


Acanthocephalan Diversity and Host Associations Revealed from a Large-Scale Biodiversity Survey

Abstract
:1. Introduction
2. Materials and Methods
2.1. Host and Acanthocephalan Collection
2.2. Molecular Data
2.3. Morphological Data
3. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acanthocephalan Species | Host | Stage | Host Type | New Record | Ref |
|---|---|---|---|---|---|
| Andracantha leucocarboi | Little shag, Microcarbo melanleucos brevirostris | Ad | D | No | [15,31] |
| Otago shag Leucocarbo chalconotus | Ad | D | No | [15,31] | |
| Spotted shag Phalacrocorax punctatus | Ad | D | No | [15,31] | |
| Andracantha sigma | Little blue penguin Eudyptula novaehollandiae | Ad | D | No | [15,31,32] |
| Otago shag L. chalconotus | Ad | D | No | [15,31] | |
| Spotted shag P. punctatus | Ad | D | No | [15,31] | |
| Sprat Sprattus antipodum | Cy | P | No | [22] | |
| Aspersentis sp. | NZ sole Peltorhampus novaezeelandiae | Ad | D | Yes | |
| Bolbosoma balaenae | Little blue penguin E. novaehollandiae | Ad | A | No | [32] |
| Sprat S. antipodum | Cy | P | No | [22] | |
| Amphipod Themisto sp. | Cy | I | No | [21,22] | |
| Corynosoma hannae | Leopard seal Hydrurga leptonyx | Ad | D | No | [22] |
| Otago shag L. chalconotus | Ad | A | No | [22] | |
| Spotted shag P. punctatus | Ad | A | No | [22] | |
| Yellow-eyed penguin Megadyptes antipodes | Ad | A | No | [22] | |
| Banded wrasse Pseudolabrus fucicola | Cy | P | No | [22] | |
| Blue cod Parapercis colias | Cy | P | No | [22] | |
| Brill Colistium guntheri | Cy | P | No | [22] | |
| Crested bellowsfish Notopogon lilliei | Cy | P | No | [22] | |
| Lemon sole Pelotretis flavilatus | Cy | P | No | [22] | |
| NZ sole P. novaezeelandiae | Cy | P | No | [22] | |
| Pigfish Congiopodus leucopaecilus | Cy | P | No | [22] | |
| Scaly gurnard Lepidotrigla brachyoptera | Cy | P | No | [22] | |
| Scarlet wrasse Pseudolabrus miles | Cy | P | No | [22] | |
| Sprat S. antipodum | Cy | P | No | [22] | |
| Stargazer Genyagnus monopterygius | Cy | P | No | [22] | |
| Tarahiki Nemadactylus macropterus | Cy | P | No | [22] | |
| Echinorhynchus sp. | Pigfish C. leucopaecilus | Ad | D | Yes | |
| Gorgorhynchoides queenslandensis | Lemon sole P. flavilatus | Ad | D | Yes | |
| Neoechinorhynchus sp. | Mullet Aldrichetta forsteri | Ad | D | Yes | |
| Sprat S. antipodum | Ad | D | Yes | ||
| Plagiorhynchus allisonae | Pied stilt Himantopus himantopus | Ad | D | Yes | |
| South Island pied oystercatcher Haematopus finschi | Ad | D | No | [33] | |
| Amphipod Transorchestia serrulata | Cy | I | No | [21,22] | |
| Profilicollis novaezealandensis | Black-backed gull Larus dominicanus | Ad | D | No | [22,34] |
