
Spatiotemporal Distributions of Scolytinae Beetles in the Subtropical Forests of Southern China
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
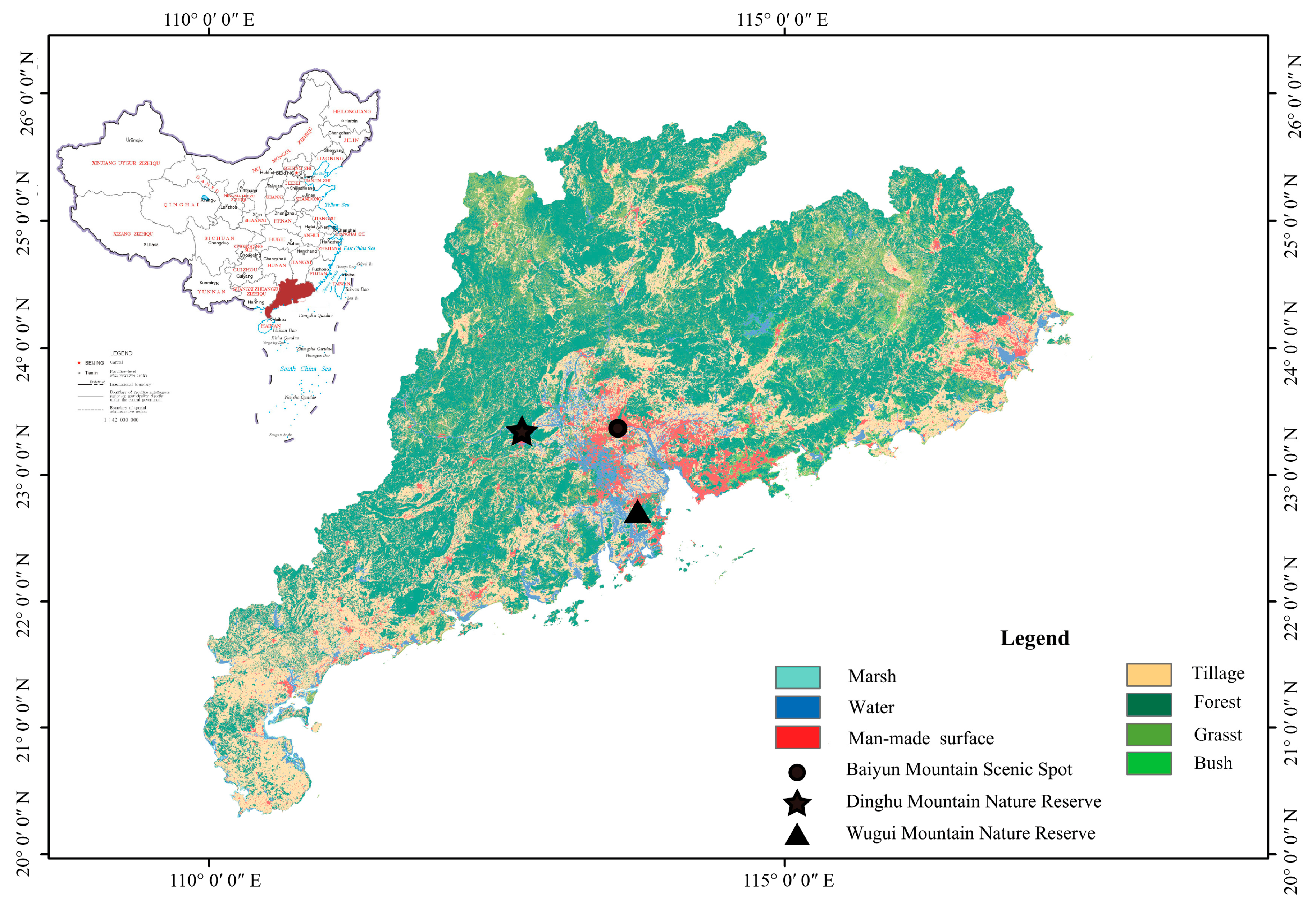
2.1. Study Areas
2.2. Scolytinae Sampling
2.3. Climatic Variables and Vegetation Conditions
2.4. Data Analyses
2.5. Abbreviations
3. Results
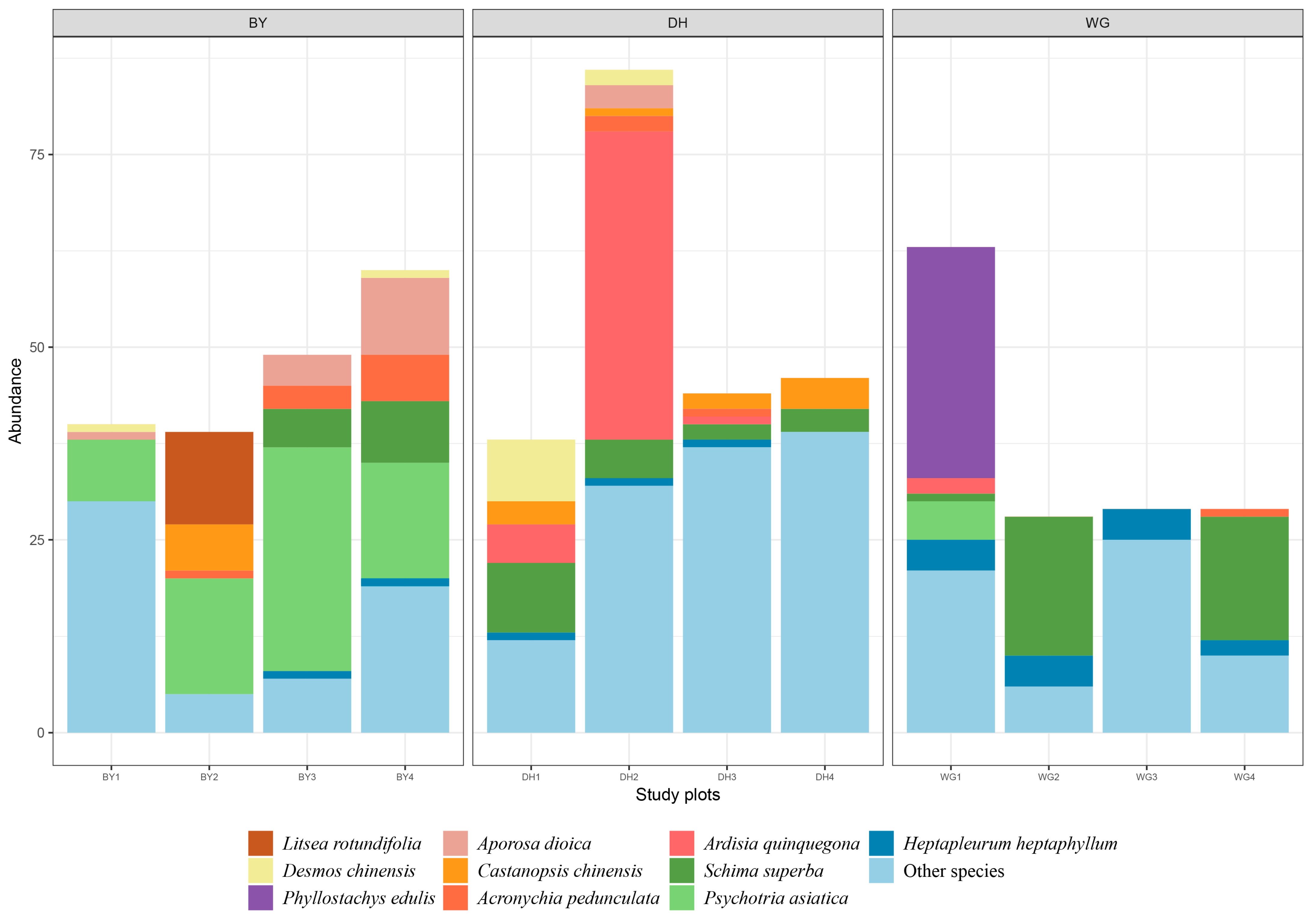
3.1. General Vegetation Conditions in the Three Study Areas
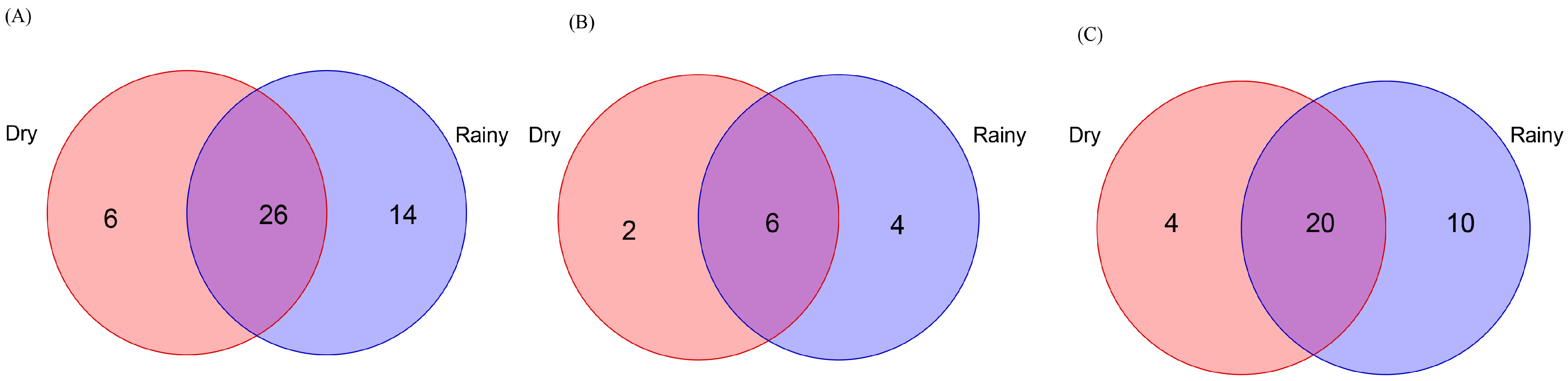
3.2. Endemic and Shared Trees
3.3. Host Tree Diversity Indices
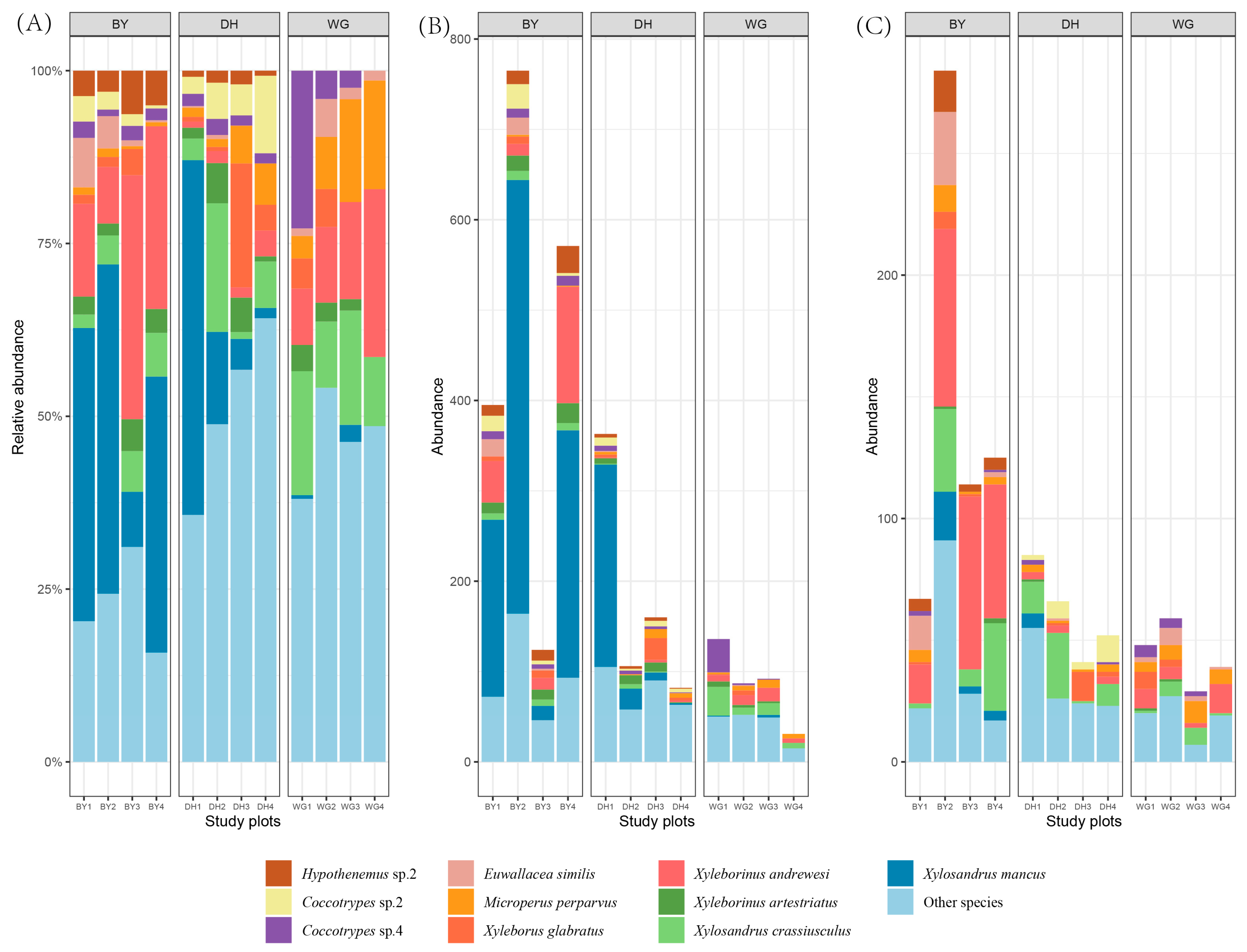
3.4. Scolytinae Beetle Community Composition in the Three Study Areas
3.5. Endemic and Shared Scolytinae Beetles
3.6. Scolytinae Beetle Diversity Indices
3.7. Indicator Species
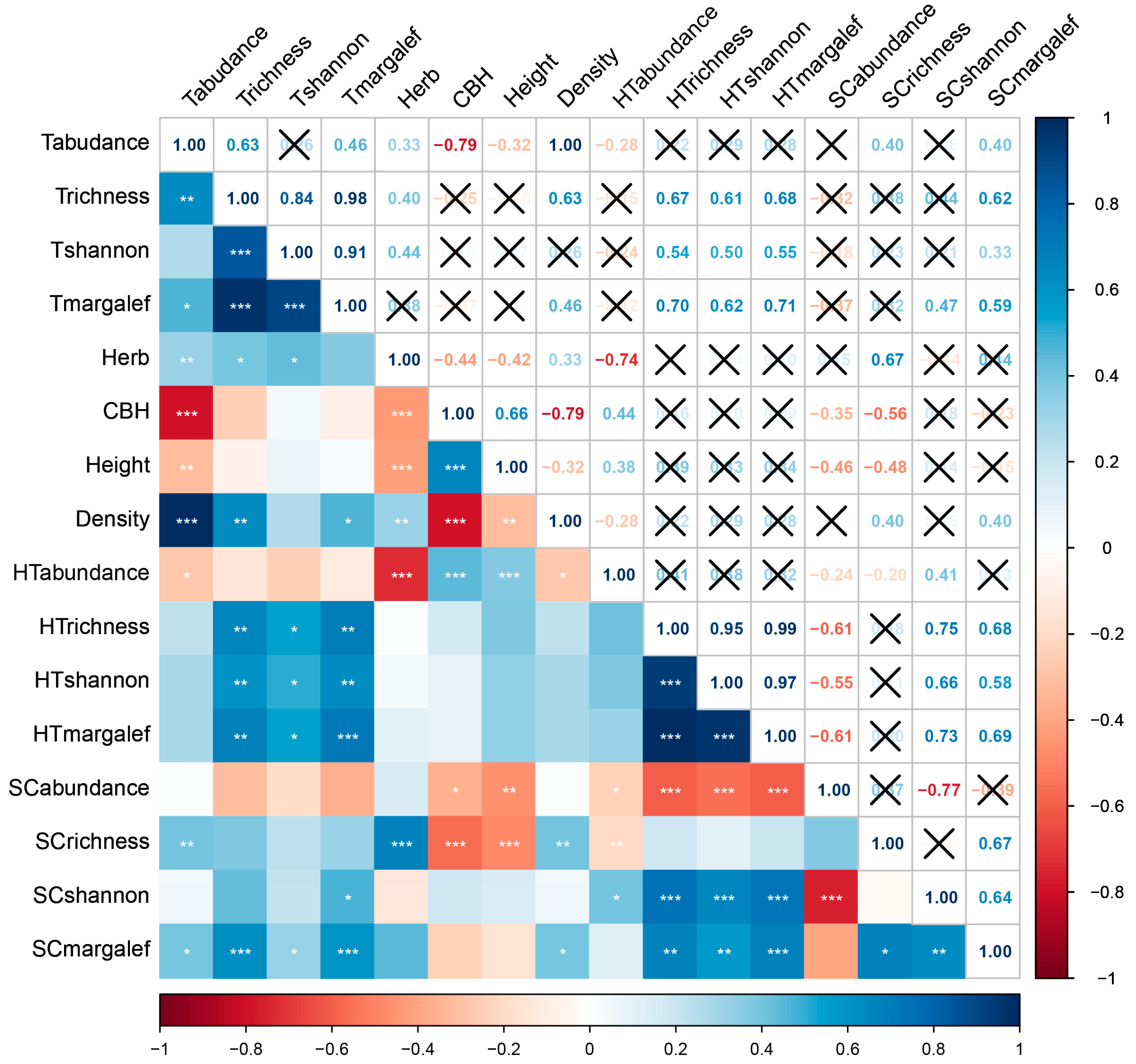
3.8. Relationships between Plants and Scolytinae Beetles
