
A New Rhynchosaur Taxon from the Popo Agie Formation, WY: Implications for a Northern Pangean Early-Late Triassic (Carnian) Fauna
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
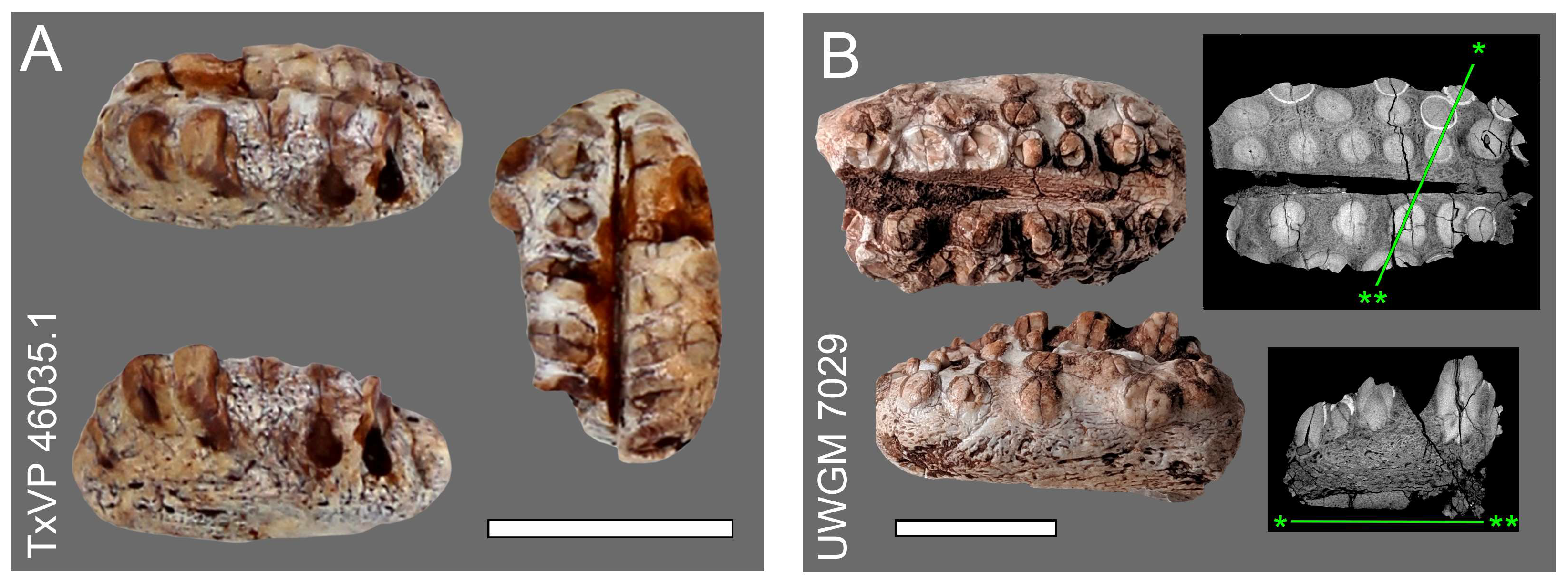
2.1. Fossil Material and Imaging
2.2. Naming/Nomenclature Statement
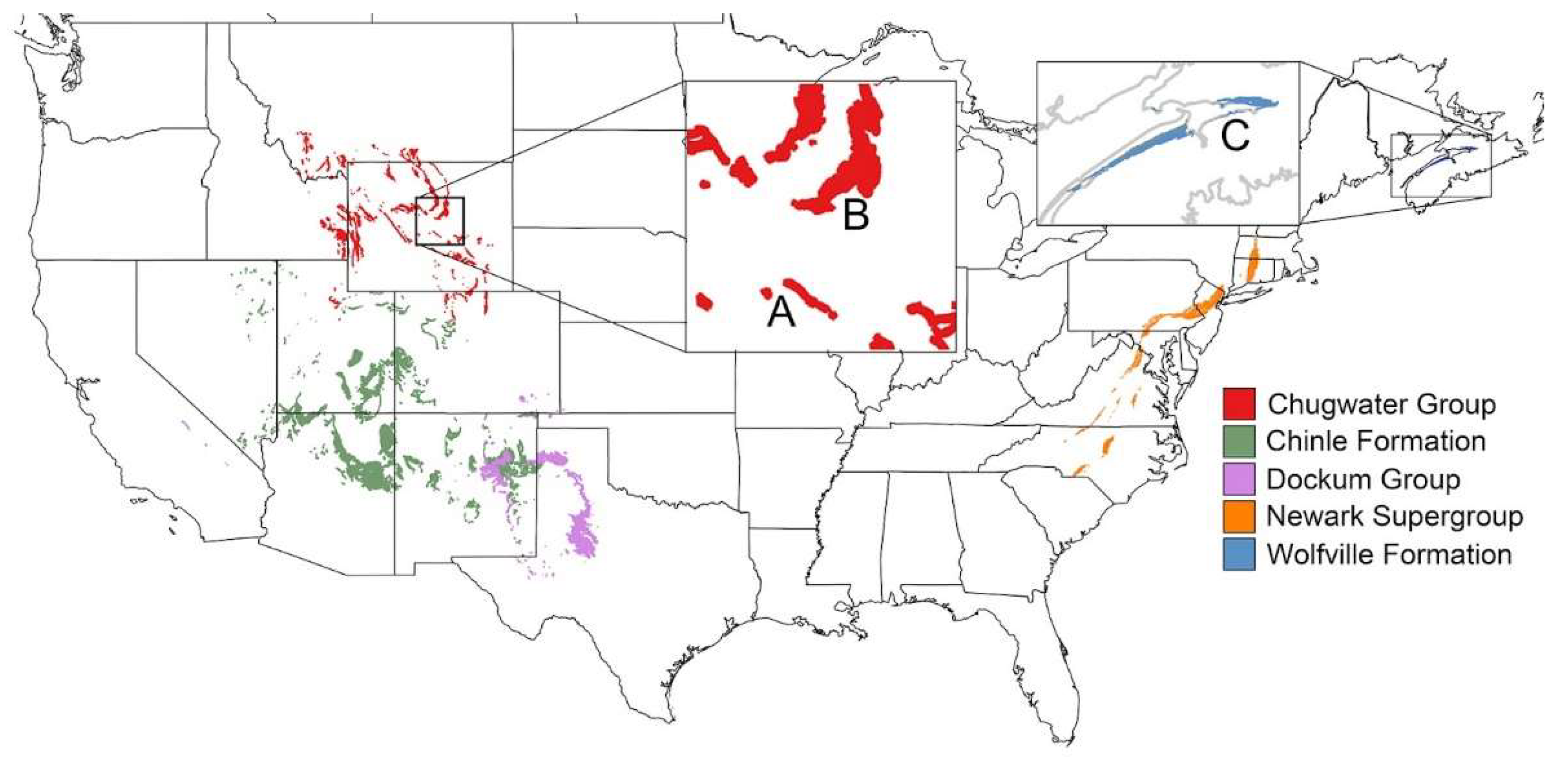
2.3. Geological Setting
2.4. Fossil Localities
2.4.1. Hole in the Wall
