
Learn the Past and Present to Teach the Future—Role of Active Surveillance of Exposure to Endemic and Emerging Viruses in the Approach of European Bison Health Protection

 , , and
, , and
Abstract
:1. Introduction
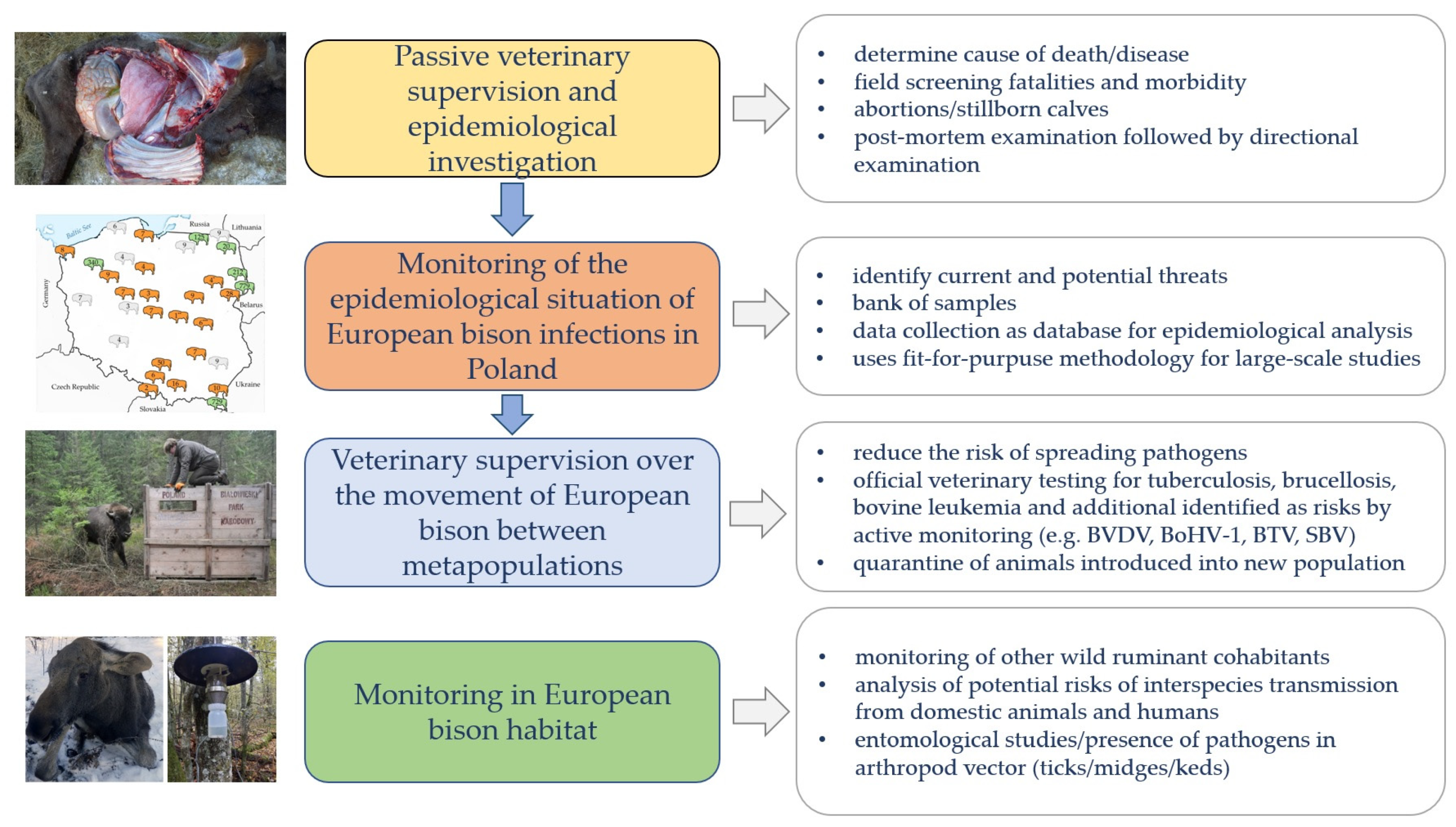
2. Materials and Methods
2.1. Study Design
2.2. Serological Testing
2.3. Molecular Testing
2.4. Statistical Analysis
3. Results
3.1. Descriptive Statistics
3.1.1. Viruses Affecting Reproduction
3.1.2. Respiratory Viruses
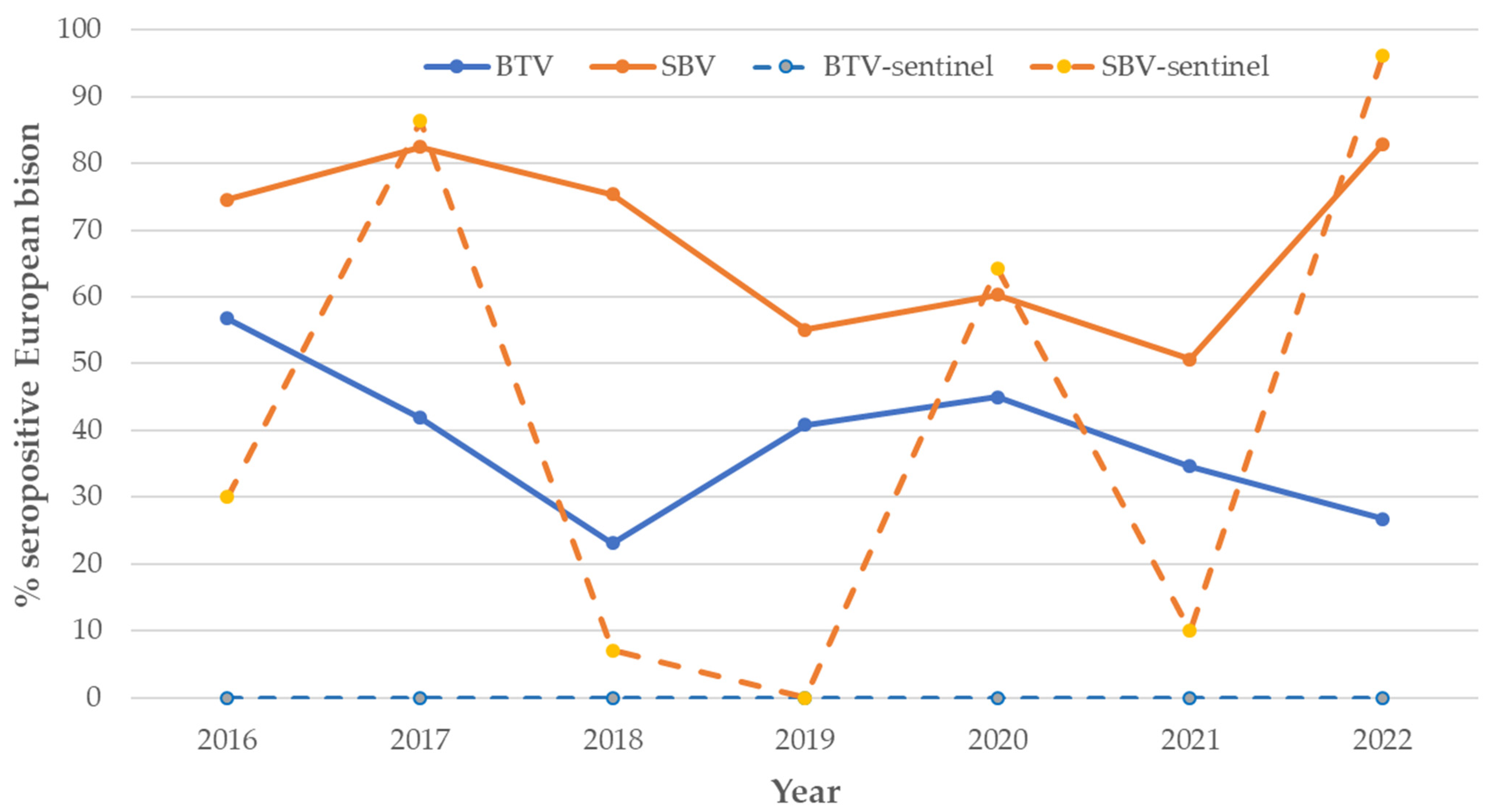
3.1.3. BTV and SBV—Representatives of Arboviruses
