Animals and Man. Principles of Trematodology); Izdatelstvo Akademii Nauk SSSR: Moscow, Russia, 1949; Volume 3, p. 600. (In Russian) [Google Scholar]
- Zhaltsanova, D.S. Occurrence of Fischoederius elongatus in the Buryat ASSR. Mater. Nauchnykh Issled. Chlenov Vsesoyuznogo Obs. Gel’mintologov 1972, 24, 49–50. (In Russian) [Google Scholar]
- Willmott, S.; Pester, F.R.N. Variations in Faecal Egg-counts in Paramphistome Infections as Determined by a New Technique. J. Helminthol. 1952, 26, 2–3+147–156. [Google Scholar] [CrossRef]
- Skrjabin, K.I. Trematody Zhivotnykh i Cheloveka. Osnovy Trematodologii (Trematodes of Animals and Man. Principles of Trematodology); Izdatelstvo Akademii Nauk SSSR: Moscow, Russia, 1952; Volume 7, p. 762. (In Russian) [Google Scholar]
- Haukisalmi, V.; Laaksonen, S.; Oksanen, A.; Beckmen, K.; Halajian, A.; Yanagida, T.; Nakao, M. Molecular taxonomy and subgeneric classification of tapeworms of the genus Moniezia Blanchard, 1891 (Cestoda, Anoplocephalidae) in northern cervids (Alces and Rangifer). Parasitol. Int. 2018, 67, 218–224. [Google Scholar] [CrossRef]
- Meradi, S.; Bentounsi, B.; Zouyed, I.; Cabaret, J. The steppe species of gastrointestinal nematodes of small ruminants, with a focus on Marshallagia: Climate as a key determinant. Parasite J. De La Société Française De Parasitol. 2011, 183, 261–269. [Google Scholar] [CrossRef]
- Skrjabin, K.I.; Shikhobalova, N.P.; Schultz, R.S. Trikhostrongilidy Zhivotnykh i cheloveka. Osnovy Nematodologii (Trichostrongylids of Animals and Man. Principles of Nematodology); Izdatelstvo Akademii Nauk SSSR: Moscow, Russian, 1954; Volume 3, p. 684. (In Russian) [Google Scholar]