2.4.2. Cottonwood Creek
2.4.3. Stratigraphic Methods
3. Results
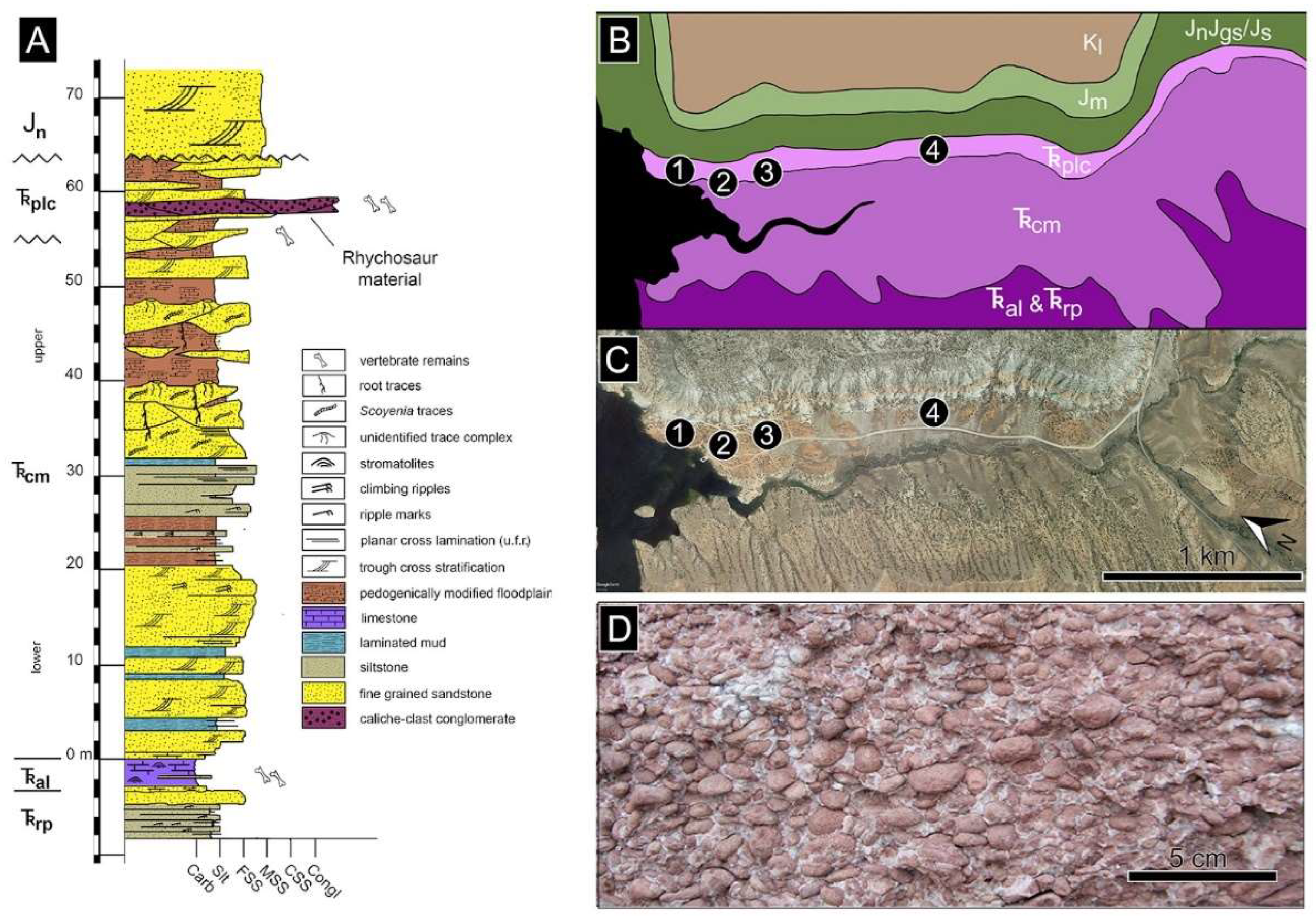
3.1. Stratigraphy
3.2. Age Assignment
3.3. Anatomic and Taxonomic Terminology
4. Systematic Paleontology
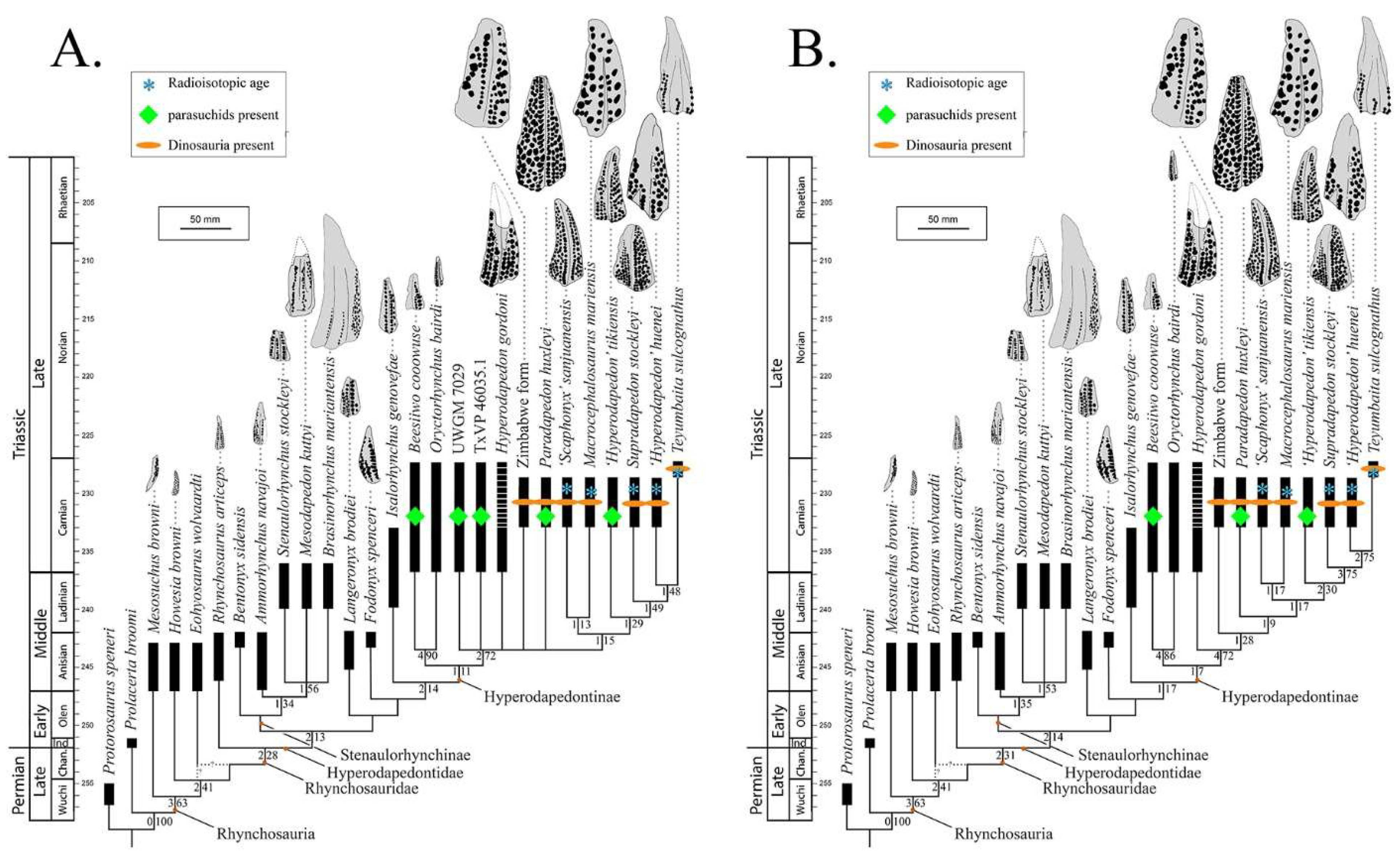
Phylogenetic Relationships
- 124. Lacrimal, contacts the ascending process of the palatine: (0) absent; (1) present.
- 125. Maxilla, occlusal view, labiolingual distance between longitudinal groove apex and L1 tooth row: (0) less than the labiolingual width of L1 teeth; (1) equal to or greater than the labiolingual width of L1 teeth.
- 126. Maxilla, ascending process: (0) anterodorsal–posteroventral orientation of contact with the prefrontal and/or lacrimal; (1) nearly vertical orientation of contact with the prefrontal and/or lacrimal.