3.2. Factors Affecting Potential Interspecies Transmission from Cattle and Intraspecies Transmission among European Bison
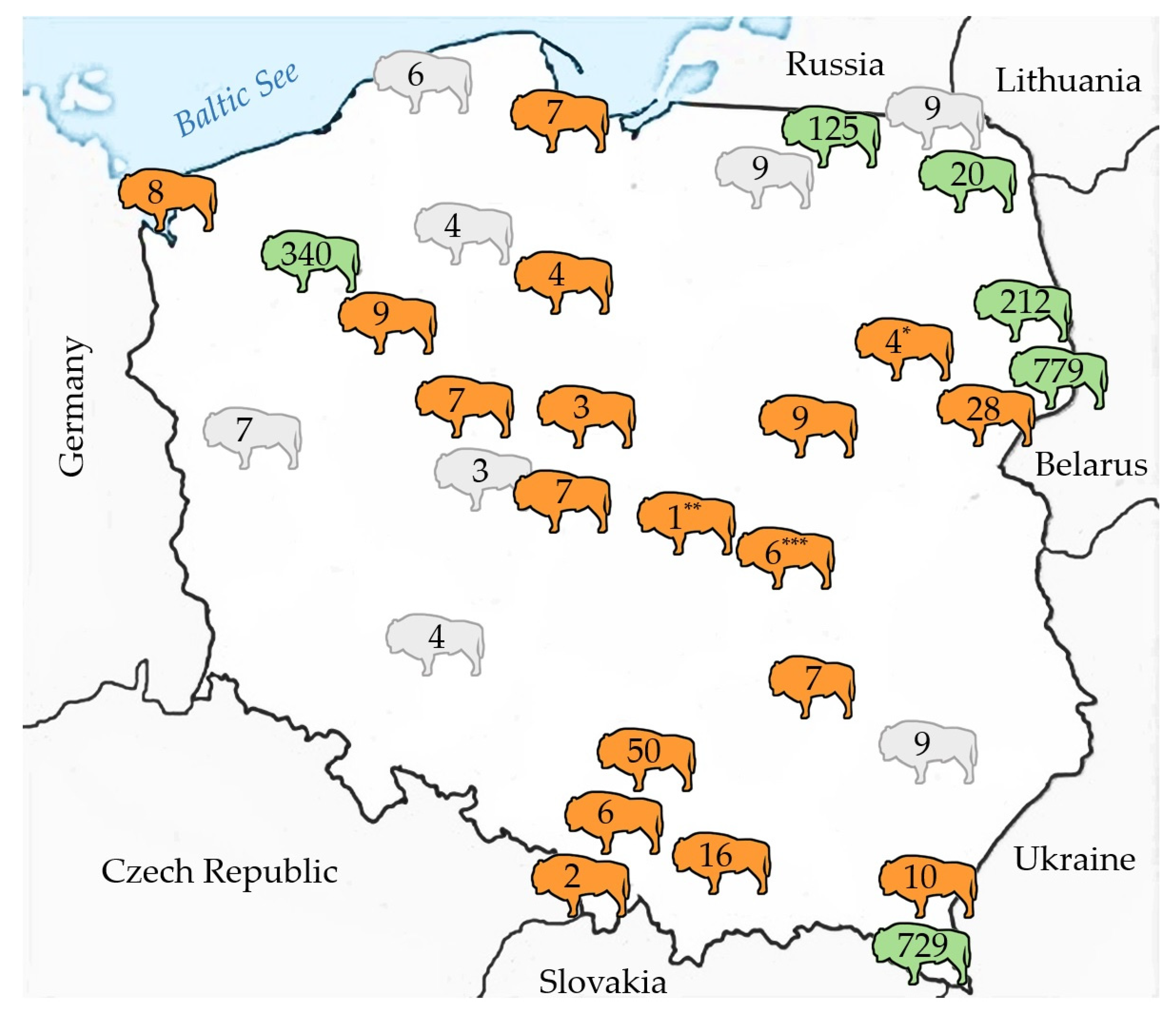
- BVDV and BoHV-1 spread from cattle (reservoir species) to European bison; however, on the basis of the results, the neighbour transmission is sporadic and concerns the largest free-living populations such as the Białowieska Forest (Table 1). Other explanations such as low susceptibility of European bison to BVDV and BoHV-1 infection, low prevalence of the infections in cattle in surrounding farms, and biosecurity/prophylactic measures present in neighbouring cattle farms should be considered.