- Loginova, O.A.; Chuprak, D.I.; Rozenfel’d, S.B. O samostoyatel’nom diagnosticheskom znachenii obolochki yaytsa Marshallagia spp. Nnematoda: Strongylida) (On independent diagnostic value of egg membranes of Marshallagia spp. (Nematoda: Strongylida)). In Proceedings of the Modern Issues of General and Applied Parasitology Conference, Voronezh, Russia, 27–28 October 2022; pp. 56–61. (In Russian). [Google Scholar] [CrossRef]
- Dallas, J.F.; Irvine, R.J.; Halvorsen, O. DNA evidence that Marshallagia marshalli Ransom, 1907 and M. occidentalis Ransom, 1907 (Nematoda: Ostertagiinae) from Svalbard reindeer are conspecific. Syst. Parasitol. 2001, 50, 101–103. [Google Scholar] [CrossRef]
- Kuchboev, A.; Sobirova, K.; Karimova, R.; Amirov, O.; von Samson-Himmelstjerna, G.; Krücken, J. Molecular analysis of polymorphic species of the genus Marshallagia (Nematoda: Ostertagiinae). Parasites Vectors 2020, 13, 1–12. [Google Scholar] [CrossRef]
- Fruetel, M.; Lankester, M.W. Gastrointestinal helminths of woodland and barren ground caribou (Rangifer tarandus) in Canada, with keys to species. Can. J. Zool. 1989, 67, 2253–2269. [Google Scholar] [CrossRef]
- Thomas, R.J. A comparative study of the infective larvae of Nematodirus species parasitic in sheep. Parasitology 1957, 47, 60–65. [Google Scholar] [CrossRef]
- Baskin, L.M. Odomashnivaniye Severnogo Olenya. Ot Okhotnika Do Pastukha i Ranchevoda (Reindeer Domestication. from Hunter to Herdsman and Rancher); Tovarishchestvo Nauchnykh Izdaniy KMK: Moscow, Russia, 2021; p. 280. ISBN 978-5-907372-89-4. (In Russian) [Google Scholar]
- Hrabok, J.T.; Oksanen, A.; Nieminen, M.; Waller, P.J. Population dynamics of nematode parasites of reindeer in the sub-arctic. Vet. Parasitol. 2006, 142, 301–311. [Google Scholar] [CrossRef]
- Jokelainen, P.; Moroni, B.; Hoberg, E.; Oksanen, A.; Laaksonen, S. Gastrointestinal parasites in reindeer (Rangifer tarandus tarandus): A review focusing on Fennoscandia. Vet. Parasitol. Reg. Stud. Rep. 2019, 17, 100317. [Google Scholar] [CrossRef]
- Turgeon, G.; Kutz, S.J.; Lejeune, M.; St-Laurent, M.H.; Pelletier, F. Parasite prevalence, infection intensity and richness in an endangered population, the Atlantic-Gaspésie caribou. Int. J. Parasitol. Parasites Wildl. 2018, 7, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Goetzmann, W.; Ibbotson, R.G.; Ross, S.A. Survivorship bias in performance studies. Rev. Financ. Stud. 1992, 5, 553–580. [Google Scholar] [CrossRef]
- Pyziel, A.M.; Laskowski, Z.; Demiaszkiewicz, A.W.; Höglund, J. Interrelationships of Dictyocaulus spp. in wild ruminants with morphological description of Dictyocaulus cervi n. sp.(Nematoda: Trichostrongyloidea) from red deer, Cervus elaphus. J. Parasitol. 2017, 103, 506–518. [Google Scholar] [CrossRef]
- Filip-Hutsch, K.; Demiaszkiewicz, A.W.; Chęcińska, A.; Hutsch, T.; Czopowicz, M.; Pyziel, A.M. First report of a newly-described lungworm, Dictyocaulus cervi (Nematoda: Trichostrongyloidea), in moose (Alces alces) in central Europe. Int. J. Parasitol. Parasites Wildl. 2020, 13, 275–282. [Google Scholar] [CrossRef]
- Verocai, G.G.; Kutz, S.J.; Simard, M.; Hoberg, E.P. Varestrongylus eleguneniensis sp. n. (Nematoda: Protostrongylidae): A widespread, multi-host lungworm of wild North American ungulates, with an emended diagnosis for the genus and explorations of biogeography. Parasites Vectors 2014, 7, 1–22. [Google Scholar] [CrossRef]