| Little shag M. melanleucos brevirostris | Ad | D | No | [15,22] | |
| Royal spoonbill Platalea regia | Ad | D | No | [22] | |
| Spotted shag P. punctatus | Ad | D | No | [15,22] | |
| Purple shore crab Hemigrapsus sexdentatus | Cy | I | No | [21,22] | |
| Stalk-eyed mud crab Hemiplax hirtipes | Cy | I | No | [21,22] | |
| Rhadinorhynchus sp. | Flesh-footed shearwater Puffinus carneipes | Ad | A | Yes | |
| Tegorhynchus sp. | Scarlet wrasse P. miles | Ad | D | Yes | |
| Tenuisoma tarapungi | Black-backed gull L. dominicanus | Ad | D | Yes | |
| Red-billed gull Chroicocephalus scopulinus | Ad | D | No | [35] |
| Acanthocephalan Species | Host | Isolate | Genbank Accession | ||
|---|---|---|---|---|---|
| 28S | 18S | cox1 | |||
| Aspersentis sp. | NZ sole | FF1aca1_10 | OQ947383 | OQ942219 | OQ947286 |
| Corynosoma hannae | Leopard seal | LSL1aca1 | OQ947384 | ||
| Otago shag | OSH1aca1 | OQ947385 | |||
| Spotted shag | SSH1aca1 | OQ947386 | |||
| Yellow-eyed penguin | YEP1aca1 | OQ947387 | |||
| Gorgorhynchoides queenslandensis | Lemon sole | FF2aca2_17 | OQ947388 | OQ942220 | |
| Neoechinorhynchus sp. | Mullet | Mull1aca1 | OQ947389 | OQ947287 | |
| Sprat | SPRAaca4 | OQ947390 | |||
| Plagiorhynchus allisonae | Pied stilt | Psti1aca1 | OQ947391 | ||
| Profilicollis novaezealandensis | Royal spoonbill | SPB1aca1 | OQ947392 | ||
| Tegorhynchus sp. | Scarlet wrasse | Fish3Baca1 | OQ947393 | OQ942221 | |
| Tenuisoma tarapungi | Black-backed gull | BBG40aca1 | OQ947394 | ||

3.1. Class Palaeacanthocephala: Order Polymorphida Petrochenko, 1956: Family Polymorphidae Meyer, 1931
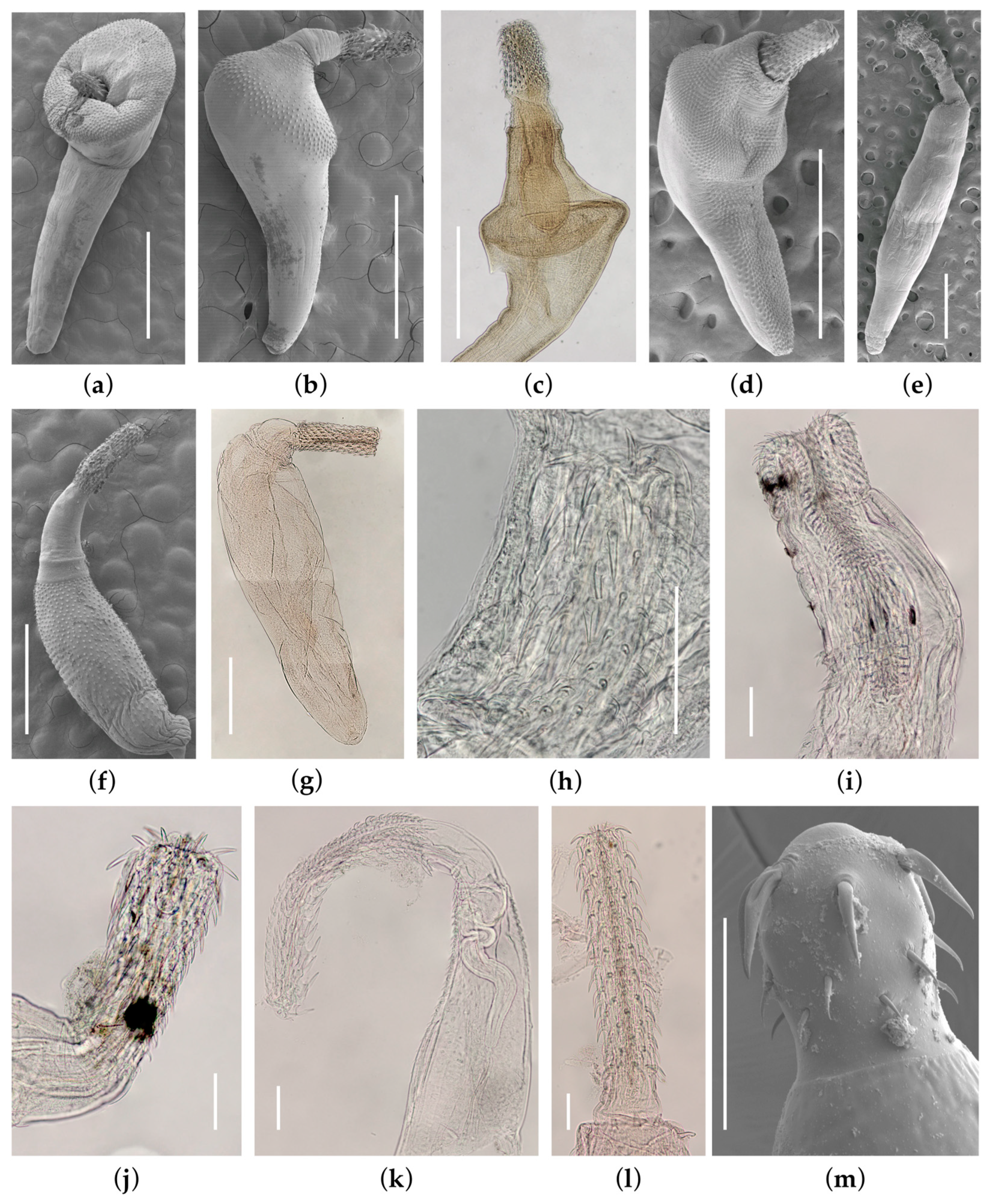
3.1.1. Andracantha leucocarboi Presswell, García-Varela & Smales, 2018 (Figure 1a)
3.1.2. Andracantha sigma Presswell, García-Varela & Smales, 2018 (Figure 1b)
3.1.3. Bolbosoma balaenae (Gmelin, 1790) Porta, 1908 (Figure 1c)
3.1.4. Corynosoma hannae Zdzitowiecki, 1984 (Figure 1d)
3.1.5. Profilicollis novaezelandensis Brockerhoff & Smales, 2002 (Figure 1e)
3.1.6. Tenuisoma tarapungi Presswell, Bennett & Smales, 2020 (Figure 1f)
3.2. Class Palaeacanthocephala: Order Polymorphida Petrochenko, 1956: Family Plagiorhynchidae Golvan, 1960
Plagiorhynchus allisonae Smales 2002 (Figure 1g)
3.3. Class Palaeacanthocephala: Order Echinorhynchida Southwell & Macfie, 1925: Family Echinorhynchidae Cobbold, 1876
Echinorhynchus sp. (Figure 1j)
3.4. Class Palaeacanthocephala: Order Echinorhynchida Southwell & Macfie, 1925: Family Heteracanthocephalidae Petrochenko, 1956
Aspersentis sp. (Figure 1h)
3.5. Class Palaeacanthocephala: Order Echinorhynchida Southwell & Macfie, 1925: Family Isthmosacanthidae Smales, 2012