- Potential direct contact between cattle and European bison was reflected by the presence of specific antibodies BoHV-4, mostly in the largest free-living populations with the highest seroprevalence in the Białowieska Forest (Table 1). The only exceptions were the Pszczyna and Białowieża captive populations, where two and one BoHV-4 seropositive bulls were detected, respectively.
- The higher seroprevalences of respiratory viruses in free-living animals also suggested possible direct spill over from cattle. However, some circulation, in particular of BRV3 and BAdV-3 was observed also in captive herds, which also corroborated independent, intraspecies circulation of these viruses in the European bison population.
- Higher seroprevalences in larger free-living populations for BRV3 and in larger captive populations for BAdV-3 suggested a density-dependent mode of transmission.
- Surprisingly, exposure to BRV3 and BRSV was higher in smaller captive herds such as ZOOs or small show enclosures, which suggested the source of infection within the herd and where European bison have multi-species contacts with other non-domestic animals, including exotic ones. Another possibility may be indirect exposure connected to the passive transmission of pathogens by human staff and visitors of the enclosures.
- Arthropod viruses BTV and SBV were more widely spread in free-ranging European bison; however, since the transmission is vector-dependent, the environmental conditions would be more likely to have an impact, rather than possible direct contact with cattle.
- Highest SBV seroprevalence in the medium and largest free-ranging populations suggests density dependence; however, significant variation in SBV seroprevalence in all and in sentinel (naïve to infection, 6 months—1 year old calves) European bison by year also indicated frequency-dependence.
3.3. Anthropogenic Factors Affecting Virus Transmission
3.4. Risk Factors
4. Discussion
5. Conclusions
- No significant increase in the interspecies and intraspecies risk of endemic viral infections was found in European bison in Poland in the last seven years;
- The free-living and larger European bison populations are more frequently exposed to some viral pathogens which impact their health;
- A new direction of research should include Bovine herpesvirus type 4 (BoHV-4) that may be relevant to the aetiology of the necrotic balanoposthitis in European bison bulls;
- No effect of anthropogenic factors on the transmission rates and frequency of selected viral infections was observed;
- Vector-borne bluetongue virus (BTV) is probably no longer circulating in the environment, while infection with emerging Schmallenberg virus (SBV) has become endemic with waves of seroprevalence increasing every 3–4 years;
- Veterinary supervision and passive monitoring of European bison herds is crucial as it leads to the identification and elimination of infected individuals and prevents further transmission;
- Active surveillance should be continued in order to monitor threats and to establish a further conservation strategy for the species.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raczyński, J. European Bison Pedigree Book 2016–2021; Białowieża National Park: Białowieża, Poland, 2017–2022; Available online: https://bpn.com.pl/index.php?option=com_content&task=view&id=1133&Itemid=213 (accessed on 3 February 2023).
- The International Union for Conservation of Nature (IUCN) Red List of Threatened Species 2020. Available online: https://www.iucnredlist.org/species/2814/45156279 (accessed on 3 February 2023).
- Wróblewski, K. Żubr Puszczy Białowieskiej; ZOO Garden Poznań: Poznań, Poland, 1927. [Google Scholar]
- Krasińska, M.; Krasiński, Z.A. Żubr, 2nd ed.; Drukarnia im. Półtawskiego: Kielce, Poland, 2017; 448p. [Google Scholar]
- Larska, M.; Krzysiak, M.K. Infectious diseases monitoring as an element of Bison bonasus species protection. In Compendium of the European Bison (Bison bonasus) Health Protection; Larska, M., Krzysiak, M.K., Eds.; National Veterinary Research Institute: Puławy, Poland, 2022; pp. 71–96. [Google Scholar]
- Krzysiak, M.K.; Dackiewicz, J.; Kęsik-Maliszewska, J.; Larska, M. Post-mortem evaluation of pathological lesions in European bison (Bison bonasus) in the Białowieża Primeval Forest between 2008 and 2013. J. Vet. Res. 2014, 58, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Krzysiak, M.K.; Jabłoński, A.; Iwaniak, W.; Krajewska, M.; Kęsik-Maliszewska, J.; Larska, M. Seroprevalence and risk factors for selected respiratory and reproductive tract pathogen exposure in European bison (Bison bonasus) in Poland. Vet. Microb. 2018, 215, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Krzysiak, M.K.; Iwaniak, W.; Kęsik-Maliszewska, J.; Olech, W.; Larska, M. Serological study of exposure to selected arthropod-borne pathogens in European bison (Bison bonasus) in Poland. Transbound. Emerg. Dis. 2017, 64, 1411–1423. [Google Scholar] [CrossRef] [PubMed]
- McCallum, H.; Barlow, N.; Hone, J. How should pathogen transmission be modelled? Trends Ecol. Evol. 2001, 16, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Telfer, S.; Kallio, E.R.; Burthe, S.; Cook, A.R.; Lambin, X.; Begon, M. Host-pathogen time series data in wildlife support a transmission function between density and frequency dependence. Proc. Natl. Acad. Sci. USA 2009, 106, 7905–7909. [Google Scholar] [CrossRef] [Green Version]
- Dohoo, I.; Martin, W.; Stryhn, H. Veterinary Epidemiologic Research, 2nd ed.; VER Inc.: Charlottetown, PE, Canada, 2010; 865p. [Google Scholar]
- Humphry, R.W.; Cameron, A.; Gunn, G.J. A practical approach to calculate sample size for herd prevalence surveys. Prev. Vet. Med. 2004, 65, 173–188. [Google Scholar] [CrossRef]
- Polska w Liczbach. Available online: https://www.polskawliczbach.pl/ (accessed on 3 February 2023).