- Verocai, G.G.; Hoberg, E.P.; Simard, M.; Beckmen, K.B.; Musiani, M.; Wasser, S.; Cuyler, C.; Manseau, M.; Chaudhry, U.N.; Kashivakura, C.K.; et al. The biogeography of the caribou lungworm, Varestrongylus eleguneniensis (Nematoda: Protostrongylidae) across northern North America. Int. J. Parasitol. Parasites Wildl. 2020, 11, 93–102. [Google Scholar] [CrossRef]
- Boev, S.N. Protostrongilidy. Osnovy Nematodologii (Protostrongylids. Principles of Nematodology); Izdatelstvo Akademii Nauk SSSR: Moscow, Russian, 1975; Volume 25, p. 338. (In Russian) [Google Scholar]
- Verocai, G.G.; Kafle, P.; Sulliotti, V.; Lejeune, M.; Hoberg, E.P.; Kutz, S.J. Morphometry of first-stage larva of Orthostrongylus macrotis (Nematoda: Protostrongylidae), lungworm of wild ungulates from Western North America. J. Parasitol. 2022, 108, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Greiner, E.C. Laboratory Diagnostics of Parasitic Diseases. In Equine Infectious Diseases, 2nd ed.; Sellon, D.C., Long, M.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 449–456. ISBN 978-1455708918. [Google Scholar]
- Oksanen, A. Endectocide treatment of the reindeer. Rangifer 1999, 19, 7–50. [Google Scholar] [CrossRef]
- Loginova, O.A. D-shaped nematode eggs in the feces of Rangifer tarandus. RJN 2023. submitted. [Google Scholar]
- Rebecchi, L.; Boschetti, C.; Nelson, D.R. Extreme-tolerance mechanisms in meiofaunal organisms: A case study with tardigrades, rotifers and nematodes. Hydrobiologia 2020, 847, 2779–2799. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
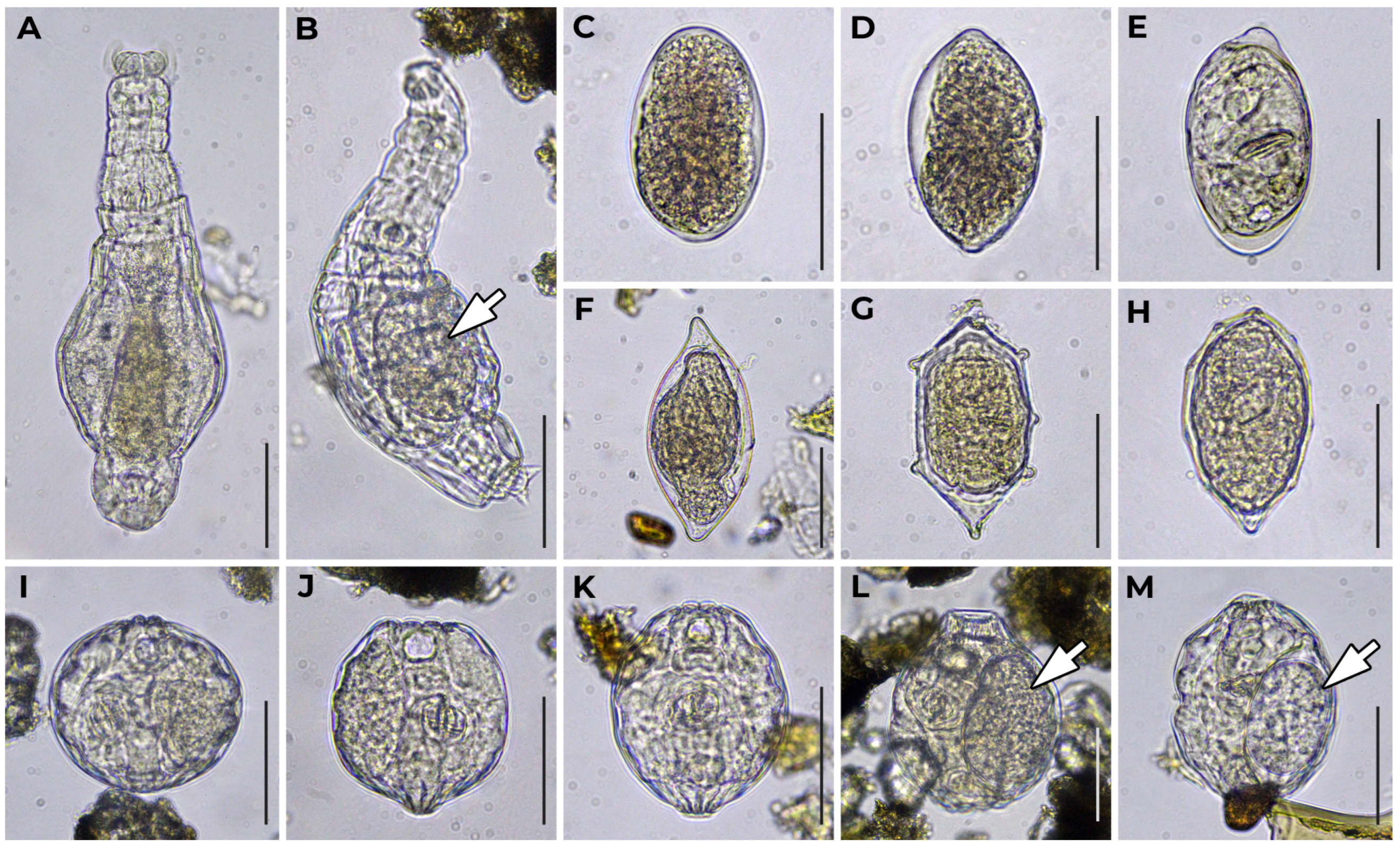{kind=link}
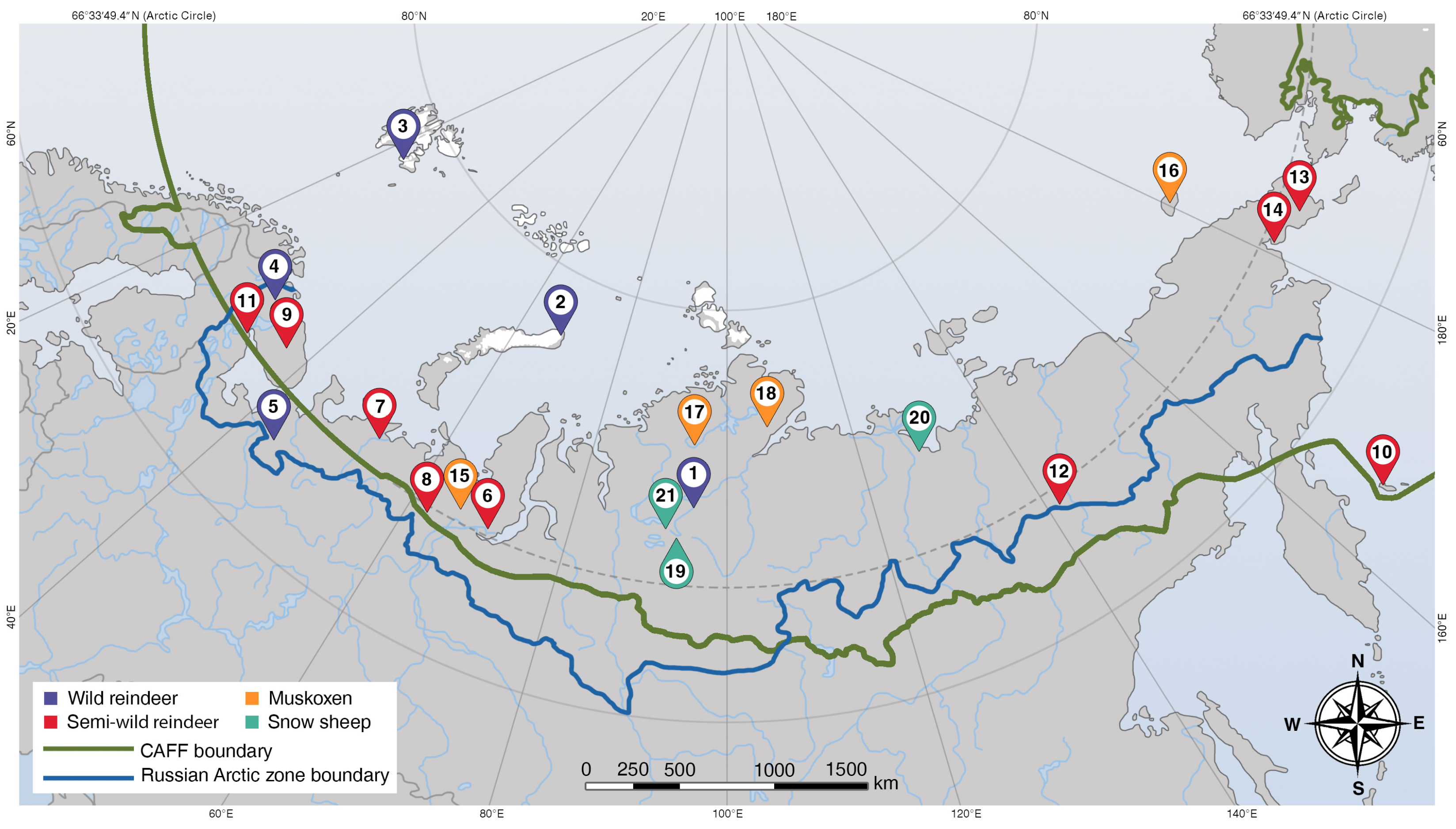
| Sample Set ID | Map Reference | Host Species | Number of Fecal Samples | Location | Coordinates (Decimal Degrees) | Date Collected |
|---|---|---|---|---|---|---|