Gorgorhynchoides queenslandensis Smales, 2014 (Figure 1i)
3.6. Class Palaeacanthocephala: Order Echinorhynchida Southwell & Macfie, 1925: Family Leptorhynchoididae Witenberg, 1923
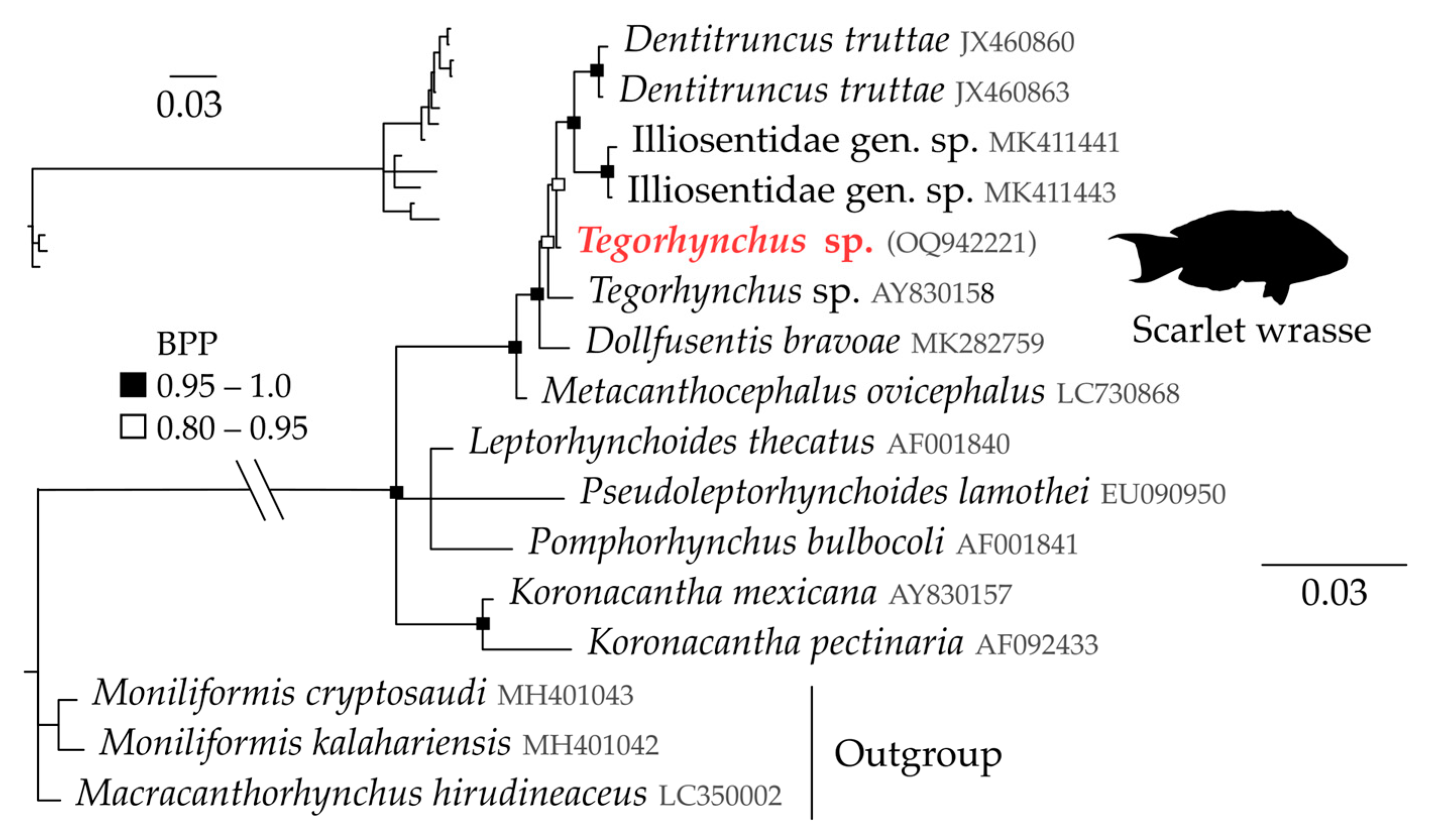
Tegorhynchus sp. (Figure 1k and Figure 3)

3.7. Class Palaeacanthocephala: Order Echinorhynchida Southwell & Macfie, 1925: Family Rhadinorhynchidae Lühe, 1911
Rhadinorhynchus sp. (Figure 1l)
3.8. Class Eoacanthocephala: Order Neoechinorhynchida Southwell & Macfie, 1925: Family Neoechinorhynchidae (Ward, 1917)
Neoechinorhynchus sp. (Figure 1m and Figure 4)

4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Cowart, D.A.; Pinheiro, M.; Mouchel, O.; Maguer, M.; Grall, J.; Miné, J.; Arnaud-Haond, S. Metabarcoding Is Powerful yet Still Blind: A Comparative Analysis of Morphological and Molecular Surveys of Seagrass Communities. PLoS ONE 2015, 10, e0117562. [Google Scholar] [CrossRef] [PubMed]
- Titley, M.A.; Snaddon, J.L.; Turner, E.C. Scientific Research on Animal Biodiversity Is Systematically Biased towards Vertebrates and Temperate Regions. PLoS ONE 2017, 12, e0189577. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, K.D.; Dobson, A.P.; Kuris, A.M. Parasites Dominate Food Web Links. Proc. Natl. Acad. Sci. USA 2006, 103, 11211–11216. [Google Scholar] [CrossRef]
- Wood, C.L.; Byers, J.E.; Cottingham, K.L.; Altman, I.; Donahue, M.J.; Blakeslee, A.M.H. Parasites Alter Community Structure. Proc. Natl. Acad. Sci. USA 2007, 104, 9335–9339. [Google Scholar] [CrossRef]
- Kuris, A.M.; Hechinger, R.F.; Shaw, J.C.; Whitney, K.L.; Aguirre-Macedo, L.; Boch, C.A.; Dobson, A.P.; Dunham, E.J.; Fredensborg, B.L.; Huspeni, T.C.; et al. Ecosystem Energetic Implications of Parasite and Free-Living Biomass in Three Estuaries. Nature 2008, 454, 515–518. [Google Scholar] [CrossRef]
- Thompson, R.M.; Mouritsen, K.N.; Poulin, R. Importance of Parasites and Their Life Cycle Characteristics in Determining the Structure of a Large Marine Food Web. J. Anim. Ecol. 2005, 74, 77–85. [Google Scholar] [CrossRef]
- Dunne, J.A.; Lafferty, K.D.; Dobson, A.P.; Hechinger, R.F.; Kuris, A.M.; Martinez, N.D.; McLaughlin, J.P.; Mouritsen, K.N.; Poulin, R.; Reise, K.; et al. Parasites Affect Food Web Structure Primarily through Increased Diversity and Complexity. PLoS Biol. 2013, 11, e1001579. [Google Scholar] [CrossRef]
- Sato, T.; Egusa, T.; Fukushima, K.; Oda, T.; Ohte, N.; Tokuchi, N.; Watanabe, K.; Kanaiwa, M.; Murakami, I.; Lafferty, K.D. Nematomorph Parasites Indirectly Alter the Food Web and Ecosystem Function of Streams through Behavioural Manipulation of Their Cricket Hosts. Ecol. Lett. 2012, 15, 786–793. [Google Scholar] [CrossRef]