- Brearley, G.; Rhodes, J.; Bradley, A.; Baxter, G.; Seabrook, L.; Lunney, D.; Liu, Y.; McAlpine, C. Wildlife disease prevalence in human-modified landscapes. Biol. Rev. Camb. Philos. Soc. 2013, 88, 427–442. [Google Scholar] [CrossRef]
- Rola, J.G.; Larska, M.; Grzeszuk, M.; Rola, J. Association between antibody status to bovine herpesvirus 1 and quality of milk in dairy herds in Poland. J. Dairy Sci. 2015, 98, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Rypuła, K.; Płoneczka-Janeczko, K.; Czopowicz, M.; Klimowicz-Bodys, M.D.; Shabunin, S.; Siegwalt, G. Occurrence of BVDV infection and the presence of potential risk factors in dairy cattle herds in Poland. Animals 2020, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Rypuła, K.; Płoneczka-Janeczko, K.; Kita, J.; Kumala, A.; Zmudziński, J.F. Seroprevalence of BHV-1 (bovine herpesvirus type 1) among non-vaccinated dairy cattle herds with respiratory disorders. Pol. J. Vet. Sci. 2012, 15, 561–563. [Google Scholar] [CrossRef] [Green Version]
- Rola, J.; Larska, M.; Socha, W.; Rola, J.G.; Materniak, M.; Urban-Chmiel, R.; Thiry, E.; Żmudziński, J.F. Seroprevalence of bovine herpesvirus 1 related alphaherpesvirus infections in free-living and captive cervids in Poland. Vet. Microbiol. 2017, 204, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frant, M.P.; Gal-Cisoń, A.; Bocian, Ł.; Ziętek-Barszcz, A.; Niemczuk, K.; Woźniakowski, G.; Szczotka-Bochniarz, A. African swine fever in wild boar (Poland 2020): Passive and active surveillance analysis and further perspectives. Pathogens 2021, 10, 1219. [Google Scholar] [CrossRef] [PubMed]
- Borchers, K.; Brackmann, J.; Wolf, O.; Rudolph, M.; Glatzel, P.; Krasinska, M.; Krasinski, Z.A.; Frölich, K. Virologic investigations of free-living European bison (Bison bonasus) from the Bialowieza Primeval Forest, Poland. J. Wildl. Dis. 2002, 38, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Wathes, D.C.; Oguejiofor, C.F.; Thomas, C.; Cheng, Z. Importance of viral disease in dairy cow fertility. Engineering 2020, 6, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Hławiczka, M.; Zuber, Ł.; Bielecki, W. An attempt to cure necrotic foreskin inflammation (NZN) posthitis in a European bison. In Żubry w Puszczy Augustowskiej; SMŻ: Augustów, Poland; pp. 45–48. Available online: https://smz.waw.pl/wp-content/uploads/2021/04/Conference-Materials.pdf (accessed on 7 February 2023).
- Thiry, E.; Dubuisson, J.; Bublot, M.; Van Bressem, M.F.; Pastoret, P.P. The biology of bovine herpesvirus-4 infection of cattle. DTW 1990, 97, 72–77. [Google Scholar]
- Li, H.; Gailbreath, K.; Flach, E.J.; Taus, N.S.; Cooley, J.; Keller, J.; Russell, G.C.; Knowles, D.P.; Haig, D.M.; Oaks, J.L.; et al. A novel subgroup of rhadinoviruses in ruminants. J. Ge.n Virol. 2005, 86, 3021–3026. [Google Scholar] [CrossRef] [PubMed]
- Dewals, B.; Gillet, L.; Gerdes, T.; Taracha, E.L.; Thiry, E.; Vanderplasschen, A. Antibodies against bovine herpesvirus 4 are highly prevalent in wild African buffaloes throughout eastern and southern Africa. Vet. Microb. 2005, 110, 209–220. [Google Scholar] [CrossRef]
- Kálmán, D.; Egyed, L. PCR detection of bovine herpesviruses from nonbovine ruminants in Hungary. J. Wildl. Dis. 2005, 41, 482–488. [Google Scholar] [CrossRef] [Green Version]
- Kędrak-Jablońska, A.; Budniak, S.; Szczawińska, A.; Reksa, M.; Krupa, M.; Krzysiak, M.; Szulowski, K. Isolation and identification of Pasteurella multocida from European bison in Poland. Post. Mikrobiol. Supl. 2017, 56, 69–70. [Google Scholar]
- Larska, M.; Krzysiak, M.; Smreczak, M.; Polak, M.P.; Zmudziński, J.F. First detection of Schmallenberg virus in elk (Alces alces) indicating infection of wildlife in Białowieża National Park in Poland. Vet. J. 2013, 198, 279–281. [Google Scholar] [CrossRef]
- Koltsov, A.; Tsybanov, S.; Gogin, A.; Kolbasov, D.; Koltsova, G. Identification and characterization of bluetongue virus serotype 14 in Russia. Front. Vet. Sci. 2020, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Kęsik-Maliszewska, J.; Collins, Á.B.; Rola, J.; Blanco-Penedo, I.; Larska, M. Schmallenberg virus in Poland endemic or re-emerging? A six-year serosurvey. Transb. Emerg. Dis. 2021, 68, 2188–2198. [Google Scholar] [CrossRef] [PubMed]
- Larska, M. Schmallenberg virus: A cyclical problem. Vet. Rec. 2018, 183, 688–689. [Google Scholar] [CrossRef] [PubMed]
- Kęsik-Maliszewska, J.; Krzysiak, M.K.; Grochowska, M.; Lechowski, L.; Chase, C.; Larska, M. Epidemiology of Schmallenberg virus in European bison (Bison bonasus) in Poland. J. Wildl. Dis. 2018, 54, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Krzysiak, M.K.; Anusz, K.; Konieczny, A.; Rola, J.; Salat, J.; Strakova, P.; Olech, W.; Larska, M. The European bison (Bison bonasus) as an indicatory species for the circulation of tick-borne encephalitis virus (TBEV) in natural foci in Poland. Ticks Tick-Borne Dis. 2021, 12, 101799. [Google Scholar] [CrossRef]
- Cuéllar, A.C.; Kjær, L.J.; Kirkeby, C.; Skovgard, H.; Nielsen, S.A.; Stockmarr, A.; Andersson, G.; Lindstrom, A.; Chirico, J.; Lühken, R.; et al. Spatial and temporal variation in the abundance of Culicoides biting midges (Diptera: Ceratopogonidae) in nine European countries. Parasites Vectors 2018, 11, 112. [Google Scholar] [CrossRef]
- Filip-Hutsch, K.; Laskowski, Z.; Myczka, A.W.; Czopowicz, M.; Moskwa, B.; Demiaszkiewicz, A.W. The occurrence and molecular identification of Thelazia spp. in European bison (Bison bonasus) in the Bieszczady Mountains. Sci. Rep. 2022, 12, 22508. [Google Scholar] [CrossRef]
- Demiaszkiewicz, A.W.; Moskwa, B.; Gralak, A.; Laskowski, Z.; Myczka, A.W.; Kołodziej-Sobocińska, M.; Kaczor, S.; Plis-Kuprianowicz, E.; Krzysiak, M.; Filip-Hutsch, K. The nematodes Thelazia gulosa Railiet and Henry, 1910 and Thelazia skrjabini Erschov, 1928 as a cause of blindness in European bison (Bison bonasus) in Poland. Acta Parasitol. 2020, 65, 963–968. [Google Scholar] [CrossRef]
- Olech, W.; Perzanowski, K. (Eds.) European Bison (Bison bonasus) Strategic Species Status Review 2020; IUCN SSC Bison Specialist Group and European Bison Conservation Center: Warsaw, Poland, 2022; pp. 1–138. Available online: http://ebcc.wisent.org/science-papers/ (accessed on 3 February 2023).