| 1 | 1 | Wild reindeer (Rangifer tarandus) | 19 | Taymyr Peninsula (Krasnoyarsk Krai) | 71.10780, 98.04150 | June 2020 |
| 2 | 2 | 3 | Novaya Zemlya, Arkhangelsk Oblast | 76.90835, 68.44880 | August 2021 | |
| 3 | 2 | 2 | Novaya Zemlya, Arkhangelsk Oblast | 76.90835, 68.44880 | July 2022 | |
| 4 | 3 | 15 | Barentsbugr, Svalbard 1 (Norway) | 78.04729, 14.28717 | August 2022 | |
| 5 | 4 | 21 | Murmansk Oblast | 68.50699, 30.03720 | September 2022 | |
| 6 | 5 | 11 | Arkhangelsk Oblast | 63.86779, 44.43266 | September 2022 | |
| 7 | 6 | Semiwild reindeer (R. tarandus) | 50 | Yamalo-Nenets Autonomous Okrug | 66.62200, 65.71646 | August 2018 |
| 8 | 7 | 59 | Nenets Autonomous Okrug | 67.97568, 52.91955 | October 2018 | |
| 9 | 8 | 2 | Khanty-Mansi Autonomous Okrug | 65.63307, 61.94561 | October 2019 | |
| 10 | 9 | 30 | Murmansk Oblast | 68.00424, 35.01228 | December 2020 | |
| 11 | 9 | 31 | Murmansk Oblast | 68.00424, 35.01228 | March 2021 | |
| 12 | 8 | 20 | Khanty-Mansi Autonomous Okrug | 65.63307, 61.94561 | August 2021 | |
| 13 | 10 | 42 | Bering Island 2 Kamchatka Krai | 55.26152, 165.93711 | October 2021 | |
| 14 | 11 | 7 | Republic of Karelia | 66.30072, 33.08216 | June 2022 | |
| 15 | 12 | 10 | Republic of Sakha (Yakutia) | 65.34745, 140.08864 | June 2022 | |
| 16 | 13 | 30 | Chukotka Autonomous Okrug | 65.46168, −173.47478 | June 2022 | |
| 17 | 14 | 8 | Chukotka Autonomous Okrug | 65.90614, −178.84644 | October 2022 | |
| 18 | 15 | Muskoxen (Ovibos moschatus) | 35 | Yamalo-Nenets Autonomous Okrug | 67.58240, 66.25280 | March 2022 |
| 19 | 16 | 24 | Wrangel Island, Chukotka Autonomous Okrug | 71.34890, −178.66722 | March 2022 | |
| 20 | 17 | 6 | Taymyr Peninsula (Krasnoyarsk Krai) | 73.10336, 95.11816 | July 2022 | |