3.9. Associations between the Abundance of Dominant Plants and Dominant Scolytinae Beetles
3.10. Relationships between Climatic Factors and Scolytinae Beetle Abundance
4. Discussion
4.1. Tree Composition Primarily Affects Bark Beetle Community Composition
4.2. Environmental Disturbances Reduce Scolytinae Beetle Diversity
4.3. Scolytinae Beetle Diversity Exhibits Seasonal Variability
4.4. Indicator Species Associated with Environmental Disturbances
4.5. Plants Exhibit a Complex Relationship with Scolytinae Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulcr, J.; Atkinson, T.H.; Cognato, A.I.; Jordal, B.H.; McKenna, D.D. Morphology, Taxonomy and Phylogenetics of Bark Beetles. Bark Beetles. In Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 41–84. [Google Scholar]
- Dole, S.A.; Hulcr, J.; Cognato, A.I. Species-rich bark and ambrosia beetle fauna (Coleoptera, Curculionidae, Scolytinae) of the ecuadorian amazonian forest canopy. ZooKeys 2021, 1044, 797–813. [Google Scholar] [CrossRef] [PubMed]
- Pistone, D.; Gohli, J.; Jordal, B.H. Molecular phylogeny of bark and ambrosia beetles (Curculionidae: Scolytinae) based on 18 molecular markers. Syst. Entomol. 2018, 43, 387–406. [Google Scholar] [CrossRef]
- Kirkendall, L.; Biedermann, P.H.W.; Jordal, B.H. Evolution and diversity of bark and ambrosia beetles. Bark Beetles. In Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 85–156. [Google Scholar]
- Wermelinger, B.; Flückiger, P.F.; Obrist, M.K.; Duelli, P. Horizontal and vertical distribution of saproxylic beetles (Col., Buprestidae, Cerambycidae, Scolytinae) across sections of forest edges. J. Appl. Entomol. 2007, 131, 104–114. [Google Scholar] [CrossRef]