{kind=link}
{kind=link}
{kind=link}
| Variable | BVDV Seropositives | BoHV-1 Seropositives | BoHV-4 Seropositives | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n/N 1 | % | χ2 | p | n/N 1 | % | χ2 | p | n/N 1 | % | χ2 | p | |
| Herd—level | ||||||||||||
| Origin | 0.14 | 1.0 | 0.1 | 1.0 | 52.2 | 0.0001 | ||||||
| Białowieska Forest | 1/105 | 1.0 | 2/105 | 1.9 | 15/57 | 26.3 | ||||||
| Borecka Forest | 0/64 | 0 | 0/64 | 0 | 0/47 | 0 | ||||||
| Knyszyńska Forest | 2/79 | 2.5 | 0/79 | 0 | 5/60 | 8.3 | ||||||
| Bieszczady | 0/48 | 0 | 0/48 | 0 | 1/22 | 4.5 | ||||||
| Zachodniopomorskie | 0/10 | 0 | 0/10 | 0 | - | - | ||||||
| Augustowska Forest | 0/2 | 0 | 0/2 | 0 | - | - | ||||||
| Bałtów | 0/7 | 0 | 0/7 | 0 | 0/7 | 0 | ||||||
| Białowieża | 0/61 | 0 | 0/61 | 0 | 1/39 | 2.6 | ||||||
| Muczne (Bieszczady) | 0/7 | 0 | 0/7 | 0 | 0/7 | 0 | ||||||
| Gołuchów | 0/15 | 0 | 0/15 | 0 | 0/11 | 0 | ||||||
| Jabłonowo | 0/14 | 0 | 0/14 | 0 | - | - | ||||||
| Kiermusy | 0/8 | 0 | 0/8 | 0 | 0/6 | 0 | ||||||
| Międzyzdroje | 0/7 | 0 | 0/7 | 0 | 0/4 | 0 | ||||||
| Niepołomice | 0/32 | 0 | 0/32 | 0 | 0/26 | 0 | ||||||
| Pszczyna Park | 0/1 | 0 | 0/2 | 0 | 0/2 | 0 | ||||||
| Pszczyna (Jankowice) | 0/96 | 0 | 0/96 | 0 | 2/62 | 3.2 | ||||||
| Smardzewice | 0/29 | 0 | 0/29 | 0 | 0/26 | 0 | ||||||
| Szewce | 0/1 | 0 | 0/1 | 0 | - | - | ||||||
| Ustroń | 0/3 | 0 | 0/3 | 0 | 0/2 | 0 | ||||||
| ZOO Bydgoszcz | 0/1 | 0 | 0/1 | 0 | - | - | ||||||
| ZOO Gdańsk | 0/3 | 0 | 0/3 | 0 | 0/1 | 0 | ||||||
| ZOO Łódź | 0/4 | 0 | 0/4 | 0 | 0/4 | 0 | ||||||
| ZOO Poznań | 0/2 | 0 | 0/2 | 0 | - | - | ||||||
| ZOO Warszawa | 0/10 | 0 | 0/10 | 0 | 0/7 | 0 | ||||||
| Year | 3.5 | 0.8 | 6.5 | 0.5 | 7.9 | 0.09 | ||||||
| 2016 | 0/63 | 0 | 0/63 | 0 | 856 | 14.3 | ||||||
| 2017 | 1/92 | 1.1 | 0/92 | 0 | 3/85 | 3.5 | ||||||
| 2018 | 1/132 | 0.8 | 0/132 | 0 | 7/127 | 5.5 | ||||||
| 2019 | 0/80 | 0 | 0/80 | 0 | 4/71 | 5.6 | ||||||
| 2020 | 1/73 | 1.4 | 1/74 | 1.4 | 2/51 | 3.9 | ||||||
| 2021 | 0/70 | 0 | 1/70 | 1.4 | - | - | ||||||
| 2022 | 0/92 | 0 | 0/92 | 0 | - | - | ||||||
| 2023 | 0/7 | 0 | 0/7 | 0 | - | - | ||||||
| Population type | 3.0 | 0.09 | 1.9 | 0.2 | 16.2 | <0.001 | ||||||
| free-living | 3/308 | 1.0 | 2/308 | 0.6 | 21/186 | 11.3 | ||||||
| captive | 0/301 | 0 | 0/302 | 0 | 3/204 | 1.5 | ||||||
| Population size—free-living | 0.9 | 0.6 | 7.6 | 0.1 | 8.5 | 0.01 | ||||||
| small (≤125) | 0/67 | 0 | 0/67 | 0 | 0/48 | 0 | ||||||
| medium (144–707) | 2/145 | 0.9 | 0/145 | 0 | 16/110 | 9.1 | ||||||
| large (≥715) | 1/96 | 1.5 | 2/96 | 3.1 | 5/25 | 11.1 | ||||||
| Population size—captive | - | - | - | - | 2.0 | 0.4 | ||||||
| small (≤6) | 0/39 | 0 | 0/40 | 0 | 0/31 | 0 | ||||||
| medium (7–42) | 0/198 | 0 | 0/198 | 0 | 3/124 | 6.2 | ||||||
| large (≥43) | 0/64 | 0 | 0/64 | 0 | 0/49 | 2.4 | ||||||
| Animal—level | ||||||||||||
| Age group | 1.2 | 0.5 | 1.4 | 0.5 | 18.5 | <0.001 | ||||||
| ≤1 year old | 0/110 | 0 | 0/110 | 0 | 0/71 | 0 | ||||||
| 2–3 years old | 1/118 | 0.8 | 0/119 | 0 | 0/82 | 0 | ||||||
| ≥4 years old | 1/331 | 0.3 | 2/332 | 0.6 | 24/212 | 11.3 | ||||||
| undefined | 1/48 | 2.0 | 0/48 | 0 | 0/25 | 0 | ||||||
| Gender | 0.2 | 0.6 | 2.2 | 0.141 | 0.6 | 0.4 | ||||||
| female | 2/306 | 0.6 | 0/306 | 0 | 11/203 | 5.4 | ||||||
| male | 1/282 | 0.3 | 2/283 | 0.7 | 13/176 | 7.4 | ||||||
| undefined | 0/21 | 0 | 0/21 | 0 | 0/11 | 0 | ||||||
| Sanitary status | 9.0 | 0.03 | 6.0 | 0.11 | 21.7 | <0.001 | ||||||
| immobilized (healthy) | 0/384 | 0 | 0/385 | 0 | 10/263 | 3.8 | ||||||
| eliminated | 3/152 | 2.0 | 2/152 | 1.3 | 10/98 | 10.2 | ||||||
| fallen | 0/58 | 0 | 0/58 | 0 | 1/22 | 4.5 | ||||||