| 21 | 18 | 25 | Taymyr Peninsula (Krasnoyarsk Krai) | 74.17696, 107.46475 | August 2022 | |
| 22 | 15 | 30 | Yamalo-Nenets Autonomous Okrug | 67.58240, 66.25280 | October 2022 | |
| 23 | 19 | Snow sheep (Ovis nivicola) | 21 | Taymyr Peninsula (Krasnoyarsk Krai) | 68.64397, 95.66562 | July 2016 |
| 24 | 20 | 13 | Republic of Sakha (Yakutia) | 70.51859, 129.50336 | June 2022 | |
| 25 | 21 | 5 | Taymyr Peninsula (Krasnoyarsk Krai) | 69.33130, 93.58480 | July 2022 |
| Sample Set ID | Species | GenBank 1 | Vouchers 2 |
|---|---|---|---|
| 1 | Orthostrongylus sp. 3 | OL700043 (for LSU) | IPEE_Parasites 14284 |
| OL700044 (for ITS2) | IPEE_Parasites 14284 | ||
| 1 | Varestrongylus eleguneniensis3 | OM743794 (for ITS2) | IPEE_Parasites 14298 |
| OL743795 (for ITS2) | IPEE_Parasites 14299 | ||
| 10 | Elaphostrongylus rangiferi 4 | OQ731723 (for ITS) | IPEE_Parasites 14310 |
| OQ730416 (for COX1) | IPEE_Parasites 14310 | ||
| OQ746340 (for LSU) | IPEE_Parasites 14310 | ||
| 12 | Elaphostrongylus rangiferi 4 | OQ731722 (for ITS2) | IPEE_Parasites 14309 |
| 13 | Ostertagia arctica 4 | OQ726410 (for ITS2) | IPEE_Parasites 14311 |
| 13 | Nematodirella longissimespiculata 4 | OQ726409 (for ITS2) | IPEE_Parasites 14312 |
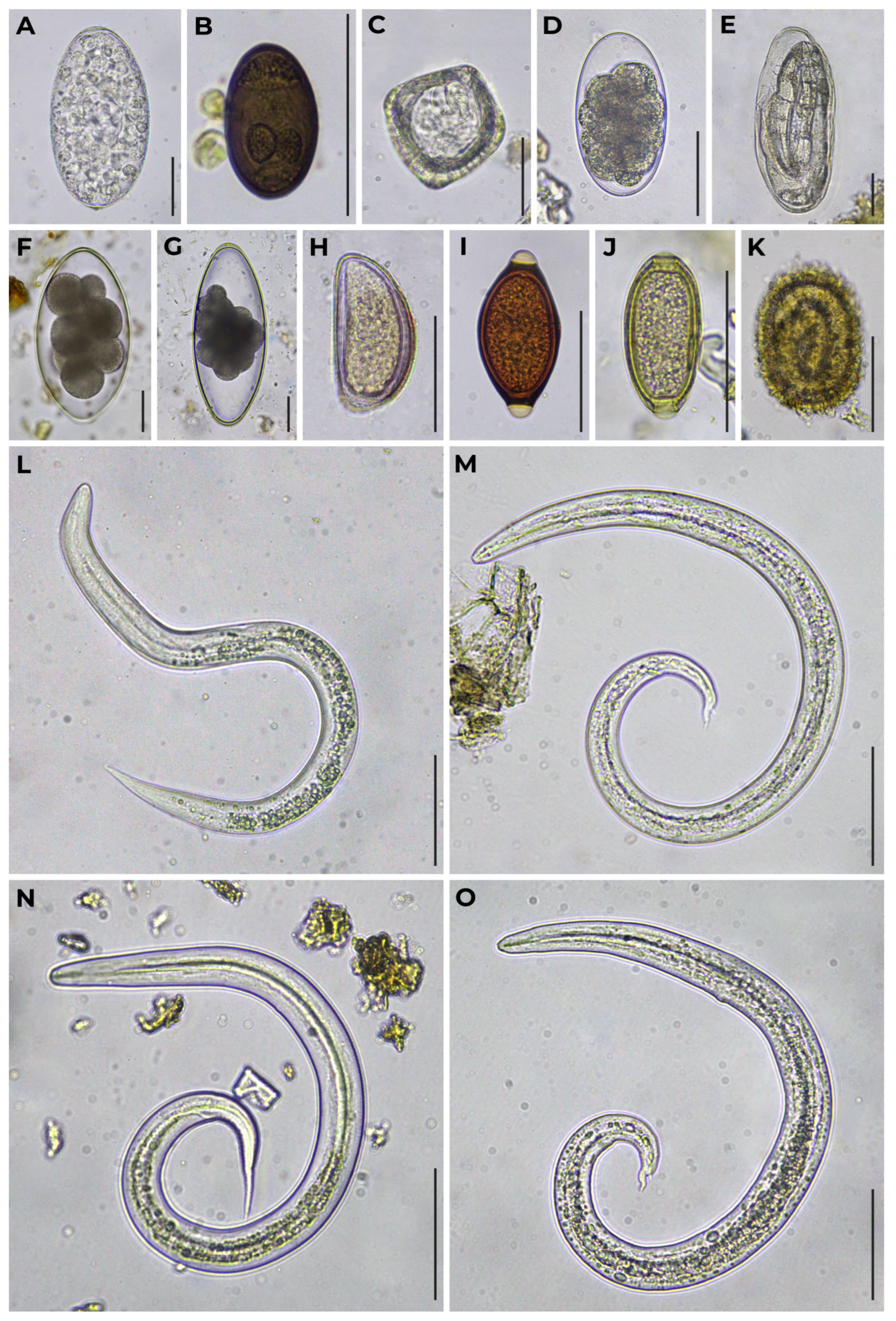
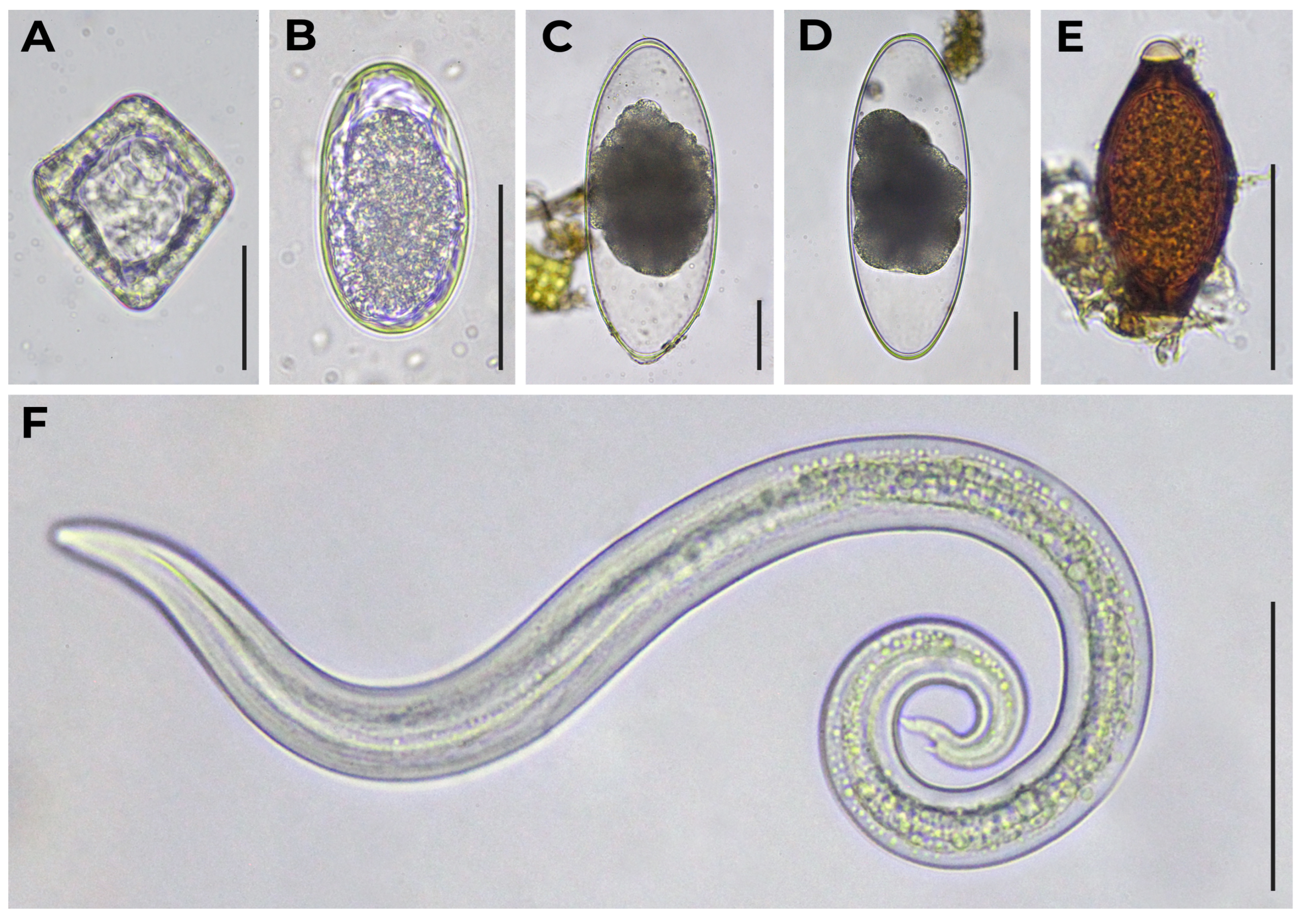
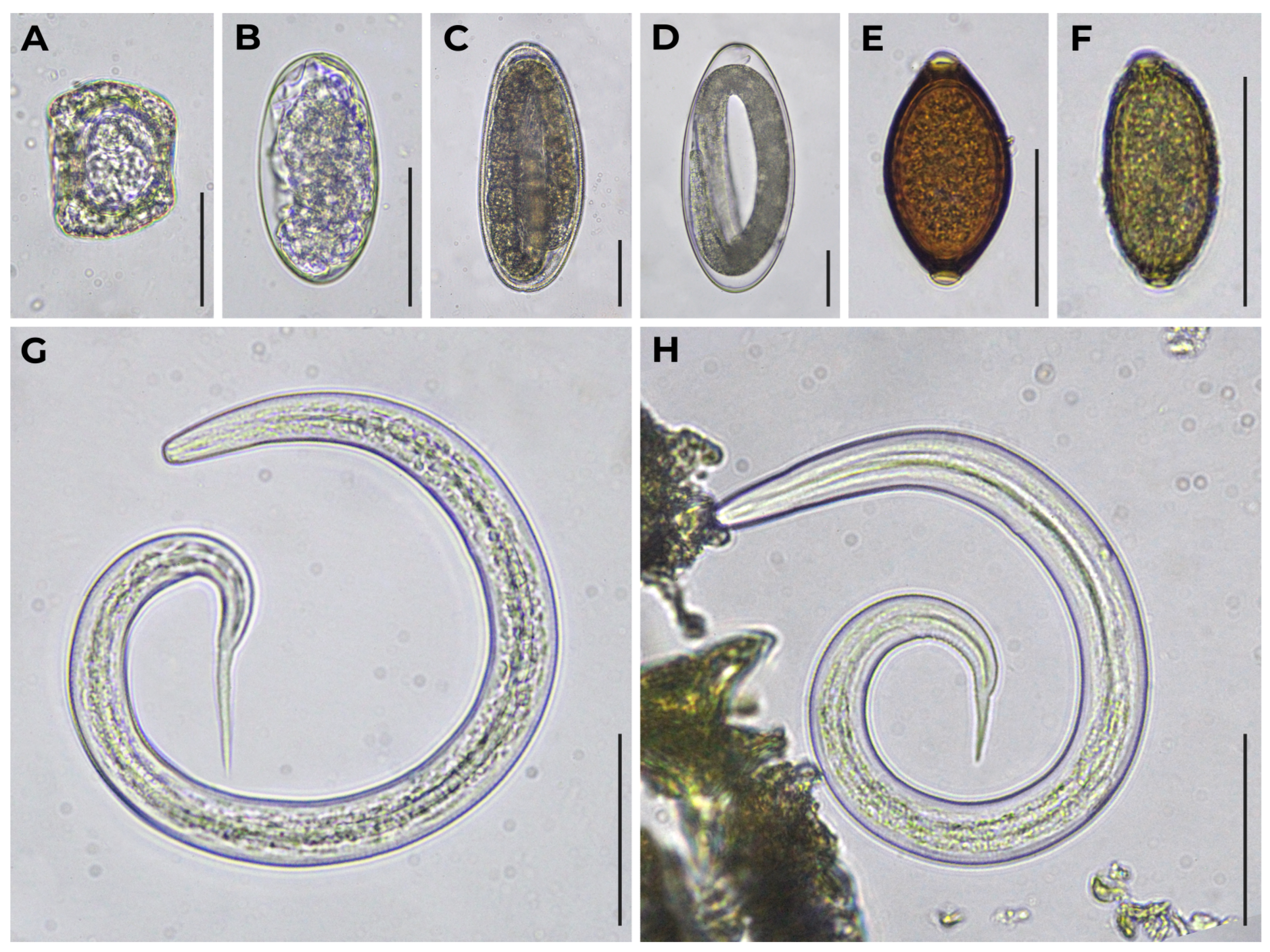
| Sample Set ID | Host Species | Trematodes | Cestodes | Nematodes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Paramphistomum sp. | Dicrocoelium sp. | Moniezia sp. | Strongyle-Type | Marshallagia sp. | Nematodirus sp. | Nematodirella sp. | Dictyocaulus sp. | Protostrongylidae | Skrjabinema sp. | Trichuris sp. | Capillarid-Type | Ascaris-Type | ||