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.-X. The ICS International Chronostratigraphic Chart. Int. Comm. Stratigr. Episodes 2013, 36, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Dal Corso, J.; Bernardi, M.; Sun, Y.; Song, H.; Seyfulla, L.J.; Preto, N.; Gianolla, P.; Ruffell, A.; Kustatscher, E.; Roghi, G.; et al. Extinction and Dawn of the Modern World in the Carnian (Late Triassic). Sci. Adv. 2020, 6, eaba0099. [Google Scholar] [CrossRef] [PubMed]
- Dubiel, R.F. Triassic Deposystems, Paleogeography, and Paleoclimate of the Western Interior; AAPG Publications: Tulsa, Oklahoma, 1994; pp. 133–168. [Google Scholar]
- Lovelace, D.M. Triassic Macrostratigraphy of the Western United States and the Ichnology, Paleoecology, and Paleoenvironments of the Early and Middle Triassic of the Chugwater Group, Wyoming. Ph.D. Thesis, University of Wisconsin-Madison, Madison, WI, USA, 2012. [Google Scholar]
- Lovelace, D. Triassic Macrostratigraphy of the Western United States. In STRATI 2013; Rocha, R., Pais, J., Kullberg, J.C., Finney, S., Eds.; Springer Geology; Springer International Publishing: Cham, Switzerland, 2014; ISBN 978-3-319-04363-0. [Google Scholar]
- Chatterjee, S. A Rhynchosaur from the Upper Triassic Maleri Formation of India. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1974, 267, 209–261. [Google Scholar] [PubMed]
- Langer, M.C.; Schultz, C.L. A New Species of the Late Triassic Rhynchosaur Hyperodapedon from the Santa Maria Formation of South Brazil. Palaeontology 2000, 43, 633–652. [Google Scholar] [CrossRef]
- Zerfass, H.; Lavina, E.L.; Schultz, C.L.; Garcia, A.J.V.; Faccini, U.F.; Chemale, F. Sequence Stratigraphy of Continental Triassic Strata of Southernmost Brazil: A Contribution to Southwestern Gondwana Palaeogeography and Palaeoclimate. Sediment. Geol. 2003, 161, 85–105. [Google Scholar] [CrossRef]
- Whatley, R.L. Phylogenetic Relationships of Isalorhynchus genovefae, the Rhynchosaur (Reptilia, Archosauromorpha) from Madagascar. Ph.D. Thesis, University of California, Santa Barbara, CA, USA, 2005. [Google Scholar]
- Mukherjee, D.; Ray, S. A New Hyperodapedon (Archosauromorpha, Rhynchosauria) from the Upper Triassic of India: Implications for Rhynchosaur Phylogeny. Palaeontology 2014, 57, 1241–1276. [Google Scholar] [CrossRef]
- Butler, R.J.; Ezcurra, M.D.; Montefeltro, F.C.; Samathi, A.; Sobral, G. A New Species of Basal Rhynchosaur (Diapsida: Archosauromorpha) from the Early Middle Triassic of South Africa, and the Early Evolution of Rhynchosauria: New Basal Rhynchosaur from South Africa. Zool. J. Linn. Soc. 2015, 174, 571–588. [Google Scholar] [CrossRef] [Green Version]
- Schultz, C.L.; Langer, M.C.; Montefeltro, F.C. A New Rhynchosaur from South Brazil (Santa Maria Formation) and Rhynchosaur Diversity Patterns across the Middle-Late Triassic Boundary. Palz 2016, 90, 593–609. [Google Scholar] [CrossRef] [Green Version]
- Langer, M.C.; da Rosa, Á.A.S.; Montefeltro, F.C. Supradapedon Revisited: Geological Explorations in the Triassic of Southern Tanzania. PeerJ 2017, 5, e4038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desojo, J.B.; Fiorelli, L.E.; Ezcurra, M.D.; Martinelli, A.G.; Ramezani, J.; Da Rosa, Á.A.S.; von Baczko, M.B.; Trotteyn, M.J.; Montefeltro, F.C.; Ezpeleta, M.; et al. The Late Triassic Ischigualasto Formation at Cerro Las Lajas (La Rioja, Argentina): Fossil Tetrapods, High-Resolution Chronostratigraphy, and Faunal Correlations. Sci. Rep. 2020, 10, 12782. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.P.; Lucas, S.G. A New Rynchosaur from the Upper Triassic of West Texas, and the Biochronology of Late Triassic Rynchosaurs. Paleontology 1991, 34, 927–938. [Google Scholar]
- Spielmann, J.; Lucas, S.; Hunt, A. The First Norian (Revueltian) Rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A. N. M. Mus. Nat. Hist. Sci. Bull. 2013, 61, 562–566. [Google Scholar]
- Ezcurra, M.D.; Montefeltro, F.; Butler, R.J. The Early Evolution of Rhynchosaurs. Front. Ecol. Evol. 2016, 3, 142. [Google Scholar] [CrossRef] [Green Version]
- Sues, H.-D.; Fitch, A.J.; Whatley, R.L. A New Rhynchosaur (Reptilia, Archosauromorpha) from the Upper Triassic of Eastern North America. J. Vertebr. Paleontol. 2020, 40, e1771568. [Google Scholar] [CrossRef]
- Nesbitt, S.J.; Stocker, M.R.; Ezcurra, M.D.; Fraser, N.C.; Heckert, A.B.; Parker, W.G.; Mueller, B.; Sengupta, S.; Bandyopadhyay, S.; Pritchard, A.C.; et al. Widespread Azendohsaurids (Archosauromorpha, Allokotosauria) from the Late Triassic of Western USA and India. Pap. Palaeontol. 2022, 8, e1413. [Google Scholar] [CrossRef]
- Benton, M.J. The Triassic Reptile Hyperodapedon from Elgin: Functional Morphology and Relationships. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1983, 302, 605–718. [Google Scholar] [CrossRef]
- Huxley, T.H. Postscript to: On the Sandstones of Morayshire (Elgin, &c.) Containing Reptillian Remains; and on Their Relations to the Old Red Sandstone of That Country (by R.J. Murchison). Q. J. Geol. Soc. 1859, 15, 435–436. [Google Scholar] [CrossRef]
- Lucas, S.G.; Heckert, A.B.; Hotton, N., III. The Rhynchosaur Hyperodapedon from the Upper Triassic of Wyoming and Its Global Biochronological Significance. N. M. Mus. Nat. Hist. Sci. Bull. 2002, 21, 149–156. [Google Scholar]
- High, L.R.; Picard, M.D. Sedimentary Petrology and Origin of Analcime-Rich Popo Agie Member, Chugwater (Triassic) Formation, West-Central Wyoming. J. Sediment. Petrol. 1965, 35, 49–70. [Google Scholar] [CrossRef]
- McCormick, C.D.; Picard, M.D. Petrology of Gartra Formation (Triassic), Uinta Mountain Area, Utah and Colorado. J. Sediment. Petrol. 1969, 39, 1484–1508. [Google Scholar]
- Colbert, E.H. Triassic Vertebrates of the Wind River Basin. Southwest Wind River Basin. Wyo. Geol. Assoc. Guideb. 1957, 12, 89–97. [Google Scholar]
- Hunt, A.P.; Lucas, S.P. The Paleorhinus Biochron and the Correlation of the Non-Marine Upper Triassic of Pangaea. Palaeontology 1991, 34, 17. [Google Scholar]
- Lucas, S.G.; Heckert, A.B.; Rinehart, L. A Giant Skull, Ontogenetic Variation and Taxonomic Validity of the Late Triassic Phytosaur Parasuchus. N. M. Mus. Nat. Hist. Sci. Bull. 2007, 41, 222–228. [Google Scholar]
- Lovelace, D.M.; Fitch, A.J. Consilience of Late Triassic Lithostratigraphic, Radioisotopic, and Biostratigraphic Data Support a Carnian Age for the Popo Agie Formation (Chugwater Group), Wyoming. GSA Bull. in revision.
- Sues, H.-D.; Olsen, P.E. Stratigraphic and Temporal Context and Faunal Diversity of Permian-Jurassic Continental Tetrapod Assemblages from the Fundy Rift Basin, Eastern Canada. Atl. Geol. 2015, 51, 139–168. [Google Scholar] [CrossRef] [Green Version]
- Lucas, S.G. The Beginning of the Age of Dinosaurs in Wyoming. Wyo. Geol. Assoc. Guideb. 1994, 44, 105–113. [Google Scholar]
- Schoch, R.R. The Capitosauria (Amphibia): Characters, Phylogeny, and Stratigraphy. Palaeodiversity 2008, 1, 189–226. [Google Scholar]
- Pipiringos, G.N. Correlation and Nomenclature of Some Triassic and Jurassic Rocks in South-Central Wyoming; Geological Survey Professional Papers; United States Government Printing Office: Washington, DC, USA, 1968; Volume 594-D. [Google Scholar]
- High, L.R.; Picard, M.D. Stratigraphic Relations of Upper Triassic Units, Northeastern Utah and Wyoming. Compass Sigma Gamma Epsil. 1967, 44, 88–98. [Google Scholar]
- Picard, M.D. Stratigraphy of Triassic Rocks in West-Central, Wyoming. Wyo. Geol. Assoc. Guideb. 1978, 30, 101–130. [Google Scholar]
- Cavaroc, V.V.; Flores, R.M. Red Beds of the Triassic Chugwater Group, Southwestern Powder River Basin, Wyoming. US Geol. Surv. Bull. 1990, 1917, 1–18. [Google Scholar]
- High, L.R., Jr.; Picard, M.D. Stratigraphic Relations Within Upper Chugwater Group (Triassic), Wyoming. AAPG Bull. 1969, 53, 1091–1104. [Google Scholar] [CrossRef]
- Irmen, A.P.; Vondra, C.F. Aeolian Sediments in Lower to Middle (?) Triassic Rocks of Central Wyoming. Sediment. Geol. 2000, 132, 69–88. [Google Scholar] [CrossRef]
- High, L.R.; Picard, M.D. Rock units and Revised Nomenclature Chugwater Group (Triassic), Western Wyoming. Mt. Geol. 1967, 4, 73–81. [Google Scholar]
- DeCelles, P.G.; Giles, K.A. Foreland Basin Systems. Basin Res. 1996, 8, 105–123. [Google Scholar] [CrossRef]
- McLellan, R. Stratigraphy of Chugwater Group (Triassic), East Side of Bighorn Mountains, Wyoming and Montana. Master’s Thesis, University of Nebraska, Lincoln, NE, USA, 1968. [Google Scholar]
- Johnson, E.A. Depositional History of Triassic Rocks in the Area of the Powder River Basin, Northeastern Wyoming and Southeastern Montana; USGS Publications Warehouse: Reston, VA, USA, 1993. [Google Scholar]
- Keller, W.D. Analcime in the Popo Agie Member of the Chugwater Formation. Science 1952, 115, 241–242. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.D.; High, L.R.; Jerseyl, N. Mineral Assemblages in the Popo Agie Member of the Chugwater Formation, West-Central Wyoming. Contrib. Geol. 1963, 2, 125–128. [Google Scholar]
- Deckman, M.E.; Lovelace, D.M.; Rogers, R.R. Stratigraphy, Sedimentary Petrology, and Depositional Environment of the Chugwater Group, near Dubois, Wyoming. Geol. Soc. Am. Abstr. Program 2019, 51, 202–209. [Google Scholar] [CrossRef]
- Deckman, M.; Lovelace, D.M.; Holland, S.M. Lateral Variability of Splay Deposits in a Distributive Fluvial System: Upper Triassic Chugwater Group, Fremont County, Wyoming. Geol. Soc. Am. Abstr. Program 2021, 53, 22–28. [Google Scholar] [CrossRef]
- Deckman, M.E. A Reinterpretation of the Triassic Crow Mountain and Popo Agie Formations as a Distributive Fluvial System, Wind River Basin, Wyoming. Master’s Thesis, University of Georgia, Athens, GA, USA, 2022. [Google Scholar]
- Dawley, R.M.; Zawiskie, J.M.; Cosgriff, J.W. A Rauisuchid Thecodont from the Upper Triassic Popo Agie Formation of Wyoming. J. Paleontol. 1979, 53, 1428–1431. [Google Scholar]
- Nesbitt, S.J.; Zawiskie, J.M.; Dawley, R.M. The Osteology and Phylogenetic Position of the Loricatan (Archosauria: Pseudosuchia) Heptasuchus clarki, from the ?Mid-Upper Triassic, Southeastern Big Horn Mountains, Central Wyoming (USA). PeerJ 2020, 8, e10101. [Google Scholar] [CrossRef]
- Lovelace, D.M.; Lovelace, S.D. Paleoenvironments and Paleoecology of a Lower Triassic Invertebrate and Vertebrate Ichnoassemblage from the Red Peak Formation (Chugwater Group), Central Wyoming. Palaios 2012, 27, 636–657. [Google Scholar] [CrossRef]
- Lovelace, D.M.; Doebbert, A.C. A New Age Constraint for the Early Triassic Alcova Limestone (Chugwater Group), Wyoming. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 424, 1–5. [Google Scholar] [CrossRef]
- Storrs, G.W. Anatomy and Relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcova Limestone of Wyoming. Yale Peabody Mus. Bull. 1991, 44, 162. [Google Scholar]
- Rieppel, O. Corosaurus alcovensis Case and the Phylogenetic Interrelationships of Triassic Stem-Group Sauropterygia. Zool. J. Linn. Soc. 1998, 124, 1–41. [Google Scholar] [CrossRef]
- Smith, J.J.; Hasiotis, S.T.; Kraus, M.J.; Woody, D.T. Naktodemasis browni: New Ichnogenus and Ichnospecies for Adhesive Meniscate Burrows (AMB), and Paleoenvironmental Implications, Paleogene Willwood Formation, Bighorn Basin, Wyoming. J. Paleontol. 2008, 82, 267–278. [Google Scholar] [CrossRef]
- Smith, J.J.; Hasiotis, S.T. Traces and Burrowing Behaviors of the Cicada Nymph Cicadetta Calliope: Neoichnology and Paleoecological Significance of Extant Soil-Dwelling Insects. Palaios 2008, 23, 503–513. [Google Scholar] [CrossRef]
- Williston, S.W. Notice of Some New Reptiles from the Upper Trias of Wyoming. J. Geol. 1904, 12, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Mehl, M.G. Angistorhinus, a New Genus of Phytosauria from the Trias of Wyoming. J. Geol. 1913, 21, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Mehl, M.G. The Phytosauria of the Wyoming Triassic. J. Denison Univ. Lab. Denison Univ. 1928, 23, 141–172. [Google Scholar]
- Eaton, T.H. A New Wyoming Phytosaur. Eaton Univ. Kans. Paleontol. Contrib. 1965, 2, 1–6. [Google Scholar]
- Kammerer, C.F.; Butler, R.J.; Bandyopadhyay, S.; Stocker, M.R. Relationships of the Indian Phytosaur Parasuchus hislopi Lydekker, 1885. Pap. Palaeontol. 2016, 2, 1–23. [Google Scholar] [CrossRef]
- Fitch, A.J.; Kufner, A.M.; Stocker, M.R.; Price, B.R.; Lovelace, D. New Material from the Popo Agie Formation of Wyoming Yields Insights into Basal Phytosaurs and the Late Carnian Fuana of the Western U.S.A. In Proceedings of the 78th Annual Meeting of Society of Vertebrate Paleontology, Albuquerque, NM, USA, 17–20 October 2018; Volume 127. [Google Scholar]
- Jones, A.S.; Butler, R.J. A New Phylogenetic Analysis of Phytosauria (Archosauria: Pseudosuchia) with the Application of Continuous and Geometric Morphometric Character Coding. PeerJ 2018, 6, e5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martz, J.W.; Parker, W.G. Revised Formulation of the Late Triassic Land Vertebrate “Faunachrons” of Western North America. In Terrestrial Depositional Systems; Elsevier: Amsterdam, The Netherlands, 2017; pp. 39–125. ISBN 978-0-12-803243-5. [Google Scholar]
- Rasmussen, C.; Mundil, R.; Irmis, R.B.; Geisler, D.; Gehrels, G.E.; Olsen, P.E.; Kent, D.V.; Lepre, C.; Kinney, S.T.; Geissman, J.W.; et al. U-Pb Zircon Geochronology and Depositional Age Models for the Upper Triassic Chinle Formation (Petrified Forest National Park, Arizona, USA): Implications for Late Triassic Paleoecological and Paleoenvironmental Change. GSA Bull. 2021, 133, 539–558. [Google Scholar] [CrossRef]
- Chatterjee, S. Rhynchosaurs in Time and Space. Proc. Geol. Soc. Lond. 1969, 1658, 203–208. [Google Scholar]
- Benton, M.J. The Species of Rhynchosaurus, A Rhynchosaur (Reptilia, Diapsida) from the Middle Triassic of England. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1990, 328, 213–306. [Google Scholar]
- Hone, D.W.E.; Benton, M.J. A New Genus of Rhynchosaur from the Middle Triassic of South-West England: New Genus of Mid Triassic Rhynchosaur from England. Palaeontology 2008, 51, 95–115. [Google Scholar] [CrossRef]
- Montefeltro, F.C.; Langer, M.C.; Schultz, C.L. Cranial Anatomy of a New Genus of Hyperodapedontine Rhynchosaur (Diapsida, Archosauromorpha) from the Upper Triassic of Southern Brazil. Earth Environ. Sci. Trans. R. Soc. Edinb. 2010, 101, 27–52. [Google Scholar] [CrossRef] [Green Version]
- Raath, M.A.; Oesterlen, P.M.; Kitching, J.W. First Record of Triassic Rhynchosauria (Reptilia: Diapsida) from the Lower Zambezi Valley, Zimbabwe. Palaeontol. Afr. 1992, 29, 1–10. [Google Scholar]
- von Huene, F. Die Fossilen Reptilien Des Südamerikanischen Gondwanalandes an Der Zeitenwende: Ergebnisse d. Sauriergrabungen in Südbrasilien 1928/29; C.H. Becksche Verlagsbuchhandlung: München, Germany, 1942. [Google Scholar]
- Sill, W.D. Scaphonyx san]uanensis, Nuevo Rincosaurio (Reptilia) De La Formacion Ischigualasto, Triasico De San Juan, Argentina. Ameghiniana 1970, 7, 341–354. [Google Scholar]
- Langer, M.C. Gilmoreteiidae New Family and Gilmoreteius New Genus (Squamata, Scincomorpha): Replacement Names for Macrocephalosauridae Sulimski, 1975 and Macrocephalosaurus Gilmore, 1943. Comun. Mus. Cienc. E Tecnol. PUCRS Ser. Zool. 1998, 11, 13–18. [Google Scholar]
- Langer, M.; Boniface, M.; Cuny, G.; Barbieri, L. The Phylogenetic Position of Isalorhynchus genovefae, a Late Triassic Rhynchosaur from Madagascar. Ann. Paléontol. 2000, 86, 101–127. [Google Scholar] [CrossRef]
- Woodward, A.S. III.—On Some Fossil Reptilian Bones from the State of Rio Grande Do Sul, Brazil2. Geol. Mag. 1908, 5, 251–255. [Google Scholar] [CrossRef] [Green Version]
- von Huene, F. Gondwana-Reptilien in Südamerika. Paleontol. Hung. 1926, II, 102pp. [Google Scholar]
- Tupí-Caldas, J.L. Contribuição Ao Estudo Do Fóssil Da Alemoa, Municipio de Santa Maria, Rio Grande Do Sul. In Curso Geral de Mineralogia e Geologia, aplicada ao Brasil, 2nd ed.; Porto Alegre Edições Livraria Globo: Porto Alegre, Brasil, 1933; pp. 333–339. [Google Scholar]
- von Huene, F. Die Fossilen Reptilien Des Südamerikanischen Gondwanalandes. Neues Jahrb. Für Geol. Paläontol. Abt. B 1938, 1938, 142–151. [Google Scholar]
- Chatterjee, S. The Evolution of Rhynchosaurs. Mém. Société Géologique Fr. Nouv. Sér. 1980, 139, 57–65. [Google Scholar]
- Buffetaut, E. Isalorhynchus genovefae, n. g. n. sp. (Reptilia, Rhynchocephalia), un nouveau Rhynchosaure du Trias de Madagascar. Neues Jahrb. Für Geol. Paläontol. Mon. 1983, 8, 465–480. [Google Scholar] [CrossRef]
- McCartney, J. Preface and Table III. In Lectures on Comparative Anatomy; Wilson and Co., for T. N. Longman and O. Rees, Paternoster Row; Oriental Press: London, UK, 1802. [Google Scholar]
- Gauthier, J.; Kluge, A.G.; Rowe, T. Amniote Phylogeny and the Importance of Fossils. Cladistics 1988, 4, 105–209. [Google Scholar] [CrossRef] [Green Version]
- von Huene, F. Die Grossen Stämme Der Tetrapoden in Den Geologischen Zeiten. Biol. Zentralblatt 1946, 65, 268–275. [Google Scholar]
- Dilkes, D.W. The Early Triassic Rhynchosaur Mesosuchus browni and the Interrelationships of Basal Archosauromorph Reptiles. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1998, 353, 501–541. [Google Scholar] [CrossRef] [Green Version]
- Cope, E.D. On the Homologies of Some of the Cranial Bones of the Reptilia: And on the Systematic Arrangement of the Class; J. Lovering: London, UK, 1871. [Google Scholar]
- Lydekker, R. The Reptilia & Amphibia of the Maleri & Denwa Groups; Geological Survey Office: Reston, VA, USA, 1885. [Google Scholar]
- Cowell, A.; Moss, A.; C’Hair, W.; C’Hair, W. Elders of the Norther Arapaho Tribe Dictionary of the Arapaho Language; Northern Arapaho Tribe: Ethete, WY, USA, 2012. [Google Scholar]
- Benton, M.J. Tooth Form, Growth and Function in Triassic Rhynchosaurs (Reptilia, Diapsida). Palaeontology 1984, 27, 737–776. [Google Scholar]
- Haughton, S.H. On a Collection of Karoo Vertebrates from Tanganyika Territory. Q. J. Geol. Soc. 1932, 88, 634–671. [Google Scholar] [CrossRef]
- Langer, M.C.; Ferigolo, J.; Schultz, C.L. Heterochrony and Tooth Evolution in Hyperodapedontine Rhynchosaurs (Reptilia, Diapsida). Lethaia 2000, 33, 119–128. [Google Scholar] [CrossRef]
- Nesbitt, S.J.; Whatley, R.L. The First Discovery of a Rhynchosaur from the Upper Moenkopi Formation (Middle Triassic) of Northern Arizona. PaleoBios 2004, 24, 1–10. [Google Scholar]