| dead in traffic accident | 0/15 | 0 | 0/15 | 0 | 3/7 | 42.8 | ||||||
| Variable | BRV3 Seropositives | BAdV-3 Seropositives | BRSV Seropositives | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n/N 1 | % | χ2 | p | n/N 1 | % | χ2 | p | n/N 1 | % | χ2 | p | |
| Herd—level | ||||||||||||
| Origin | 178.1 | <0.001 | 82.6 | <0.001 | 90.0 | <0.001 | ||||||
| Białowieska Forest | 56/71 | 78.9 | 56/71 | 78.9 | 22/71 | 31.0 | ||||||
| Borecka Forest | 24/49 | 49.0 | 36/49 | 73.5 | 12/49 | 24.5 | ||||||
| Knyszyńska Forest | 44/48 | 91.7 | 42/48 | 87.5 | 3/48 | 6.2 | ||||||
| Bieszczady | 3/30 | 10 | 17/30 | 56.7 | 2/30 | 6.7 | ||||||
| Zachodniopomorskie | - | - | - | - | - | - | - | |||||
| Augustowska Forest | 2/2 | 100 | 1/2 | 50 | 2/2 | 100 | ||||||
| Bałtów | 4/6 | 66.7 | 4/6 | 66.7 | 0/6 | 0 | ||||||
| Białowieża | 7/43 | 16.3 | 22/43 | 51.1 | 2/43 | 4.6 | ||||||
| Muczne (Bieszczady) | 0/7 | 0 | 0/7 | 0 | 0/7 | 0 | ||||||
| Gołuchów | 0/10 | 0 | 2/10 | 20 | 7/10 | 70 | ||||||
| Jabłonowo | 3/9 | 33.3 | 2/9 | 22.2 | 0/9 | 0 | ||||||
| Kiermusy | 4/8 | 50 | 3/8 | 37.5 | 3/8 | 37.5 | ||||||
| Międzyzdroje | 1/6 | 16.7 | 0/6 | 0 | 0/6 | 0 | ||||||
| Niepołomice | 5/16 | 31.2 | 6/16 | 37.5 | 0/16 | 0 | ||||||
| Pszczyna Park | 0/2 | 0 | 1/2 | 50 | 0/2 | 0 | ||||||
| Pszczyna (Jankowice) | 1/43 | 2.3 | 23/43 | 53.5 | 0/43 | 0 | ||||||
| Smardzewice | 7/9 | 77.8 | 1/9 | 11.1 | 2/9 | 22.2 | ||||||
| Szewce | 1/1 | 100 | 0/1 | 0 | 1/1 | 100 | ||||||
| Ustroń | 0/3 | 0 | 1/3 | 33.3 | 0/3 | 0 | ||||||
| ZOO Bydgoszcz | 0/1 | 0 | 1/1 | 100 | 0/1 | 0 | ||||||
| ZOO Gdańsk | 2/3 | 66.6 | 1/3 | 33.3 | 0/3 | 0 | ||||||
| ZOO Łódź | 0/1 | 0 | 0/1 | 0 | 1/1 | 100 | ||||||
| ZOO Poznań | - | - | - | - | - | - | ||||||
| ZOO Warszawa | 0/10 | 0 | 6/10 | 60.0 | 3/10 | 15.3 | ||||||
| Year | 0.8 | 0.9 | 6.74 | 0.2 | 4.4 | 0.4 | ||||||
| 2016 | - | - | - | - | - | - | ||||||
| 2017 | - | - | - | - | - | - | ||||||
| 2018 | 57/129 | 44.2 | 82/129 | 63.6 | 18/129 | 14.0 | ||||||
| 2019 | 35/80 | 43.7 | 42/80 | 52.5 | 13/80 | 16.2 | ||||||
| 2020 | 29/74 | 39.2 | 50/74 | 67.6 | 14/74 | 19.0 | ||||||
| 2021 | 31/70 | 44.3 | 36/70 | 51.4 | 14/70 | 20.0 | ||||||
| 2022 | 12/25 | 48.0 | 15/25 | 60.0 | 1/24 | 4.0 | ||||||
| 2023 | - | - | - | - | - | - | ||||||
| Population type | 77.1 | <0.001 | 47.9 | <0.001 | 6.8 | <0.001 | ||||||
| free-living | 129/200 | 64.5 | 152/200 | 76.0 | 41/200 | 20.5 | ||||||
| captive | 35/178 | 19.7 | 73/178 | 41.0 | 19/178 | 10.7 | ||||||
| Population size—free-living | 6.3 | 0.04 | 0.49 | 0.8 | 12.3 | 0.002 | ||||||
| small (≤125) | 26/51 | 51.0 | 37/51 | 72.5 | 14/51 | 27.4 | ||||||
| medium (144–707) | 54/82 | 65.8 | 63/82 | 76.8 | 7/82 | 8.5 | ||||||
| large (≥715) | 49/67 | 73.1 | 52/67 | 77.6 | 20/67 | 29.8 | ||||||
| Population size—captive | 10.6 | 0.005 | 5.9 | 0.05 | 25.8 | <0.001 | ||||||
| small (≤6) | 8/36 | 22.2 | 10/36 | 27.8 | 12/36 | 33.3 | ||||||
| medium (7–42) | 26/100 | 26.0 | 40/100 | 40.0 | 7/100 | 7.0 | ||||||
| large (≥43) | 1/42 | 2.3 | 23/42 | 54.8 | 0/42 | 0 | ||||||
| Animal—level | ||||||||||||
| Age group | 70.6 | <0.001 | 49.8 | <0.001 | 19.3 | <0.001 | ||||||
| ≤1 year old | 5/61 | 8.2 | 30/61 | 49.2 | 2/61 | 3.2 | ||||||
| 2–3 years old | 22/80 | 27.5 | 25/80 | 31.2 | 7/80 | 8.8 | ||||||
| ≥4 years old | 129/204 | 63.2 | 153/204 | 75.0 | 49/204 | 24.0 | ||||||
| undefined | 8/33 | 24.2 | 17/33 | 51.5 | 2/33 | 6.1 | ||||||
| Gender | 0.02 | 0.9 | 0.006 | 0.9 | 0.0007 | 1.0 | ||||||
| female | 88/201 | 43.8 | 120/201 | 59.7 | 32/201 | 15.9 | ||||||
| male | 76/177 | 42.9 | 105/177 | 59.3 | 28/177 | 15.8 | ||||||
| Sanitary status | 41.6 | <0.001 | 20.7 | <0.001 | 6.3 | 0.1 | ||||||
| immobilized (healthy) | 76/242 | 31.4 | 127/242 | 52.5 | 31/242 | 12.8 | ||||||
| eliminated | 58/85 | 68.2 | 67/85 | 78.8 | 20/85 | 23.5 | ||||||
| fallen | 22/40 | 55.0 | 22/40 | 55.0 | 8/40 | 20.0 | ||||||
| dead in traffic accident | 8/11 | 72.7 | 9/11 | 81.8 | 1/11 | 9.1 | ||||||
| Variable | BTV Seropositives | SBV Seropositives | ||||||
|---|---|---|---|---|---|---|---|---|
| n/N 1 | % | χ2 | p | n/N 1 | % | χ2 | p | |
| Herd—level | ||||||||
| Origin | 220.0 | <0.001 | 79.4 | <0.001 | ||||
| Białowieska Forest | 69/105 | 65.7 | 75/102 | 73.5 | ||||
| Borecka Forest | 10/64 | 16.1 | 36/62 | 58.1 | ||||
| Knyszyńska Forest | 37/79 | 46.1 | 63/78 | 80.8 | ||||
| Bieszczady | 1/48 | 2.1 | 40/47 | 85.1 | ||||
| Zachodniopomorskie | 0/9 | 0 | 8/9 | 87.5 | ||||
| Augustowska Forest | 0/2 | 0 | 2/2 | 100 | ||||
| Bałtów | 0/7 | 0 | 7/7 | 100 | ||||
| Białowieża | 14/61 | 23.0 | 40/61 | 65.6 | ||||
| Muczne (Bieszczady) | 0/7 | 0 | 0/7 | 0 | ||||
| Gołuchów | 0/15 | 0 | 7/15 | 46.7 | ||||
| Jabłonowo | 0/14 | 0 | 5/14 | 35.7 | ||||
| Kiermusy | 1/8 | 12.5 | 5/8 | 62.5 | ||||
| Międzyzdroje | 0/7 | 0 | 1/7 | 14.3 | ||||
| Niepołomice | 0/32 | 0 | 21/32 | 65.6 | ||||
| Pszczyna Park | 0/2 | 0 | 2/2 | 100 | ||||
| Pszczyna (Jankowice) | 2/96 | 2.1 | 68/94 | 72.3 | ||||
| Smardzewice | 0/29 | 0 | 28/29 | 96.5 | ||||
| Szewce | 0/1 | 0 | 1/1 | 100 | ||||
| Ustroń | 0/3 | 0 | 3/3 | 100 | ||||
| ZOO Bydgoszcz | 0/1 | 0 | 1/1 | 100 | ||||
| ZOO Gdańsk | 0/3 | 0 | 2/3 | 33.3 | ||||
| ZOO Łódź | 1/4 | 25.0 | 3/4 | 25.0 | ||||
| ZOO Poznań | 0/2 | 0 | 0/2 | 0 | ||||
| ZOO Warszawa | 0/10 | 0 | 5/10 | 16.6 | ||||
| Year | 12.7 | 0.08 | 43.5 | <0.001 | ||||
| 2016 | 21/63 | 33.3 | 47/63 | 74.6 | ||||
| 2017 | 19/92 | 21.0 | 75/91 | 82.4 | ||||
| 2018 | 22/132 | 16.2 | 98/130 | 75.4 | ||||
| 2019 | 21/80 | 26.9 | 43/78 | 55.1 | ||||
| 2020 | 18/74 | 24.6 | 44/73 | 60.3 | ||||
| 2021 | 19/70 | 27.5 | 35/69 | 50.7 | ||||
| 2022 | 15/91 | 14.7 | 74/89 | 82.9 | ||||
| 2023 | 0/7 | 0 | 7/7 | 100 | ||||
| Population type | 91.2 | <0.001 | 5.0 | 0.02 | ||||
| free-living | 117/307 | 38.1 | 224/300 | 74.6 | ||||
| captive | 18/302 | 6.0 | 199/300 | 66.3 | ||||
| Population size—free-living | 18.6 | <0.001 | 9.4 | 0.009 | ||||
| small (≤125) | 11/67 | 16.4 | 39/65 | 60.0 | ||||
| medium (144–707) | 59/144 | 41.0 | 112/142 | 78.9 | ||||
| large (≥715) | 47/96 | 24.0 | 73/93 | 78.5 | ||||
| Population size—captive | 1.7 | 0.4 | 0.04 | 0.4 | ||||
| small (≤6) | 2/40 | 5.0 | 26/40 | 65.0 | ||||
| medium (7–42) | 10/198 | 5.0 | 131/197 | 66.5 | ||||
| large (≥43) | 6/64 | 9.3 | 42/63 | 66.7 | ||||
| Animal—level | ||||||||
| Age group | 67.7 | <0.001 | 64.2 | <0.001 | ||||
| ≤1 year old | 7/110 | 6.4 2 | 48/110 | 43.6 | ||||
| 2–3 years old | 5/118 | 4.3 | 81/118 | 68.4 | ||||
| ≥4 years old | 116/332 | 34.9 | 269/324 | 83.0 | ||||
| undefined | 7/49 | 14.2 | 26/48 | 54.2 | ||||
| Gender | 1.3 | 0.2 | 0.4 | 0.5 | ||||
| female | 66/306 | 21.6 | 205/299 | 68.6 | ||||
| male | 67/282 | 23.8 | 204/280 | 72.8 | ||||
| undefined | 2/21 | 9.5 | 15/22 | 68.2 | ||||
| Sanitary status | 63.4 | <0.001 | 35.9 | <0.001 | ||||
| immobilized (healthy) | 46/385 | 11.6 | 257/379 | 67.8 | ||||
| eliminated | 60/152 | 39.3 | 130/150 | 86.7 | ||||
| fallen | 23/57 | 41.8 | 26/56 | 45.4 | ||||
| dead in traffic accident | 6/15 | 40.0 | 10/15 | 66.6 | ||||
| Human Population Density (Person/km2) | Seroprevalence (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| BVDV | BoHV-1 | BoHV-4 | BRV3 | BAdV-3 | BRSV | BTV | SBV | |
| 19 1–25 2 | 0.5 | 1.1 | 15.4 | 46.3 | 64.7 | 19.1 | 44.1 | 70.3 |
| 31 3–32 4 | 2.8 | 0 | 3.6 | 89.8 | 85.7 | 4.1 | 39.4 | 84.5 |
| 50 5–51 6 | 0 | 0 | 3.5 | 50.9 | 75.5 | 24.5 | 23.7 | 59.4 |
| χ2 | 3.9 | 1.6 | 9.2 | 28.5 | 8.3 | 8.3 | 9.5 | 11.1 |
| p 7 | 0.1 | 0.4 | 0.01 | <0.001 | 0.02 | 0.02 | 0.009 | 0.004 |
| Degree of Urbanization— DEGURBA (%) | ||||||||
| 22 3–38.5 6 | 0 | 0 | 12.5 | 60.0 | 90.0 | 10.0 | 44.4 | 72.2 |
| 41.7 4–42.9 1 | 2.2 | 0 | 4.5 | 63.2 | 72.1 | 5.9 | 32.2 | 81.0 |
| 50.5 2–57.6 5 | 0.4 | 0.9 | 11.4 | 53.1 | 70.0 | 22.5 | 40.4 | 66.8 |
| χ2 | 2.4 | 1.0 | 2.7 | 2.0 | 1.9 | 9.6 | 2.1 | 6.1 |
| p 7 | 0.3 | 0.6 | 0.3 | 0.3 | 0.4 | 0.008 | 0.3 | 0.05 |
| BRV3 (N = 345) | ||||||
| Variable | Category | Odds ratio (OR) | β(SE) | z | p > |z| | 95%CI |
| Age group | ||||||
| ≤1 year old | reference | |||||
| 2–3 years old | 2.1 | 1.3 | 1.2 | 0.2 | 0.6–7.2 | |
| ≥4 years old | 9.6 | 5.3 | 4.1 | <0.001 | 3.2–28.3 | |
| Random effects | Variance | SEM | 95% CI | |||
| origin | 3.9 | 2.1 | 1.3–11.5 | |||
| BTV (N = 560) | ||||||
| Variable | Category | Odds ratio (OR) | β(SE) | z | p > |z| | 95%CI |
| Year | ||||||
| 2016 | reference | |||||
| 2017 | 0.4 | 0.2 | −1.8 | 0.08 | 0.1–1.1 | |
| 2018 | 0.3 | 0.1 | −2.5 | 0.01 | 0.1–0.8 | |
| 2019 | 1.3 | 0.7 | 0.4 | 0.7 | 0.4–3.9 | |
| 2020 | 0.2 | 0.1 | −2.4 | 0.02 | 0.07–0.8 | |
| 2021 | 0.3 | 0.2 | −2.3 | 0.02 | 0.09–0.8 | |
| 2022 | 0.2 | 0.1 | −2.3 | 0.02 | 0.07–0.8 | |
| Age group | ||||||
| ≤1 year old | reference | |||||
| 2–3 years old | 0.4 | 0.3 | −1.3 | 0.2 | 0.1–1.6 | |
| ≥4 years old | 6.4 | 3.3 | 3.6 | <0.001 | 2.3–17.5 | |
| Sanitary status | ||||||
| immobilized (healthy) | reference | |||||
| eliminated | 2.4 | 0.8 | 2.5 | 0.01 | 1.2–4.6 | |
| fallen | 2.0 | 0.9 | 1.5 | 0.1 | 0.8–4.9 | |
| dead in traffic accident | 1.1 | 0.9 | 0.1 | 0.9 | 0.2–5.4 | |
| Random effects | Variance | SEM | 95% CI | |||
| origin | 4.4 | 2.7 | 1.3–14.4 | |||
| SBV (N = 552) | ||||||
| Variable | Category | Odds ratio (OR) | β(SE) | z | p > |z| | 95%CI |
| Year | ||||||
| 2016 | reference | |||||
| 2017 | 3.1 | 1.5 | 2.4 | 0.02 | 1.2–7.9 | |
| 2018 | 1.5 | 0.6 | 1.0 | 0.3 | 0.7–3.3 | |
| 2019 | 0.5 | 0.2 | −1.4 | 0.1 | 0.2–1.2 | |
| 2020 | 0.8 | 0.4 | −0.5 | 0.6 | 0.3–2.0 | |
| 2021 | 0.6 | 0.2 | −1.3 | 0.2 | 0.2–1.3 | |
| 2022 | 3.3 | 1.7 | 2.3 | 0.02 | 1.2–8.8 | |
| Age group | ||||||
| ≤1 year old | reference | |||||
| 2–3 years old | 3.2 | 1.1 | 3.7 | <0.001 | 1.7–6.1 | |
| ≥4 years old | 8.0 | 2.5 | 6.7 | <0.001 | 4.3–14.7 | |
| Random effects | Variance | SEM | 95% CI | |||
| origin | 0.2 | 0.2 | 0.02–1.9 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larska, M.; Tomana, J.; Socha, W.; Rola, J.; Kubiś, P.; Olech, W.; Krzysiak, M.K. Learn the Past and Present to Teach the Future—Role of Active Surveillance of Exposure to Endemic and Emerging Viruses in the Approach of European Bison Health Protection. Diversity 2023, 15, 535. https://doi.org/10.3390/d15040535
Larska M, Tomana J, Socha W, Rola J, Kubiś P, Olech W, Krzysiak MK. Learn the Past and Present to Teach the Future—Role of Active Surveillance of Exposure to Endemic and Emerging Viruses in the Approach of European Bison Health Protection. Diversity. 2023; 15(4):535. https://doi.org/10.3390/d15040535
Chicago/Turabian StyleLarska, Magdalena, Jarosław Tomana, Wojciech Socha, Jerzy Rola, Piotr Kubiś, Wanda Olech, and Michał K. Krzysiak. 2023. "Learn the Past and Present to Teach the Future—Role of Active Surveillance of Exposure to Endemic and Emerging Viruses in the Approach of European Bison Health Protection" Diversity 15, no. 4: 535. https://doi.org/10.3390/d15040535