| 1 | Wild reindeer (Rangifer tarandus) | 9 (47%) | – | – | 15 (79%) | – | 1 (5%) | – | – | 1 (5%) 3 3 (16%) 4 | – | – | – | – |
| 2 | – | – | – | 1 (33%) | – | – | – | – | – | – | – | – | – | |
| 3 | – | – | – | 1 (50%) | – | – | – | – | – | – | – | – | – | |
| 4 | – | – | – | 11 (73%) | 1 (7%) | – | – | – | – | – | – | – | – | |
| 5 | – | – | – | 14 (67%) | – | – | – | – | 4 (19%) 5 | – | – | – | – | |
| 6 | – | – | – | 11 (100%) | – | – | – | – | – | – | 1 (9%) | – | – | |
| 7 | Semiwild reindeer (R. tarandus) | – | – | 2 (4%) | 44 (88%) | – | 1 (2%) | – | – | 6 (12%) 5 | – | – | – | – |
| 8 | – | 1 (2%) | 16 (27%) | 47 (80%) | – | 18 (31%) | 11 (19%) | – | 12 (21%) 5 | 2 (3%) 9 | – | – | – | |
| 9 | – | – | 1 (50%) | 2 (100%) | – | – | – | – | – | – | 1 (50%) | – | – | |
| 10 | – | – | – | 15 (50%) | – | 7 (23%) | – | – | 21 (70%) 6 | – | – | 12 (40%) 10 | – | |
| 11 | 8 (26%) | – | – | 6 (19%) | – | – | – | 1 (3%) | 16 (52%) 5 | – | – | – | – | |
| 12 | 9 (45%) | – | – | 1 (5%) | – | – | – | 1 (5%) | 6 (30%) 6 | – | – | 3 (15%) 10 | – | |
| 13 | – | – | 2 (5%) | 27 (64%) 1 | – | 4 (10%) | 5 (12%) 2 | 2 (5%) | – | – | – | 14 (33%) 10 | – | |
| 14 | – | – | – | 4 (57%) | – | – | – | – | 4 (57%) 5 | – | – | 1 (14%) 10 | – | |
| 15 | – | – | – | 4 (40%) | – | – | – | – | – | – | – | – | – | |
| 16 | – | – | 2 (7%) | 18 (60%) | – | – | – | – | – | 2 (7%) 9 | – | – | 1 (3%) 11 | |
| 17 | – | – | – | – | – | 2 (25%) | 3 (38%) | – | – | – | – | – | – | |
| 18 | Muskoxen (Ovibos moschatus) | – | – | 2 (6%) | – | – | 1 (3%) | – | – | 19 (54%) 5 | – | 1 (3%) | – | – |
| 19 | – | – | 4 (17%) | 3 (13%) | – | – | 1 (4%) | – | – | – | – | – | – | |
| 20 | – | – | – | 2 (33%) | – | – | – | – | – | – | – | – | – | |