- Dilkes, D.W. The Ryhnchosaur Howesia browni from the Lower Triassic of South Africa. Palaeontology 1995, 38, 665–685. [Google Scholar]
- Langer, M.C.; Montefeltro, F.C.; Hone, D.E.; Whatley, R.; Schultz, C.L. On Fodonyx spenceri and a New Rhynchosaur from the Middle Triassic of Devon. J. Vertebr. Paleontol. 2010, 30, 1884–1888. [Google Scholar] [CrossRef]
- Griffin, C.T.; Wynd, B.M.; Munyikwa, D.; Broderick, T.J.; Zondo, M.; Tolan, S.; Langer, M.C.; Nesbitt, S.J.; Taruvinga, H.R. Africa’s Oldest Dinosaurs Reveal Early Suppression of Dinosaur Distribution. Nature 2022, 609, 313–319. [Google Scholar] [CrossRef]
- Gentil, A.R.; Ezcurra, M.D. Skull Osteology of the Holotype of the Rhynchosaur Hyperodapedon sanjuanensis (Sill, 1970) from the Upper Triassic Ischigualasto Formation of Argentina. Anat. Rec. 2022, 305, 1168–1200. [Google Scholar] [CrossRef]
- Gow, C.E. The Morphology and Relationships of Youngina capensis Broom and Prolacerta broomi Parrington. Palaeontol. Afr. 1975, 18, 89–131. [Google Scholar]
- Modesto, S.P.; Sues, H.-D. The Skull of the Early Triassic Archosauromorph Reptile Prolacerta broomi and Its Phylogenetic Significance. Zool. J. Linn. Soc. 2004, 140, 335–351. [Google Scholar] [CrossRef]
- Gottmann-Quesada, A.; Sander, P.M. A Redescription of the Early Archosauromorph Protorosaurus speneri Meyer, 1832, and Its Phylogenetic Relationships. Palaeontogr. Abt. A 2009, 287, 123–220. [Google Scholar] [CrossRef]
- Gentil, A.R.; Ezcurra, M.D. Reconstruction of the Masticatory Apparatus of the Holotype of the Rhynchosaur Hyperodapedon sanjuanensis from the Late Triassic of Argentina: Implications for the Diagnosis of the Species. Ameghiniana 2017, 55, 137–149. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a Free Program for Phylogenetic Analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Catalano, S.A. TNT Version 1.5, Including a Full Implementation of Phylogenetic Morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Langer, M.C.; Ribeiro, A.M.; Schultz, C.L.; Ferigolo, J. The Continental Tetrapod-Bearing Triassic of South Brazil. N. M. Mus. Nat. Hist. Sci. Bull. 2007, 41, 201–217. [Google Scholar]
- Flynn, J.J.; Parrish, J.M.; Rakotosamimanana, B.; Ranivoharimanana, L.; Simpson, W.F.; Wyss, A.R. New Traversodontids (Synapsida: Eucynodontia) from the Triassic of Madagascar. J. Vertebr. Paleontol. 2000, 20, 422–427. [Google Scholar] [CrossRef]
- Philipp, R.P.; Schultz, C.L.; Kloss, H.P.; Horn, B.L.D.; Soares, M.B.; Basei, M.A.S. Middle Triassic SW Gondwana Paleogeography and Sedimentary Dispersal Revealed by Integration of Stratigraphy and U-Pb Zircon Analysis: The Santa Cruz Sequence, Paraná Basin, Brazil. J. South Am. Earth Sci. 2018, 88, 216–237. [Google Scholar] [CrossRef]
- Langer, M.C.; Ramezani, J.; Da Rosa, Á.A.S. U-Pb Age Constraints on Dinosaur Rise from South Brazil. Gondwana Res. 2018, 57, 133–140. [Google Scholar] [CrossRef]
- Benton, M.J.; Walker, A.D. Saltopus, a Dinosauriform from the Upper Triassic of Scotland. Earth Environ. Sci. Trans. R. Soc. Edinb. 2010, 101, 285–299. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fitch, A.J.; Haas, M.; C’Hair, W.; Ridgley, E.; Ridgley, B.; Oldman, D.; Reynolds, C.; Lovelace, D.M. A New Rhynchosaur Taxon from the Popo Agie Formation, WY: Implications for a Northern Pangean Early-Late Triassic (Carnian) Fauna. Diversity 2023, 15, 544. https://doi.org/10.3390/d15040544
Fitch AJ, Haas M, C’Hair W, Ridgley E, Ridgley B, Oldman D, Reynolds C, Lovelace DM. A New Rhynchosaur Taxon from the Popo Agie Formation, WY: Implications for a Northern Pangean Early-Late Triassic (Carnian) Fauna. Diversity. 2023; 15(4):544. https://doi.org/10.3390/d15040544
Chicago/Turabian StyleFitch, Adam J., Merle Haas, Wayne C’Hair, Eugene Ridgley, Ben Ridgley, Devin Oldman, Crystal Reynolds, and David M. Lovelace. 2023. "A New Rhynchosaur Taxon from the Popo Agie Formation, WY: Implications for a Northern Pangean Early-Late Triassic (Carnian) Fauna" Diversity 15, no. 4: 544. https://doi.org/10.3390/d15040544