- Hulcr, J.; Beaver, R.A.; Puranasakul, W.; Dole, S.A.; Sonthichai, S. A comparison of bark and ambrosia beetle communities in two forest types in Northern Thailand (Coleoptera: Curculionidae: Scolytinae and Platypodinae). Environ. Entomol. 2008, 37, 1461–1470. [Google Scholar] [CrossRef]
- Kalapanida-Kantartzi, M.; Milonas, D.N.; Buchelos, C.T.; Avtsiz, D.N. How does pollution affect insect diversity? A study on bark beetle entomofauna of two pine forests in Greece. J. Biol. Res. 2010, 13, 67–74. [Google Scholar]
- Martínez, M.; Cognato, A.I.; Guachambala, M.; Boivin, T. Bark and ambrosia beetle (Coleoptera: Curculionidae: Scolytinae) diversity in natural and plantation forests in Ecuador. Environ. Entomol. 2019, 48, 603–613. [Google Scholar] [CrossRef]
- Sanguansub, S.; Buranapanichpan, S.; Beaver, R.A.; Saowaphak, T.; Tanaka, N.; Kamata, N. Influence of seasonality and climate on captures of wood-boring Coleoptera (Bostrichidae and Curculionidae (Scolytinae and Platypodinae)) using ethanol-baited traps in a seasonal tropical forest of northern Thailand. J. Forest Res. 2020, 25, 223–231. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Rassati, D. Effect of trap color on captures of bark- and wood-boring beetles (Coleoptera; Buprestidae and Scolytinae) and associated predators. Insects 2020, 11, 749. [Google Scholar] [CrossRef]
- Huang, F.S.; Lu, J. The Classification Outline of Scolytidae from China; Tongji University Press: Shanghai, China, 2015. [Google Scholar]
- Johnson, A.J.; Li, Y.; Mandelshtam, M.Y.; Park, S.; Lin, S.C.; Gao, L.; Hulcr, J. East Asian Cryphalus Erichson (Curculionidae, Scolytinae): New species, new synonymy and redescriptions of species. ZooKeys 2020, 995, 15–66. [Google Scholar] [CrossRef]
- Smith, S.M.; