


Hemogregarine Diversity Infecting Brazilian Turtles with a Description of Six New Species of Haemogregarina (Apicomplexa: Adeleorina: Haemogregarinidae)

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site and Animal Collection
2.2. Blood Smears, Histological Slides, and Morphological Data
2.3. Molecular Data
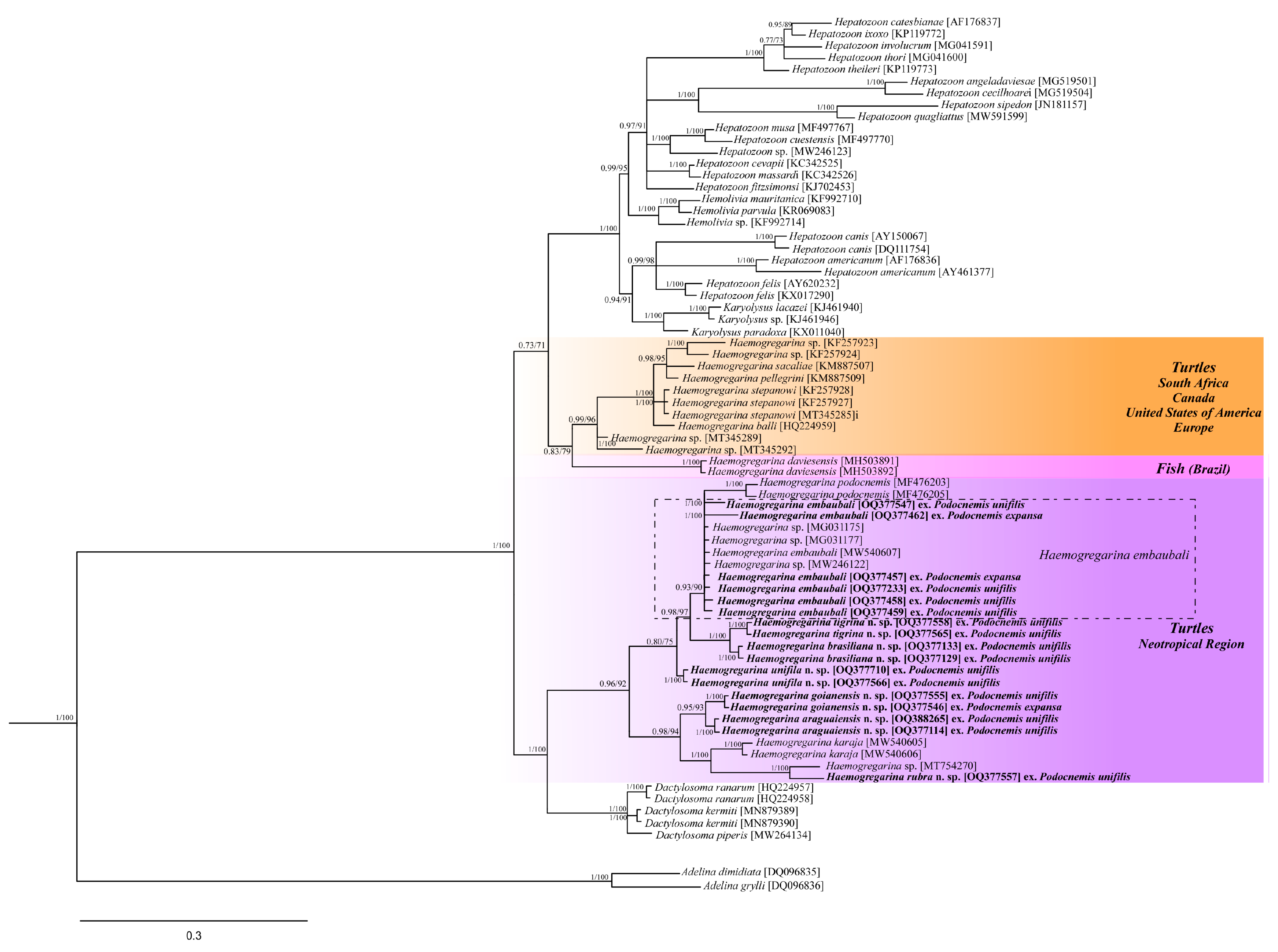
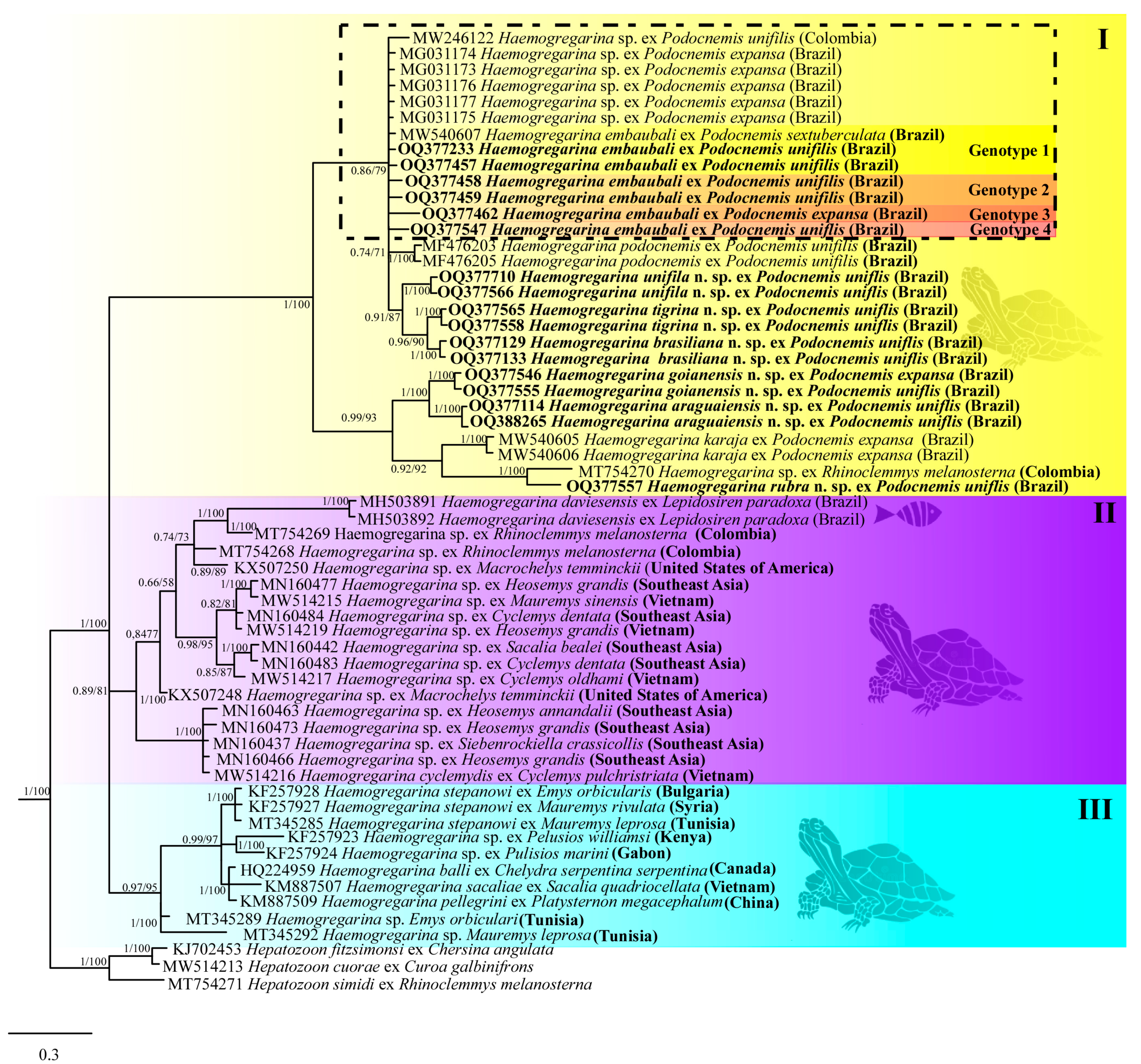
3. Results
3.1. Prevalence
3.2. Molecular Data
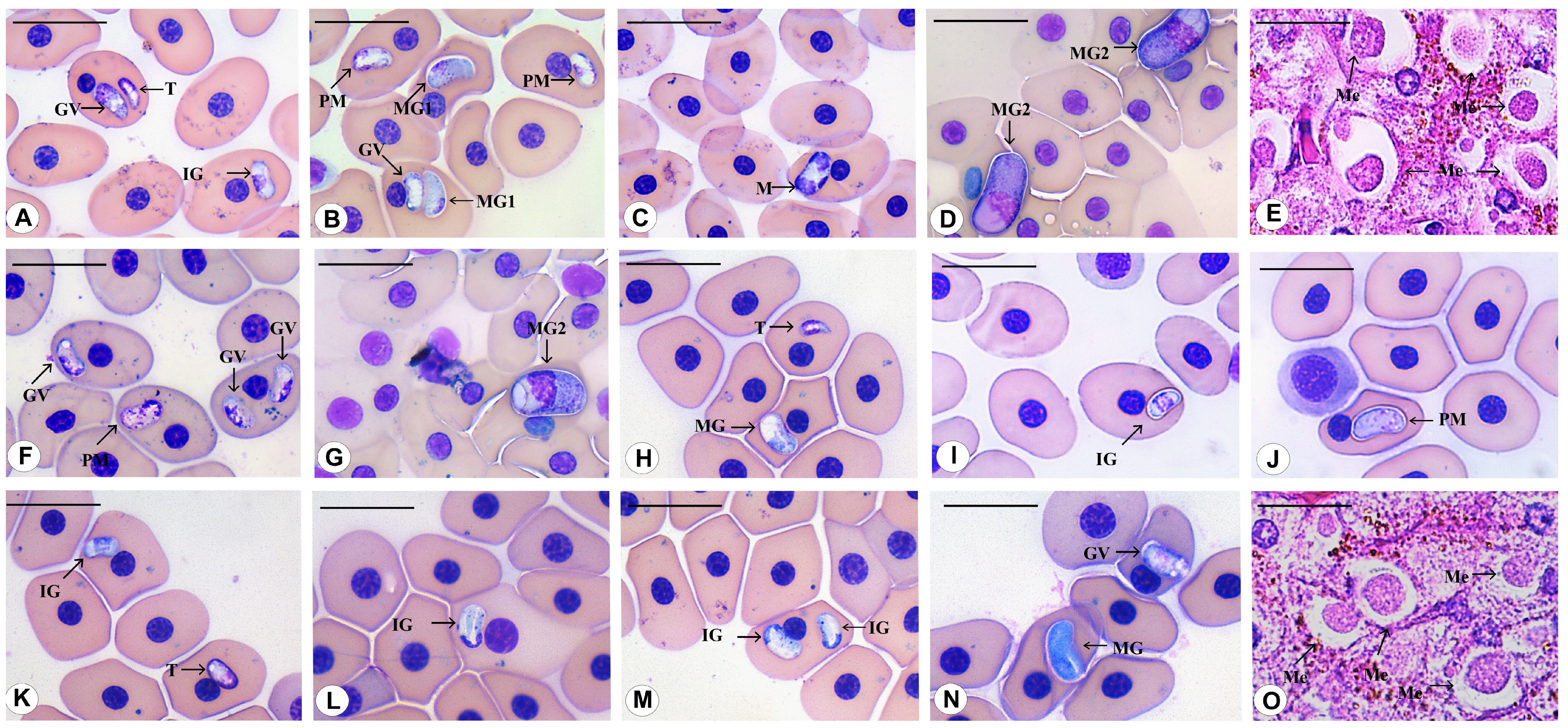
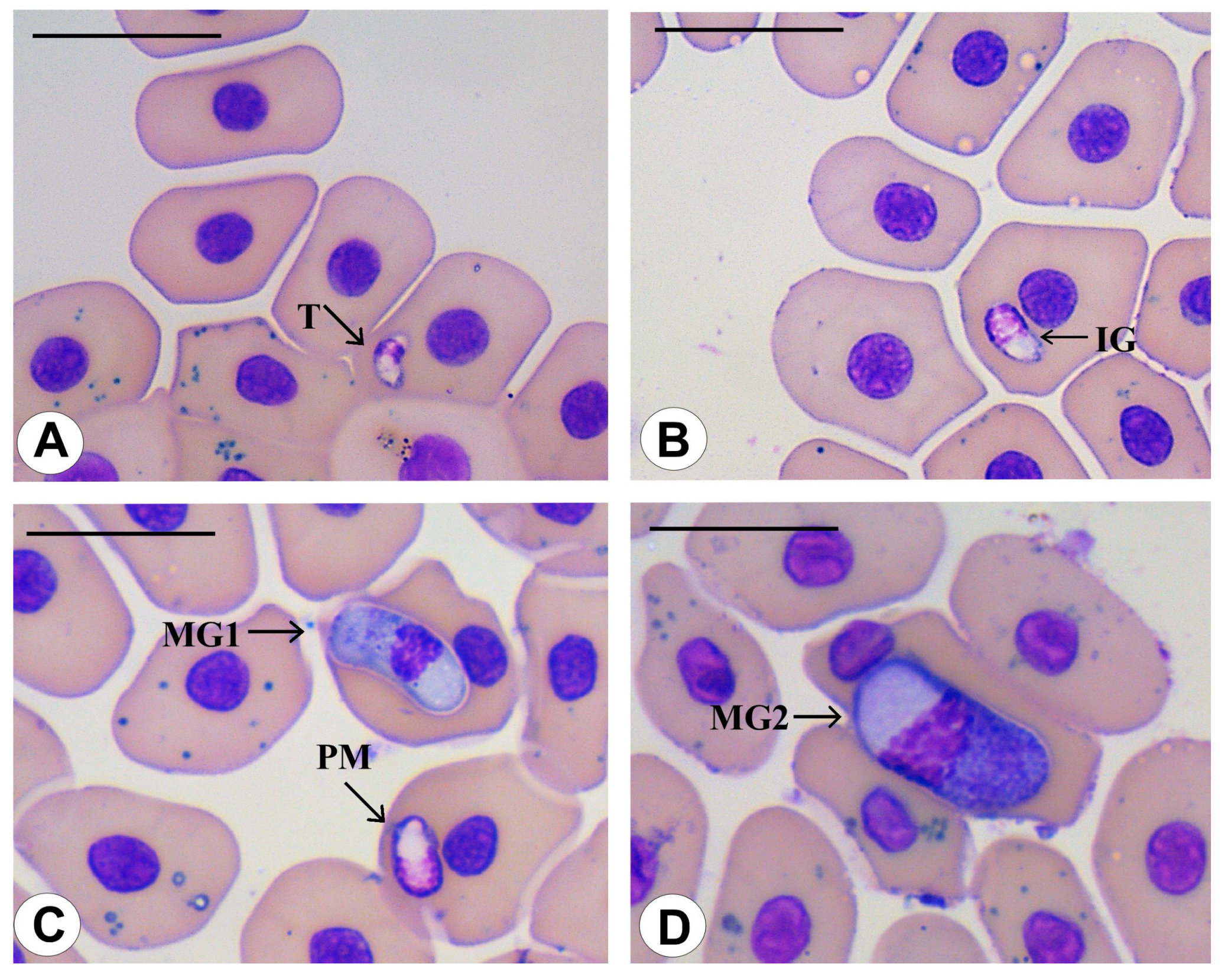
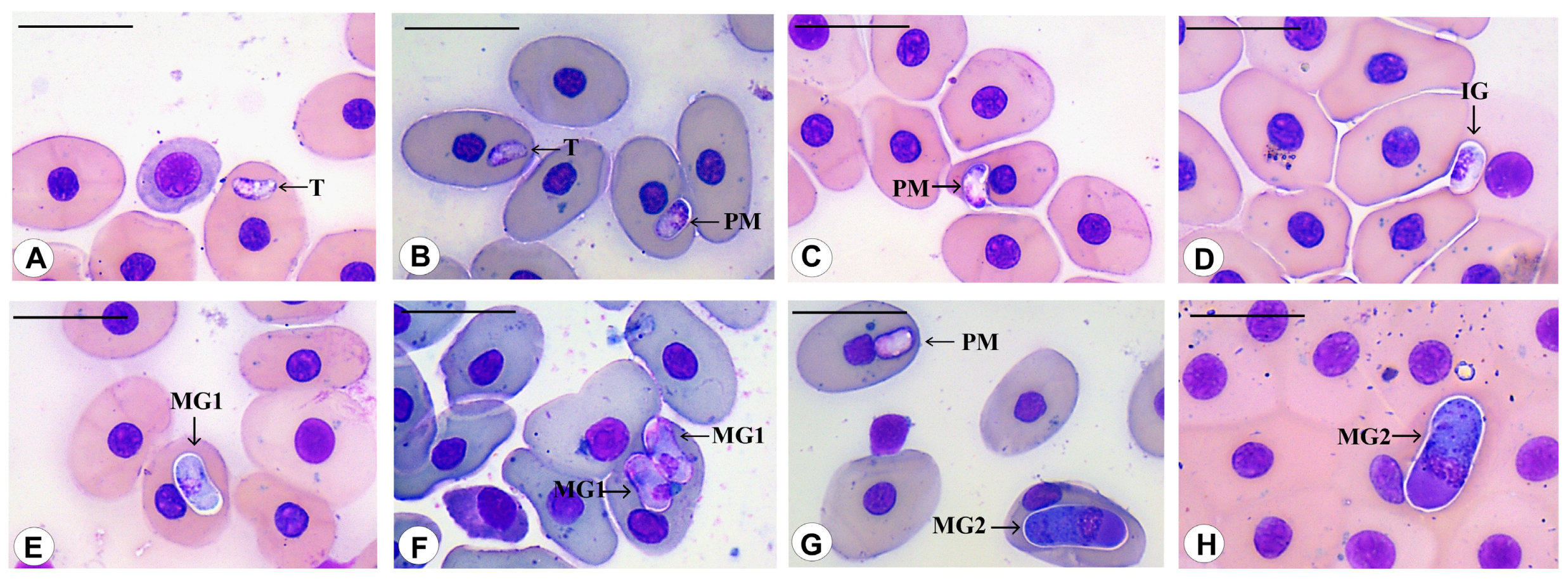
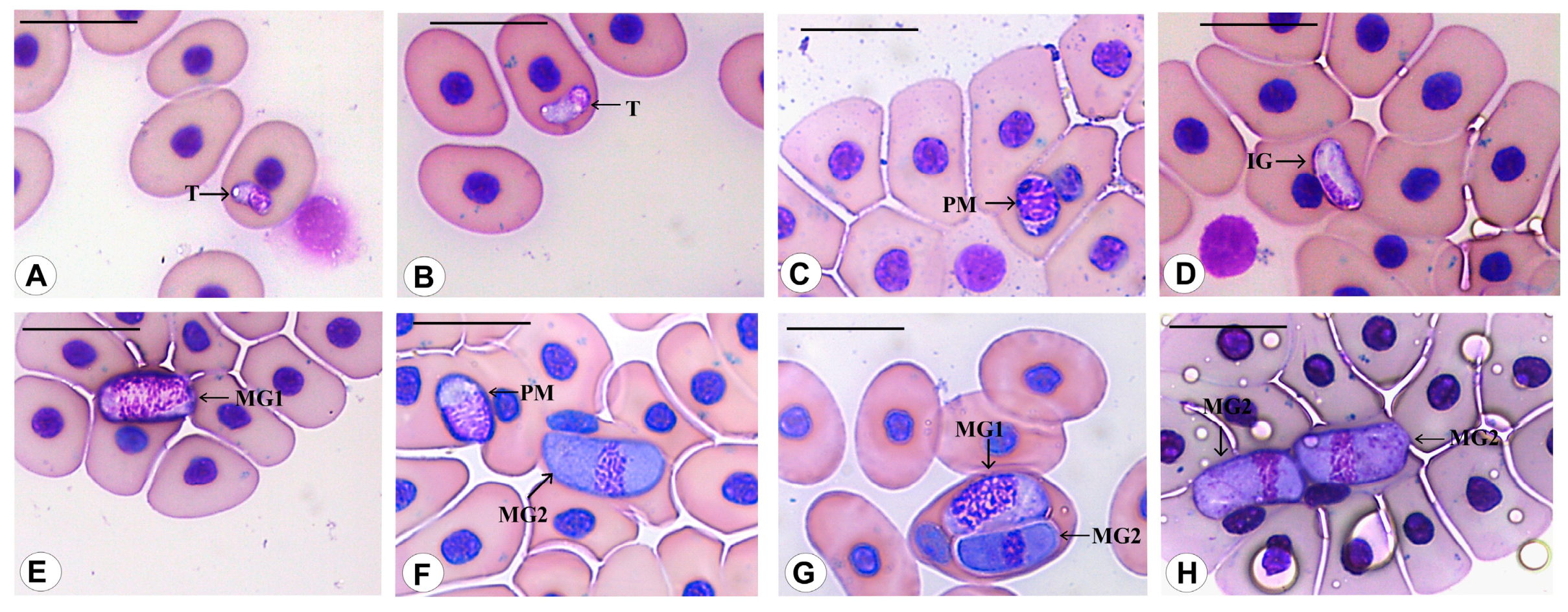
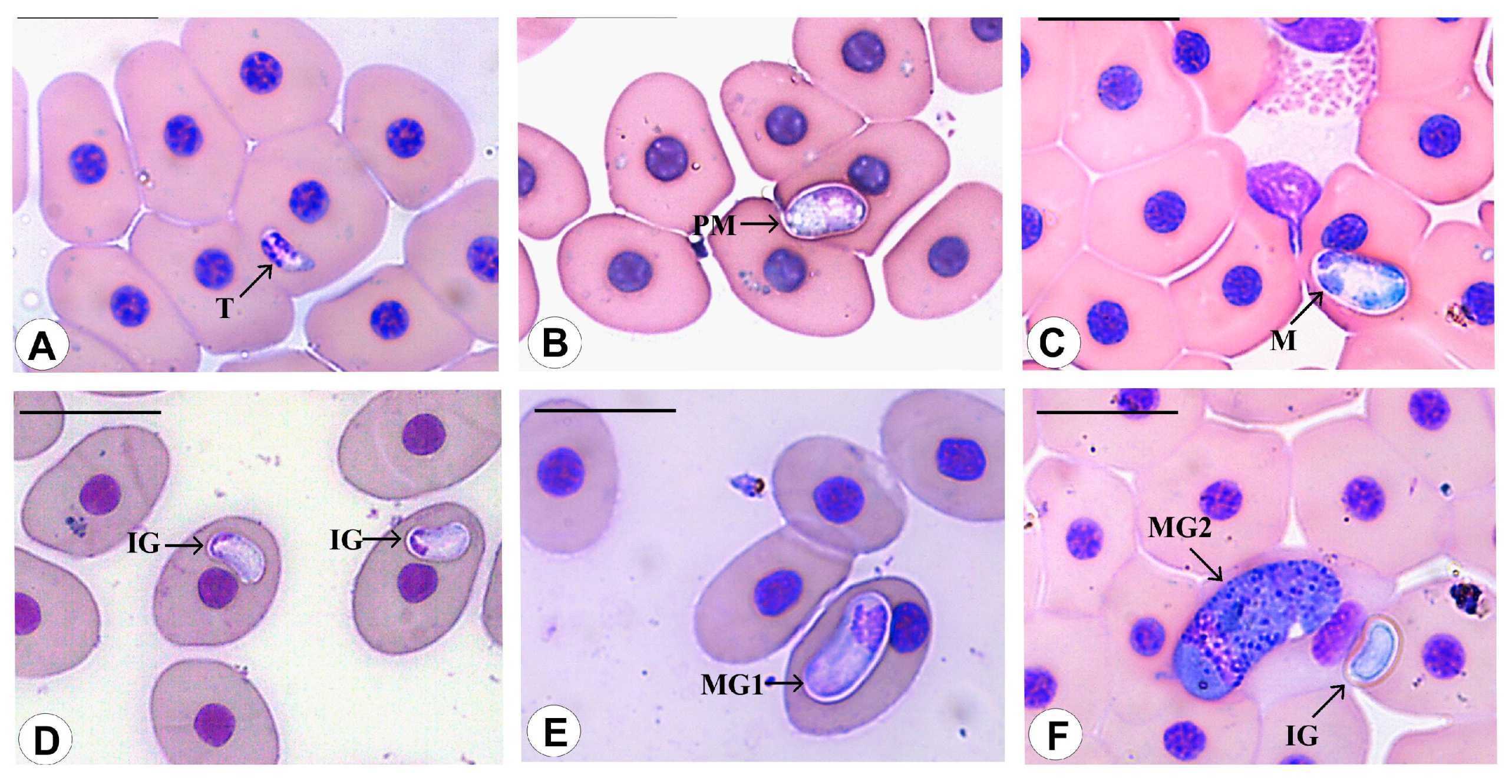
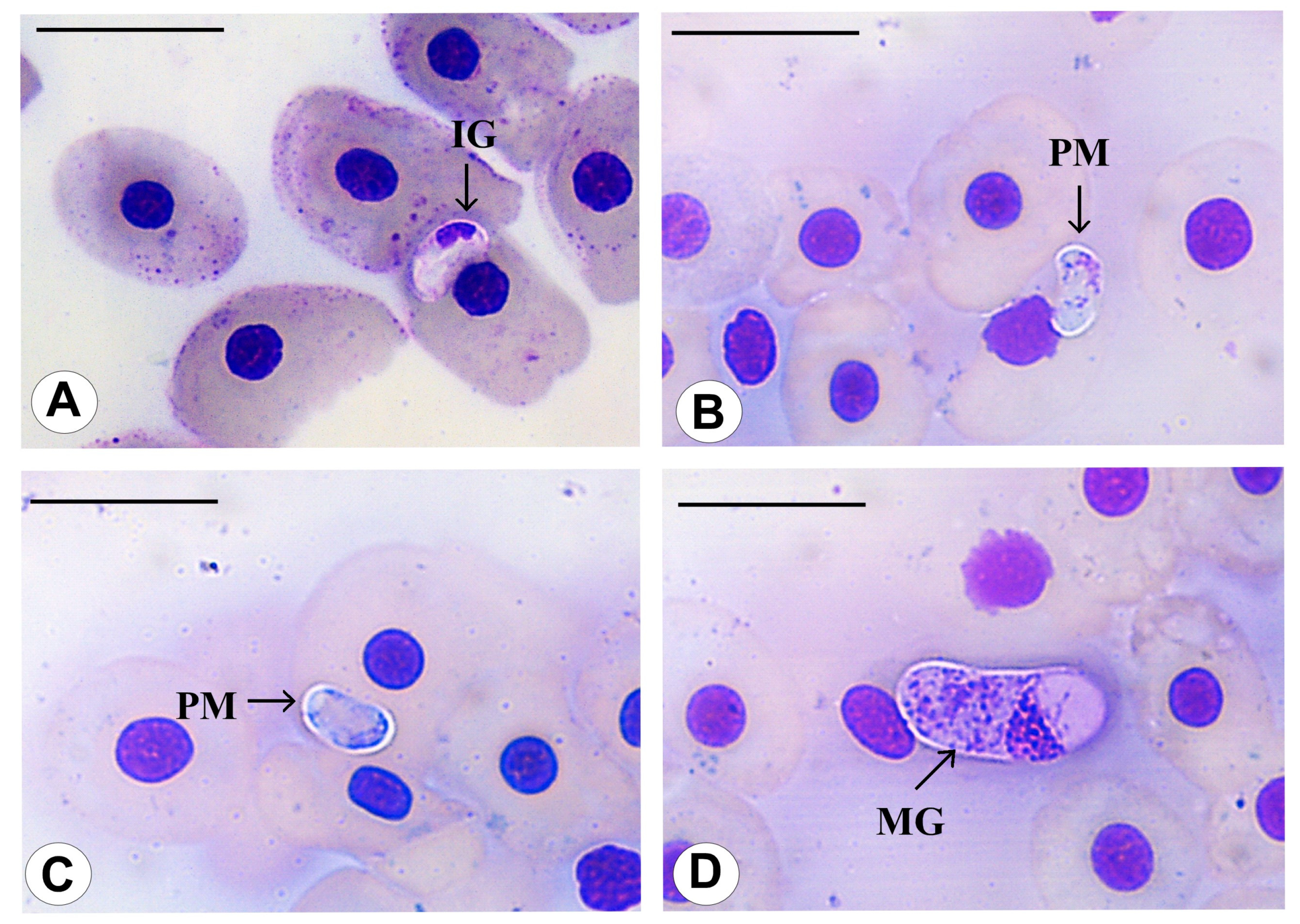
3.3. Morphological Data
3.4. Species Descriptions
3.4.1. Haemogregarina rubra n. sp. Úngari, Silva and O’Dwyer 2023

3.4.2. Haemogregarina araguaiensis n. sp. Úngari, Silva and O’Dwyer 2023

3.4.3. Haemogregarina goianensis n. sp. Úngari, Silva and O’Dwyer 2023

3.4.4. Haemogregarina unifila n. sp. Úngari, Silva and O’Dwyer 2023

3.4.5. Haemogregarina brasiliana n. sp. Úngari, Silva and O’Dwyer 2023

3.4.6. Haemogregarina tigrina n. sp. Úngari, Silva and O’Dwyer 2023

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thrall, M.A.; Baker, D.C.; Campbell, T.W.; De Nicola, D.; Fettman, M.J.; Lassend, E.D.; Rebar, A.; Weiser, G. Hematologia e Bioquímica Clínica e Veterinária; Roca: São Paulo, Brazil, 2007. [Google Scholar]
- Siřoký, P.; Kamler, M.; Modrý, D. Long-term occurrence of Hemolivia cf. mauritanica (Apicomplexa: Adeleina: Haemogregarinidae) in the marginated tortoise Testudo marginata (Reptilia: Testudinae): Evidence from experimental infection. J. Parasitol. 2004, 54, 13–18. [Google Scholar]
- Harris, D.J.; Graciá, E.; Jorge, F.; Maia, J.P.M.C.; Pereira, A.; Carretero, M.A.; Giménez, A. Molecular detection of Hemolivia (Apicomplexa: Haemogregarinidae) from ticks of North African Testudo graeca (Testudines: Testudinidae) and an estimation of their phylogenetic relationship using 18S rRNA sequences. Comp. Patasitol. 2013, 80, 292–296. [Google Scholar] [CrossRef]
- Kviceřová, J.; Hypša, V.; Dvořáková, N.; Mikuliček, P.; Jandizik, D.; Gardner, G.; Javanbakht, H.; Tiar, G.; Široký, P. Hemolivia and Hepatozoon: Heaemogregarines with tangled evolutionary relationships. Protist 2014, 165, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.A.; Lawton, S.P.; Davies, A.J.; Smit, N.J. Reassignment oif the land tortoise haemogregarine Haemogreagrina fitzsimonsi Dias 1953 (Adeleorina: Haemogregarinidae) to the genus Hepatozoon Miller 1908 (Adeleorina: Hepatozoidae) based on parasites morphology, life cycle and ohylogenetic analysis of 18S rDNA sequence fragments. Parasitology 2014, 141, 1611–1620. [Google Scholar]
- Gutiérrez-Liberato, G.A.; Lotta-Arévalo, I.A.; Rodrígues-Almonacid, C.C.; Vargas-Ramírez, M. Molecular and morphological description of the first Hepatozoon (Apicomplexa: Hepatozoidae) species infecting a neotropical turtle, with an approach to its phylogenetic relationships. Parasitology 2021, 148, 747–759. [Google Scholar] [CrossRef]
- Jakes, K.A.; O’Donoghue, P.; Munro, M.; Adlard, R. Hemoprotozoa of freshwater turtles in Queensland. J. Wildl. Dis. 2001, 37, 12–19. [Google Scholar] [CrossRef]
- Telford, S.R., Jr. Hemoparasite of the Reptilian: Color Atlas and Text; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Davis, A.K.; Sterrett, S. Prevalence of haemogregarine parasites in three freshwater turtle species in a population in northeast Georgia, USA. Int. J. Zool. Res. 2011, 7, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Pineda-Catalan, O.; Perkins, S.L.; Peirce, M.A.; Esgstrand, R.; Garcia-Davila, C.; Pinedo-Vasquez, M.; Aguirre, A.A. Revision of Hemoproeid genera and description and redescription of two species of chelonian hemoproteid parasites. J. Parasitol. 2013, 99, 1089–1098. [Google Scholar] [CrossRef]
- Rossow, J.A.; Hernandez, S.M.; Summer, S.M.; Altman, B.R.; Crider, C.G.; Gammage, M.B. Haemogregarine infections of three species of aquatic freshwater turtles from two sites in Costa Rica. Int. J. Parasitol. Parasites Wildl. 2013, 2, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Devořálová, N.; Kviceřová, J.; Ppapousek, I.; Javany-Bakht, H.; Tiar, G.; Kami, H.; Siřoký, P. Haemomogregarines from western Paleartic fresh turtles (genera Emys, Mauremys) are coespecific with Haemogregarina stepanowi Denilewsky, 1885. Parasitology 2013, 141, 522–530. [Google Scholar]
- Devořálová, N.; Kviceřová, J.; Hostovský, M.; Siřoký, P. Haemogregarines of freshwater turtles from Southeast Asia with a description of Haemogregarina sacaliae sp. n. and a redescription of Haemogregarina pellegrini Laveran and Pettit, 1910. Parasitology 2015, 142, 816–826. [Google Scholar]
- Picelli, A.M.; Carvalho, A.V.; Viana, L.A.; Malvasio, A. Prevalence and parasitemia of Haemogregarina sp. in Podocnemis expansa (Testudines: Podocnemididae) form the Brazilian Amazon. Braz. J. Vet. Parasitol. 2015, 42, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arizza, V.; Sacco, D.; Russo, D.; Scardino, R.; Arculeo, M.; Vamberger, M.; Marrone, F. The good, the bad and the ugly: Emys trinacris, Placobdella costata and Haemogregarina stepanowi in Sicily (Testudines, Annelida and Apicomplexa). Folia Parasitol. 2016, 63, 29. [Google Scholar] [CrossRef]
- Úngari, L.P.; Santos, A.L.Q.; O’Dwyer, L.H.; Da Silva, M.R.L.; Fava, N.N.M.; Paiva, G.C.M.; Pinto, R.M.C.; Cury, M.C. Haemogregarina podocnemis sp. nov.: Description of a new species of Haemogregarina Danilewsky 1885 (Adeleina: Haemogregarinidae) in free-living and captive yellow-spotted river turtles Podocnemis unifilis (Testudines: Podocnemididae) from Brazil. Parasitol. Res. 2018, 117, 1535–1548. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.P.; André, M.R.; Alves Júnior, J.R.F.; Lustosa, A.P.G.; Werther, K. Molecular detection of hemogregarines and haemosporidians in Brazilian free-living testudines. Int. J. Parasitol. Parasites Wildl. 2018, 7, 75–84. [Google Scholar] [CrossRef]
- Correa, J.K.C.; Picelli, A.M.; da Silva, M.R.L.; Valadão, R.M.; Hernández-Ruz, E.J.; Viana, L.A. Phylogenetic analysis of chelonian hemogregarines reveals shared species among the Amazonian freshwater turtle Podocnemis spp. and provides a description of two new species of Haemogregarina. Parasitol. Res. 2022, 121, 691–701. [Google Scholar] [CrossRef]
- Levine, N.D. Progress in taxonomy of the Apicomplexan protozoa. J. Protozool. 1988, 35, 518–520. [Google Scholar] [CrossRef]
- Siddall, M.E.; Desser, S.S. Ultrastructure of merogonic development of Haemogregarina (sensu lato) myoxocephali (Apicomplexa: Adeleina) in the marine leech Malmiana scorpii and localization of infective stages in the salivary cells. Eur. J. Protistol. 1993, 29, 191–201. [Google Scholar] [CrossRef]
- Siddall, M.E.; Desser, S.S. Transmission of Haemogregarina balli from painted turtles to snapping turtles through the leech Placobdella ornata. J. Parasitol. 2001, 87, 1217–1218. [Google Scholar] [CrossRef]
- Siddall, M.E. Phylogeny of adeleid blood parasites with a partial systematic revision of the haemogregarine complex. J. Eukaryot. Microbiol. 1995, 42, 116–125. [Google Scholar] [CrossRef]
- Davies, A.J.; Smit, N.J.; Hayes, P.M.; Seddon, A.M.; Wertheim, D. Haemogregarina bigemina (Protozoa: Apicomplexa: Adeleorina)-past, present and future. Folia Parasitol. 2004, 51, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Siddall, M.E.; Desser, S.S. Merogonic development of Haemogregarina balli (Apicomplexa: Adeleina: Haemogregarinidae) in the leech Placobdella ornata (Glossiphoniidae), its transmission to a chelonian intermediate host and phylogenetic implications. J. Parasitol. 1991, 77, 426–436. [Google Scholar] [CrossRef]
- Siddall, M.E.; Desser, S.S. Prevalence and intensity of Haemogregarina balli (Apicomplexa: Adeleina: Haemogregarinidae) in three turtles species from Ontaro with observations on intraerythrocytic development. Canad. J. Zool. 1992, 70, 123–128. [Google Scholar] [CrossRef]
- Davies, A.J.; Johnston, M.R. The biology of someintraerytrocytic parasites of fishes, amphibian and reptiles. Adv. Parasitol. 2000, 45, 1–107. [Google Scholar] [PubMed]
- Soares, P.; Brito, E.S.; Paiva, F.; Pavan, D.; Viana, L.A. Haemogregarina spp. in wild population from Podocnemis unifilis Trschel, 1848 in Brazilian Amazonia. Parasitol. Res. 2014, 113, 4499–4503. [Google Scholar] [CrossRef]
- Costa, H.C.; Bérnils, R.S. Répteis do Brasil e Suas Unidades Federativas: Lista de Espécies. Soiedade Brasileira de Herpetologia. 2018. Available online: http://www.sbherpetologia.org.br (accessed on 23 August 2021).
- Campos-Brites, V.L.; Ratin, F.T. The influence of agricultural and urban contamination on leech infestation of freshwater turtles, Phrynops geoffroanus, taken from two areas of the Uberabinha river. Environ. Monit. Assess. 2004, 96, 273–281. [Google Scholar] [CrossRef]
- Pessoa, L.M.B.; Scalon, M.C.; Carneiro, F.T.; Lota, M.G.M.; Silva, T.F.; Monteiro, R.V.; Paludo, G.R. Ocorrência de hemogregarina em duas espécies de cágados brasileiros. Acta Sci. Vet. 2016, 44, 1381. [Google Scholar]
- Carvalho, A.V.; Da Silva, A.C.L.; Pacheco, V.R. Relação entre parasitemia de Sauroplasma sp. (Piroplasmorida: Haemohormidiidae) e índices leucocitários em Podocnemis expansa (tartatuga da amazônia). RICA 2019, 10, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Goes, V.C.; Brito, E.S.; Valadão, R.M.; Gutierrez, C.O.; Picelli, A.M.; Viana, L.A. Haemogregarine (Apicomplexa: Adeleorina) infection in Vanderhaege’s toad-headed turtle, Mesoclemmys vanderhaegei (Chelidae), from a Brazilian Neotropical savanna region. Folia Parasitol. 2018, 65, 12. [Google Scholar] [CrossRef]
- Portelinha, T.C.G.; Malvasio, A.; Piña, C.I.; Bertoluci, J.A. Reprodutive Allometry of Podocnemis expansa (Testudines: Podocnemididae) in Southern Brazilian Amazon. J. Herpetol. 2013, 47, 232–236. [Google Scholar] [CrossRef]
- Malvasio, A.; Souza, A.M.; Gomes, N.; Sampaio, F.A.A.; Molina, F.B. Variações ontogenéticas na morfometria e morfologia do canal alimentar pós-faríngeo de Trachemys dorbignyi (Duméril & Bibron, 1835), (Schweigger, 1812), (Cornalia, 1849) (Anapsida; Testudines). Podocnemis expansa, P. unifilis (Troschel, 1848) e P. sexturberculata. Publ. Avulsas Inst. Pau. Bras. His. Nat. 2002, 5, 39–51. [Google Scholar]
- Araújo, F.A.A.; Santa Lúcia, M.; Cabral, R.F. Epidemiologia dos acidentes por animais peçonhentos. In Animais Peçonhentos no Brasil: Biologia, Clínica e Terapêutica dos Acidentes; Cardoso, J.L., França, F., Wen, F., Malaque, C., Junior, V.H., Eds.; Sarvier: São Paulo, Brazil, 2003; pp. 6–12. [Google Scholar]
- Zippel, K.C.; Lillywhite, H.B.; Mlandnich, C.R.J. New vascular system in reptiles: Anatomy and postural hemodynamics of vertebral venomous lexus in snakes. J. Morphol. 2001, 250, 179–184. [Google Scholar] [CrossRef]
- Eisen, R.J.; Schall, J.J. Life history of malaria parasite (Plasmodium mexicanum): Independent traits and basis for variation. Proc. R. Soc. B Biol. Sci. 2000, 267, 793–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, C.A.; Smit, N.J.; Davies, A.J. A redescription of Haemogregarina fitzsimonsi Dias, 1953 and some comments on Haemogregarina parvula Dias, 1953 (Adeleorina: Haemogregarinidae) from Southern African tortoises (Cyptodira: Testudinidae) with new host data and distribution records. Folia Parasitol. 2009, 56, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Sebben, A. Microdissecação fisiológica a fresco: Uma nova visão sobre a anatomia de anfíbios e répteis. In Herpetologia No Brasil II, 1st ed.; Nascimento, L.B., Oliveira, M.E., Eds.; Sociedade Brasileira de Herpetologia: Belo Horizonte, Brazil, 2007; pp. 301–325. [Google Scholar]
- Ujvari, B.; Madsen, T.; Olsson, M. High prevalence of Hepatozoon spp. (Apicomplexa: Hepatozoidae) infection in Water Phythons (Liasis fuscus) from Tropical Australia. J. Parasitol. 2004, 90, 670–672. [Google Scholar] [CrossRef] [Green Version]
- Perkins, S.L.; Keller, A.K. Phylogeny of nuclear small subunit rRNA genes of hemogregarines amplified with specific oligonucleotideos. J. Parasitol. 2001, 87, 870–876. [Google Scholar] [CrossRef]
- O’Dwyer, L.H.; Moço, T.C.; Paduan, K.S.; Spenassatto, C.; Silva, R.J.; Ribolla, P.E.M. Description of three new species of Hepatozoon (Apicomplexa: Hepatozoidae), from Rattlesnakes (Crotallus durrissus terrificus) based on molecular, morphometric and morphologic characters. Exp. Parasitol. 2013, 135, 200–207. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Sones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14–14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tavaré, S.; Miura, R.M. Some probabilistic and statistical problems in the analysis of DNA sequences, Lectures on mathematics in the life sciences. Lect. Math. Life Sci. 1986, 17, 57–86. [Google Scholar]
- Rambaut, A. FigTree v1.4.2. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 11 March 2022).
- Vogt, R.C.; Fagundes, C.K.; Bataus, Y.S.L.; Balestra, R.A.M.; Batista, F.R.W.; Uhlig, V.M.; Silveira, A.L.; Bager, A.; Batistella, A.M.; Souza, F.L.; et al. Avaliação do Risco de Extinção de Podocnemis unifilis Troschel, 1848 No Brasil. Processo de Avaliação do Risco de Extinção da Fauna Brasileira. 2015. ICMBio. Available online: http://www.icmbio.gov.br/portal/biodiversidade/fauna-brasileira/estado-de-conservacao/7426-repteis-podocnemis-unifilis-tracaja.html (accessed on 9 July 2022).
- Ferrara, C.R.; Fagundes, C.K.; Morcatty, T.Q.; Vogt, R.C. Quelônios Amazônicos: Guia de Identificação e Distribuição; Wildlife Conservation Society: Manaus, Brazil, 2017. [Google Scholar]
- Vanzolini, P.E. On the eggs of Brasilian Podocnemis (Testudines Podocnemididae). Biol. Geral. Exp. 2001, 2, 3–17. [Google Scholar]
- Pritchard, P.C.H.; Trebbau, P. The Turtles of Venezuela; SSAR: New York, NY, USA, 1984; 403p. [Google Scholar]
- Selera, G., Jr.; Malvasio, A.; Portelinha, T.C.G. Avaliação da predação de Podocnemis expansa e Podocneis unilifis (Testudines, Podocnemidae) no rio Javaés, Tocantins. Acta Amaz. 2009, 39, 207–214. [Google Scholar] [CrossRef]
- Streit, B.; Peter, H.M. Long-term effects on atrazine to selected freshwater invertebrates. Arch. Hydrobiol. Suppl. 1978, 55, 62–77. [Google Scholar]
- Pearse, D.E.; Arndt, A.D.; Valenzuela, N.; Miller, B.A.; Cantarelli, V.; Sites, J.W., Jr. Estimating population structure under nonequilibrium conditions in a conservation context: Continent-wide population genetics of the giant Amazon river turtle, Podocnemis expansa (Chelonia; Podocnemididae). Mol. Ecol. 2006, 15, 985–1006. [Google Scholar] [CrossRef]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Petry, P. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Albert, J.S.; Reis, R. Historical Biogeography of Neotropical Freshwater Fshes; University of California Press: Berkeley, CA, USA, 2001. [Google Scholar]
- Roberto, I.J.; Bittencourt, P.S.; Muniz, F.L.; Hernández-Rangel, S.M.; Nóbrega, Y.C.; Ávila, R.W.; Farias, I.P. Unexpected but unsurprising lineage diversity within the most widespread Neotropical crocodilian genus Caiman (Crocodylia, Alligatoridae). Syst. Biodivers. 2020, 18, 377–395. [Google Scholar] [CrossRef]
- Escalona, T.; Engstrom, T.N.; Hernandez, O.E.; Bock, B.C.; Vogt, R.C.; Valenezuela, N. Population genetics of the endangered South American freshwater turtle, Podocnemis uniflis, inferred from microsatellite DNA data. Conserv. Genet. 2009, 10, 1683–1696. [Google Scholar] [CrossRef]
- Páez, V.P.; Morales-Betancourt, M.A.; Lasso, C.A.; Castaño, M.O.V.; Bock, B.C. Biología y conservación de las tortugas continentales de Colombia. In Serie Recursos Hidrobiológicos y Pesqueros Continentales de Colombia; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2012. [Google Scholar]
- Fermino, B.R.; Viola, L.B.; Paiva, F.; Garcia, H.A.; de Paula, C.D.; Botero-Arias, R.; Takata, C.S.A.; Campaner, M.; Hamilton, P.B.; Camargo, E.P.; et al. The phylogeography of trypanosomes from South American alligatorids and African crocodilids is consistent with the geological history of South American river basins and the transoceanic dispersal of Crocodylus at the Miocene. Parasit. Vectors 2013, 6, 313 . [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Municipality | Year | State * | Coordinates |
|---|---|---|---|---|
| São Sebastião do Paraíso farm | Boa Esperança do Sul | 2019 | SP | 21°51′36.83″ S, 48°27′3.56″ W |
| Vermelho river | Britânia | 2017 | GO | 15°10′44.82″ S, 51°9′58.48″ W |
| Araguaia river | Itacaiú | 2019 | GO | 15°04′39.81″ S, 51°26′68,91″ W |
| Stream (unnamed) | Nova Xavantina | 2018/2020 | MT | 14°31′48.03″ S, 51°41′43.88″ W |
| Sertaneja Retiro farm | Cocalinho | 2018/2020 | MT | 14°26′19.98″ S, 51°35′20.17″ W |
| Pindaíba river | Araguaiana | 2018/2020 | MT | 14°35′47″ S, 51°43′9.59″ W |
| Testudines: Pelurodira | Family | Year | State | N | P |
|---|---|---|---|---|---|
| Chelonoidis carbonarius (Spix, 1824) | Testudinidae | 2018/2020 | MT | 4 | 0 |
| Phrynops geoffroanus (Schweigger, 1812) | Chelidae | 2019 | GO, SP | 2 | 0 |
| Podocnemis expansa (Schweigger, 1812) | Podocnemididae | 2017/2019 | GO | 3 | 3 |
| Podocnemis unifilis (Troschel, 1848) | Podocnemididae | 2017/2019 | GO, MT | 31 | 31 |
| Haemogregarina Isolates | 1. | 2. | 3. | 4. | 5. | 6. | 7. | 8. | 9. | 10. |
|---|---|---|---|---|---|---|---|---|---|---|
| 1. OQ377557 - H. rubra n. sp. | 98.1 | 98.9 | ------------- | 98.1 | 98.1 | 98.3 | 97.2 | 98.5 | 98.3 | |
| 2. OQ388265 - H. araguaiensis n. sp. | 96.33 | 97.7 | ------------- | 97.5 | 97.5 | 97.8 | 96.9 | 97.3 | 97.9 | |
| 3. OQ377555 - H. goianensis n. sp. | 95.95 | 99.61 | ------------- | 97.5 | 97.8 | 97.8 | 96.9 | 97.1 | 97.4 | |
| 4. OQ377710 - H. unifila n. sp. | 93.82 | 96.52 | 96.32 | ------------- | ------------- | ------------- | ------------- | ------------- | ------------- | |
| 5. OQ377129 - H. brasiliana n. sp. | 94.02 | 96.91 | 96.71 | 98.84 | 99.7 | 98.6 | 96.9 | 99.3 | 99.4 | |
| 6. OQ377558 - H. tigrina n. sp. | 94.40 | 97.29 | 97.10 | 98.45 | 99.61 | 98.5 | 97.0 | 99.7 | 99.7 | |
| 7. OQ377547 - H. embaubali (genotype 4) | 93.24 | 95.55 | 95.36 | 98.84 | 98.45 | 98.26 | 97.2 | 98.5 | 98.6 | |
| 8. OQ377462 - H. embaubali (genotype 3) | 92.86 | 95.55 | 95.36 | 99.03 | 98.26 | 97.87 | 99.03 | 96.7 | 97.0 | |
| 9. OQ377458 - H. embaubali (genotype 2) | 93.44 | 96.13 | 95.94 | 99.61 | 98.84 | 98.45 | 99.23 | 99.42 | 99.81 | |
| 10. OQ377457 - H. embaubali (genotype 1) | 93.24 | 95.94 | 95.74 | 99.42 | 98.65 | 98.26 | 99.42 | 99.61 | 99.81 | |
| 11. MW540607—H. embaubali | 93.24 | 95.94 | 95.74 | 99.42 | 98.65 | 98.26 | 99.42 | 99.61 | 95.81 | 100 |
| 12. MW540605—H. karaja | 95.95 | 97.49 | 97.10 | 95.94 | 95.16 | 95.55 | 95.16 | 95.16 | 95.55 | 95.55 |
| 13. MF476203—H. podocnemis | 92.47 | 95.16 | 95.35 | 98.26 | 97.49 | 97.29 | 98.26 | 98.45 | 98.65 | 98.84 |
| 14. HQ224959—H. balli | 90.15 | 92.26 | 92.46 | 92.84 | 92.07 | 91.88 | 92.26 | 92.46 | 92.84 | 92.84 |
| 15. KM887507—H. sacaliae | 89.77 | 92.26 | 92.46 | 93.23 | 92.46 | 92.26 | 92.65 | 92.84 | 93.23 | 93.23 |
| 16. KF257928—H. stepanowi | 90.35 | 92.07 | 92.26 | 92.65 | 91.88 | 91.68 | 92.07 | 92.26 | 92.65 | 92.65 |
| 17. KM887509—H. pellegrini | 90.35 | 92.28 | 92.47 | 92.86 | 92.08 | 91.89 | 92.28 | 92.47 | 92.86 | 92.86 |
| Haemogregarina | DS | N | P (%) | C | PL | PW | PA | NL | NW | NA |
|---|---|---|---|---|---|---|---|---|---|---|
| H. rubra n. sp. | T | 4 | 0.99 | 7.26 ± 0.34 | 3.50 ± 0.19 | 19.38 ± 3.44 | 2.63 ± 0.81 | 3.50 ± 0.19 | ||
| IG | 5 | 11.3 ± 0.12 | 5.95 ± 0.37 | 55.0 ± 1.29 | 3.44 ± 0.14 | 3.99 ± 0.25 | ||||
| PM | 2 | 11.7 ± 0.08 | 5.31 ± 0.44 | 56.2 ± 2.02 | ||||||
| MG1 | 5 | 16.98 ± 0.92 | 7.55 ± 0.79 | 113.79 ± 9.47 | 4.26 ± 0.60 | |||||
| MG2 | 8 | 20.08 ± 0.61 | 9.56 ± 0.76 | 164.27 ± 6.33 | 6.13 ± 1.08 | 7.82 ± 1.69 | ||||
| H. araguaiensis n. sp. | T | 7 | 1.4 and 2.14 | 10.53 ± 1.66 | 3.40 ± 0.28 | 28.66 ± 1.30 | 3.06 ± 0.04 | 2.77 ± 0.27 | 5.82 ± 0.68 | |
| PM | 5 | 12.57 ± 0.45 | 3.75 ± 0.78 | 30.26 ± 2.08 | 3.12 ± 0.97 | 2.44 ± 1.87 | 4.89 ± 1.99 | |||
| IG | 15 | 10.72 ± 0.41 | 5.42 ± 0.82 | 42.82 ± 2.78 | 2.67 ± 0.62 | 1.20 ± 0.19 | 4,44 ± 0.53 | |||
| MG1 | 20 | 13.65 ± 1.04 | 6.11 ± 0.26 | 62.69 ± 5.14 | 4.76 ± 1.47 | 4.35 ± 0.99 | 10.30 ± 1.89 | |||
| MG2 | 20 | 21.01 ± 4.99 | 11.50 ± 2.78 | 180.0 ± 21.08 | 6.99 ± 2.65 | 7.76 ± 2.56 | 30.05 ± 11.02 | |||
| H. goianensis n. sp. | T | 5 | 0.6 and 5.0 | 8.49 ± 0.62 | 3.84 ± 0.63 | 25.52 ± 3.70 | 3.51 ± 0.67 | 3.50 ± 0.19 | 9.82 ± 2.17 | |
| PM | 5 | 11.93 ± 1.42 | 7.08 ± 1.35 | 65.62 ± 18.14 | 7.09 ± 0.59 | 6.90 ± 0.91 | 38.8 ± 4.27 | |||
| IG | 15 | 11.05 ± 2.64 | 4.58 ± 1.15 | 40.99 ± 9.23 | ||||||
| MG1 | 20 | 11.57 ± 0.21 | 5.41 ± 0.34 | 56.24 ± 0.78 | 4.48 ± 0.12 | 0.80 ± 0.02 | 7.99 ± 2.01 | |||
| MG2 | 25 | 21.96 ± 2.07 | 9.28 ± 1.92 | 178.48 ± 40.10 | 7.85 ± 1.41 | 7.78 ± 1.03 | 30.77 ± 9.01 | |||
| H. unifila n. sp. | T | 5 | 1.0–6.42 | 7.03 ± 0.48 | 3.05 ± 0.33 | 16.04 ± 2.20 | 4.66 ± 0.08 | 2.57 ± 0.01 | 9.43 ± 0.96 | |
| IG | 15 | 7.99 ± 2.58 | 3.79 ± 1.44 | 26.74 ± 13.72 | 4.35 ± 1.01 | 3.95 ± 1.33 | 13.95 ± 5.52 | |||
| MG1 MG2 PM | 15 15 3 | 13.58 ± 0.06 19.43 ± 4.84 10.55 ± 0.82 | 6.13 ± 0.06 9.28± 1.46 6.24 ± 1.99 | 66.35 ± 2.05 150.65 ± 29.24 61.74 ± 4.78 | 2.63 ± 1.37 5.76 ± 0.35 | 3.47 ± 0.28 7.35 ± 0.59 | 9.70 ± 0.12 39.39 ± 11.50 | |||
| M | 2 | 14.67 ± 0.78 | 8.25 ± 1.43 | 91.7 ± 3.24 | 4.46 ± 0.15 | 3.79 ± 0.06 | 11.04 ± 1.40 | |||
| H. brasiliana n. sp. | PM | 6 | 0.20 and 2.5 | 13.83 ± 2.22 | 6.25 ± 0.97 | 65.41 ± 10.12 | ||||
| IG | 15 | 12.87 ± 1.33 | 6.54 ± 1.76 | 62.78 ± 8.04 | 3.27 ± 1.17 | 1.06 ± 0.88 | 4.11 ± 0.58 | |||
| MG | 10 | 21.38 ± 1.48 | 9.58 ± 0.77 | 168.16 ± 19.55 | 5.51 ± 0.52 | 7.14 ± 0.49 | 28.45 ± 1.61 | |||
| H. tigrina n. sp. | Me | 5 | 3.9 and 5.0 | 6.55 ± 0.21 | 2.10 ± 0.33 | 14.07 ± 1.38 | 4.47 ± 1.30 | 1.99 ± 0.28 | 10.04 ± 1.85 | |
| T | 10 | 7.41 ± 0.78 | 3.03 ± 0.46 | 18.86 ± 2.13 | 4.28 ± 0.81 | 3.86 ± 0.47 | 13.12 ± 1.22 | |||
| PM | 5 | 10.45 ± 2.92 | 5.12 ± 1.39 | 45.14 ± 15.07 | ||||||
| M | 3 | 21.57 ± 2.01 | 9.88 ± 1.78 | 174.32 ± 21.06 | ||||||
| IG | 25 | 11.02 ± 0.34 | 5.45 ± 1.40 | 45.14 ± 3.98 | 2.33 ± 0.88 | 0.98 ± 0.01 | 4.03 ± 0.87 | |||
| MG | 15 | 25,37 ± 1.43 | 12.23 ± 0.78 | 230.01 ± 12.85 | 6.02 ± 0.34 | 9.40 ± 0.97 | 53.36 ± 7.35 | |||
| H. embaubali (Genotype 1) | T | 15 | 0.01–12.49 | 7.39 ± 1.44 | 3.24 ± 0.79 | 19.02 ± 1.64 | 3.80 ± 0.61 | 2.89 ± 0.31 | 6.10 ± 1.22 | |
| GV | 15 | 11.1 ± 0.78 | 4.3 ± 0.88 | 57.80 ± 5.01 | 5.0 ± 0.74 | 3.22 ± 0.95 | 11.08 ± 3.41 | |||
| PM | 15 | 10.27 ± 0.60 | 5.18 ± 0.49 | 53.13 ± 6.01 | ||||||
| M | 5 | 12.50 ± 0.26 | 7.08 ± 0.46 | 85.27 ± 6.08 | 2.27 ± 0.25 | 1.00 ± 0.17 | 4.05 ± 0.55 | |||
| IG | 25 | 10.30 ± 0.65 | 4.59 ± 0.73 | 50.84 ± 4.00 | 2.13 ± 0.21 | 2.98 ± 0.47 | 5.17 ± 1.33 | |||
| MG1 | 25 | 12.90 ± 1.27 | 6.85 ± 2.20 | 81.17 ± 3.38 | 3.19 ± 0.88 | 1.01 ± 0.12 | 5.06 ± 1.11 | |||
| MG2 | 30 | 20.10 ± 1.62 | 9.59 ± 0.83 | 134.82 ± 15.00 | 5.94 ± 1.89 | 7.26 ± 2.16 | 33.59 ± 4.97 | |||
| Me | 10 | 3.75 ± 0.25 | 20.10 ± 1.62 | 13.21 ± 3.51 | 152.46 ± 77.36 | |||||
| H. embaubali (Genotype 2) | GV GM PM | 20 10 10 | 0.45–7.65 | 10.06 ± 0.79 17.32 ± 1.14 11.75 ± 1.31 | 5.80 ± 1.01 7.88 ± 0.90 6.33 ± 0.87 | 50.84 ± 4.45 119.22 ± 3.56 55.60 ± 2.64 | ||||
| H. embaubali (Genotype 3) | T | 15 | 13.97 | 5.88 ± 0.97 | 2.21 ± 0.43 | 16.46 ± 2.81 | 3.12 ± 0.39 | 2.18 ± 0.26 | ||
| GV | 15 | 8.13 ± 1.41 | 3.99 ± 0.41 | 30.3 ± 2.47 | ||||||
| IG | 15 | 7.98 ± 1.21 | 4.27 ± 0.54 | 33.72 ± 3.69 | 4.15 ± 0.15 | 2.12 ± 0.36 | ||||
| MG | 20 | 10.42 ± 1.07 | 5.46 ± 0.70 | 49.12 ± 4.45 | 4.27 ± 0.20 | 2.19 ± 0.41 | ||||
| Me | 10 | 3.03 ± 0.32 | 15.57 ± 2.51 | 12.66 ± 1.70 | 133.61 ± 22.99 | |||||
| H. embaubali (Genotype 4) | IG | 10 | 1.25 | 14.78 ± 0.21 | 7.00 ± 0.15 | 86.06 ± 1.47 | 3.66 ± 0.52 | 3.43 ± 0.22 | 9.11 ± 1.78 | |
| MG | 5 | 10.48 ± 3.22 | 5.56 ± 1.71 | 52.98 ± 16.81 | 3.85 ± 0.65 | 3.69 ± 0.45 | 10.82 ± 1.72 | |||
| PM | 4 | 12.08 ± 0.96 | 6.29 ± 0.44 | 62.30 ± 6.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Úngari, L.P.; Santos, A.L.Q.; da Silva, R.J.; O’Dwyer, L.H. Hemogregarine Diversity Infecting Brazilian Turtles with a Description of Six New Species of Haemogregarina (Apicomplexa: Adeleorina: Haemogregarinidae). Diversity 2023, 15, 296. https://doi.org/10.3390/d15020296
Úngari LP, Santos ALQ, da Silva RJ, O’Dwyer LH. Hemogregarine Diversity Infecting Brazilian Turtles with a Description of Six New Species of Haemogregarina (Apicomplexa: Adeleorina: Haemogregarinidae). Diversity. 2023; 15(2):296. https://doi.org/10.3390/d15020296
Chicago/Turabian StyleÚngari, Letícia Pereira, André Luiz Quagliatto Santos, Reinaldo José da Silva, and Lucia Helena O’Dwyer. 2023. "Hemogregarine Diversity Infecting Brazilian Turtles with a Description of Six New Species of Haemogregarina (Apicomplexa: Adeleorina: Haemogregarinidae)" Diversity 15, no. 2: 296. https://doi.org/10.3390/d15020296