- Kennedy, C.R. Ecology of the Acanthocephala; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Haye, P.A.; Ojeda, F.P. Metabolic and Behavioral Alterations in the Crab Hemigrapsus crenulatus (Milne-Edwards 1837) Induced by Its Acanthocephalan Parasite Profilicollis antarcticus (Zdzitowiecki 1985). J. Exp. Mar. Biol. Ecol. 1998, 228, 73–82. [Google Scholar] [CrossRef]
- Latham, A.; Poulin, R. Effect of Acanthocephalan Parasites on the Behaviour and Coloration of the Mud Crab Macrophthalmus hirtipes (Brachyura: Ocypodidae). Mar. Biol. 2001, 139, 1147–1154. [Google Scholar] [CrossRef]
- Richardson, D.J.; Nickol, B.B. Acanthocephala. In Parasitic Diseases of Wild Birds; Atkinson, C.T., Thomas, N.J., Hunter, D.B., Eds.; Wiley-Blackwell: Oxford, UK, 2009; pp. 277–288. ISBN 978-0-8138-0462-0. [Google Scholar]
- Shanebeck, K.M.; Thacker, C.; Lagrue, C. Corynosoma strumosum (Acanthocephala) Infection in Marine Foraging Mink (Neogale vison) and River Otter (Lontra canadensis) and Associated Peritonitis in a Juvenile Mink. Parasitol. Int. 2022, 89, 102579. [Google Scholar] [CrossRef]
- Presswell, B.; Bennett, J. Helminth Parasites of Shags (Phalacrocoracidae) from the Otago Region of Southern New Zealand. J. Helminthol. 2021, 95, e9. [Google Scholar] [CrossRef]
- Bennett, J.; Presswell, B.; Poulin, R. Biodiversity of Marine Helminth Parasites in New Zealand: What Don’t We Know? N. Z. J. Mar. Freshw. Res. 2022, 56, 175–190. [Google Scholar] [CrossRef]
- Bennett, J.; Poulin, R.; Presswell, B. Large-Scale Genetic Investigation of Nematode Diversity and Their Phylogenetic Patterns in New Zealand’s Marine Animals. Parasitology 2022, 149, 1794–1809. [Google Scholar] [CrossRef]
- Faltýnková, A.; Georgieva, S.; Kostadinova, A.; Bray, R.A. Biodiversity and Evolution of Digeneans of Fishes in the Southern Ocean. In Biodiversity and Evolution of Parasitic Life in the Southern Ocean; Klimpel, S., Kuhn, T., Mehlhorn, H., Eds.; Parasitology Research Monographs; Springer International Publishing: Cham, Switzerland, 2017; Volume 9, pp. 49–75. ISBN 978-3-319-46342-1. [Google Scholar]
- Mattiucci, S.; Cipriani, P.; Paoletti, M.; Levsen, A.; Nascetti, G. Reviewing Biodiversity and Epidemiological Aspects of Anisakid Nematodes from the North-East Atlantic Ocean. J. Helminthol. 2017, 91, 422–439. [Google Scholar] [CrossRef]
- Santacruz, A.; Barluenga, M.; Pérez-Ponce de León, G. The Macroparasite Fauna of Cichlid Fish from Nicaraguan Lakes, a Model System for Understanding Host–Parasite Diversification and Speciation. Sci. Rep. 2022, 12, 3944. [Google Scholar] [CrossRef]
- Bennett, J.; Poulin, R.; Presswell, B. Annotated Checklist and Genetic Data for Parasitic Helminths Infecting New Zealand Marine Invertebrates. Invertebr. Biol. 2022, 141, e12380. [Google Scholar] [CrossRef]
- Bennett, J.; Presswell, B.; Poulin, R. Tracking Life Cycles of Parasites across a Broad Taxonomic Scale in a Marine Ecosystem. Int. J. Parasitol. 2023; in press. [Google Scholar] [CrossRef]
- Otago Regional Council. Regional Plan: Coast of Otago; Otago Regional Council: Dunedin, New Zealand, 2012; p. 196. [Google Scholar]
- Harper, J.T.; Saunders, G.W. Molecular Systematics of the Florideophyceae (Rhodophyta) Using Nuclear Large and Small Subunit RDNA Sequence Data. J. Phycol. 2001, 37, 1073–1082. [Google Scholar] [CrossRef]
- Bowles, J.; Blair, D.; Mcmanus, D. Genetic Variants within the Genus Echinococcus Identified by Mitochondrial DNA Sequencing. Mol. Biochem. Parasitol. 1992, 54, 165–173. [Google Scholar] [CrossRef]
- Littlewood, D.T.J.; Olson, P.D. Small Subunit RDNA and the Platyhelminthes: Signal, Noise, Conflict and Compromise. In Interrelationships of the Platyhelminthes; CRC Press: London, UK, 2001; p. 376. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2010. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Presswell, B.; García-Varela, M.; Smales, L.R. Morphological and Molecular Characterization of Two New Species of Andracantha (Acanthocephala: Polymorphidae) from New Zealand Shags (Phalacrocoracidae) and Penguins (Spheniscidae) with a Key to the Species. J. Helminthol. 2018, 92, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.; McPherson, O.; Presswell, B. Gastrointestinal Helminths of Little Blue Penguins, Eudyptula novaehollandiae (Stephens), from Otago, New Zealand. Parasitol. Int. 2021, 80, 102185. [Google Scholar] [CrossRef] [PubMed]
- Smales, L.R. Plagiorhynchidae Meyer, 1931 (Acanthocephala) from Australasian Birds and Mammals, with Descriptions of Plagiorhynchus (Plagiorhynchus) menurae (Johnston, 1912) and P. (P.) allisonae n. Sp. Syst. Parasitol. 2002, 51, 207–216. [Google Scholar] [CrossRef]
- Latham, A.D.M.; Poulin, R. New Records of Gastrointestinal Helminths from the Southern Black-backed Gull (Larus dominicanus) in New Zealand. N. Z. J. Zool. 2002, 29, 253–257. [Google Scholar] [CrossRef]
- Presswell, B.; Bennett, J.D.L.; Smales, L.R. Morphological and Molecular Characterisation of a New Genus and Species of Acanthocephalan, Tenuisoma tarapungi n. g., n. Sp. (Acanthocephala: Polymorphidae) Infecting Red-Billed Gulls in New Zealand, with a Key to the Genera of the Polymorphidae Meyer, 1931. Syst. Parasitol. 2020, 97, 25–39. [Google Scholar] [CrossRef]
- Hernández-Orts, J.S.; Smales, L.R.; Pinacho-Pinacho, C.D.; García-Varela, M.; Presswell, B. Novel Morphological and Molecular Data for Corynosoma hannae Zdzitowiecki, 1984 (Acanthocephala: Polymorphidae) from Teleosts, Fish-Eating Birds and Pinnipeds from New Zealand. Parasitol. Int. 2017, 66, 905–916. [Google Scholar] [CrossRef]
- García-Varela, M.; Pérez-Ponce de León, G.; Aznar, F.J.; Nadler, S.A. Phylogenetic Relationship among Genera of Polymorphidae (Acanthocephala), Inferred from Nuclear and Mitochondrial Gene Sequences. Mol. Phylogenet. Evol. 2013, 68, 176–184. [Google Scholar] [CrossRef]
- Santoro, M.; Palomba, M.; Gili, C.; Marcer, F.; Marchiori, E.; Mattiucci, S. Molecular and Morphological Characterization of Bolbosoma balaenae (Acanthocephala: Polymorphidae), a Neglected Intestinal Parasite of the Fin Whale Balaenoptera physalus. Parasitology 2021, 148, 1293–1302. [Google Scholar] [CrossRef]
- Gregori, M.; Aznar, F.; Abollo, E.; Roura, Á.; González, Á.; Pascual, S. Nyctiphanes couchii as Intermediate Host for the Acanthocephalan Bolbosoma balaenae in Temperate Waters of the NE Atlantic. Dis. Aquat. Organ. 2012, 99, 37–47. [Google Scholar] [CrossRef]
- Zdzitowiecki, K. Some Antarctic Acanthocephalans of the Genus Corynosoma Parasitizing Pinnipedia, with Descriptions of Three New Species. Acta Parasitol. Pol. 1984, 29, 359–377. [Google Scholar]
- Aznar, F.J.; Hernández-Orts, J.; Suárez, A.A.; García-Varela, M.; Raga, J.A.; Cappozzo, H.L. Assessing Host–Parasite Specificity through Coprological Analysis: A Case Study with Species of Corynosoma (Acanthocephala: Polymorphidae) from Marine Mammals. J. Helminthol. 2012, 86, 156–164. [Google Scholar] [CrossRef]
- Anglade, T.; Randhawa, H. Gaining Insights into the Ecological Role of the New Zealand Sole (Peltorhamphus novaezeelandiae) through Parasites. J. Helminthol. 2018, 92, 187–196. [Google Scholar] [CrossRef]
- Brockerhoff, A.M.; Smales, L.R. Profilicollis novaezelandensis n. Sp. (Polymorphidae) and Two Other Acanthocephalan Parasites from Shore Birds (Haematopodidae and Scolopacidae) in New Zealand, with Records of Two Species in Intertidal Crabs (Decapoda: Grapsidae and Ocypodidae). Syst. Parasitol. 2002, 52, 55–65. [Google Scholar] [CrossRef]
- Fredensborg, B.L.; Latham, A.D.; Poulin, R. New Records of Gastrointestinal Helminths from the Red-billed Gull (Larus novaehollandiae scopulinus). N. Z. J. Zool. 2004, 31, 75–80. [Google Scholar] [CrossRef]
- Lagrue, C.; Heaphy, K.; Presswell, B.; Poulin, R. Strong Association between Parasitism and Phenotypic Variation in a Supralittoral Amphipod. Mar. Ecol. Prog. Ser. 2016, 553, 111–123. [Google Scholar] [CrossRef]
- WoRMS Editorial Board World Register of Marine Species. Available online: https://www.marinespecies.org (accessed on 20 January 2022).
- Smales, L.R. Echinorhynchus lesteri N. SP. (Acanthocephala: Echinorhynchidae) Parasitic in the Intestine of the Orange Roughy Hoplostethus atlanticus. Trans. R. Soc. S. Aust. 2012, 136, 50–55. [Google Scholar] [CrossRef]
- Baylis, H.A. XLIII.—Three New Acanthocephala from Marine Fishes of Australasia. Ann. Mag. Nat. Hist. 1944, 11, 462–472. [Google Scholar] [CrossRef]
- Van Cleave, H.J. A New Genus and New Species of Acanthocephala from the Antarctic. Ann. Mag. Nat. Hist. 1929, 4, 229–331. [Google Scholar] [CrossRef]
- Pichelin, S.; Smales, L.; Bray, R.A. A Discussion on the Heteracanthocephalidae Petrochenko, 1956 (Acanthocephala: Palaeacanthocephala). Syst. Parasitol. 2002, 52, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, S.J.; Smales, L.R. A New Species of Acanthocephala from the Greenback Flounder, Rhombosolea tapirina Günther, 1862. Trans. R. Soc. S. Aust. 1992, 116, 35–38. [Google Scholar]
- Baylis, H.A. Parasitic Nematoda and Acanthocephala Collected in 1925–1927. Discov. Rep. 1929, 1, 541–560. [Google Scholar]
- Smales, L.R. A Redescription of Aspersentis zanclorhynchi (Johnston and Best, 1937) Comb. Nov. (Heteracanthocephalidae: Acanthocephala). Trans. R. Soc. S. Aust. 1996, 120, 167–171. [Google Scholar]
- Smales, L.R. New Species of Gorgorhynchoides (Acanthocephala: Isthmosacanthidae) from Australian Fish (Carangidae) with a Key to the Species of the Genus. Comp. Parasitol. 2014, 81, 240–247. [Google Scholar] [CrossRef]
- Kita, Y.; Hiruta, S.F.; Sasaki, M.; Kajihara, H. Systematic Position of the Genus Metacanthocephalus Yamaguti, 1959 (Palaeacanthocephala: Echinorhynchida) Inferred from Molecular Evidence, with a Redescription of Metacanthocephalus ovicephalus (Zhukov, 1960). Parasitol. Int. 2023, 94, 102731. [Google Scholar] [CrossRef]
- García-Varela, M.; Nadler, S.A. Phylogenetic Relationships of Palaeacanthocephala (Acanthocephala) Inferred from SSU and LSU RDNA Gene Sequences. J. Parasitol. 2005, 91, 1401–1409. [Google Scholar] [CrossRef]
- Smales, L. The Genus Rhadinorhynchus (Acanthocephala: Rhadinorhynchidae) from Marine Fish in Australia with the Description of Four New Species. Acta Parasitol. 2014, 59, 721–736. [Google Scholar] [CrossRef]
- Jones, J.B. Movements of Albacore Tuna (Thunnus alalunga) in the South Pacific: Evidence from Parasites. Mar. Biol. 1991, 111, 1–9. [Google Scholar] [CrossRef]
- Gibson, D.; Jones, J.B. Fed up with Parasites? A Method for Estimating Asymptotic Growth in Fish Populations. Mar. Biol. 1993, 117, 495–500. [Google Scholar] [CrossRef]
- Edmonds, S.J. Australian Acanthocephala No. 13: Three New Species. Trans. R. Soc. S. Aust. 1971, 95, 55–60. [Google Scholar]
- Boustead, N.C. Fish Diseases Recorded in New Zealand, with a Discussion on Potential Sources and Gertification Procedures; Fisheries Research Division Occasional Publication; New Zealand Ministry of Agriculture and Fisheries: Wellington, New Zealand, 1982. [Google Scholar]
- Huston, D.C.; Cutmore, S.C.; Smales, L.R. Molecular Characterisation and Updated Description of Neoechinorhynchus aldrichettae Edmonds, 1971 (Acanthocephala: Neoechinorhynchidae), Based on Material from Aldrichetta Forsteri (Valenciennes) Collected in Tasmania, Australia. Syst. Parasitol. 2022, 99, 241–251. [Google Scholar] [CrossRef]
- Pinacho-Pinacho, C.D.; Sereno-Uribe, A.L.; García-Varela, M. Morphological and Molecular Data Reveal a New Species of Neoechinorhynchus (Acanthocephala: Neoechinorhynchidae) from Dormitator maculatus in the Gulf of Mexico. Parasitol. Int. 2014, 63, 763–771. [Google Scholar] [CrossRef]
- Shanebeck, K.; Lakemeyer, J.; Siebert, U.; Lehnert, K. Novel Infections of Corynosoma enhydri and Profilicollis sp. (Acanthocephala: Polymorphidae) Identified in Sea Otters Enhydra lutris. Dis. Aquat. Organ. 2020, 137, 239–246. [Google Scholar] [CrossRef]
- Shanebeck, K.M.; Besson, A.A.; Lagrue, C.; Green, S.J. The Energetic Costs of Sub-lethal Helminth Parasites in Mammals: A Meta-analysis. Biol. Rev. 2022, 97, 1886–1907. [Google Scholar] [CrossRef]
- Thompson, R.M.; Poulin, R.; Mouritsen, K.N.; Thieltges, D.W. Resource Tracking in Marine Parasites: Going with the Flow? Oikos 2013, 122, 1187–1194. [Google Scholar] [CrossRef]
- Carlson, C.J.; Burgio, K.R.; Dougherty, E.R.; Phillips, A.J.; Bueno, V.M.; Clements, C.F.; Castaldo, G.; Dallas, T.A.; Cizauskas, C.A.; Cumming, G.S.; et al. Parasite Biodiversity Faces Extinction and Redistribution in a Changing Climate. Sci. Adv. 2017, 3, e1602422. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennett, J.; Poulin, R.; Presswell, B. Acanthocephalan Diversity and Host Associations Revealed from a Large-Scale Biodiversity Survey. Diversity 2023, 15, 665. https://doi.org/10.3390/d15050665
Bennett J, Poulin R, Presswell B. Acanthocephalan Diversity and Host Associations Revealed from a Large-Scale Biodiversity Survey. Diversity. 2023; 15(5):665. https://doi.org/10.3390/d15050665
Chicago/Turabian StyleBennett, Jerusha, Robert Poulin, and Bronwen Presswell. 2023. "Acanthocephalan Diversity and Host Associations Revealed from a Large-Scale Biodiversity Survey" Diversity 15, no. 5: 665. https://doi.org/10.3390/d15050665