| 21 | – | – | – | 6 (24%) | – | – | – | – | – | – | – | – | – | |
| 22 | – | – | 1 (3%) | 9 (30%) | – | 1 (3%) | – | – | 8 (26%) 5 | – | – | – | – | |
| 23 | Snow sheep (Ovis nivicola) | – | – | – | – | 4 (19%) | – | – | – | 16 (76%) 7 | – | 4 (19%) | – | – |
| 24 | – | – | 2 (15%) | – | 7 (54%) | 1 (8%) | – | – | 9 (69%) 8 | – | 5 (39%) | 1 (8%) | – | |
| 25 | – | – | – | 1 (20%) | – | – | – | – | – | – | – | – | – | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loginova, O.A.; Rozenfeld, S.B.; Sipko, T.P.; Mizin, I.A.; Panchenko, D.V.; Laishev, K.A.; Bondar, M.G.; Kolpashchikov, L.A.; Gruzdev, A.R.; Kulemeev, P.S.; et al. Diversity and Distribution of Helminths in Wild Ruminants of the Russian Arctic: Reindeer (Rangifer tarandus), Muskoxen (Ovibos moschatus), and Snow Sheep (Ovis nivicola). Diversity 2023, 15, 672. https://doi.org/10.3390/d15050672
Loginova OA, Rozenfeld SB, Sipko TP, Mizin IA, Panchenko DV, Laishev KA, Bondar MG, Kolpashchikov LA, Gruzdev AR, Kulemeev PS, et al. Diversity and Distribution of Helminths in Wild Ruminants of the Russian Arctic: Reindeer (Rangifer tarandus), Muskoxen (Ovibos moschatus), and Snow Sheep (Ovis nivicola). Diversity. 2023; 15(5):672. https://doi.org/10.3390/d15050672
Chicago/Turabian StyleLoginova, Olga A., Sofya B. Rozenfeld, Taras P. Sipko, Ivan A. Mizin, Danila V. Panchenko, Kasim A. Laishev, Mikhail G. Bondar, Leonid A. Kolpashchikov, Aleksandr R. Gruzdev, Pavel S. Kulemeev, and et al. 2023. "Diversity and Distribution of Helminths in Wild Ruminants of the Russian Arctic: Reindeer (Rangifer tarandus), Muskoxen (Ovibos moschatus), and Snow Sheep (Ovis nivicola)" Diversity 15, no. 5: 672. https://doi.org/10.3390/d15050672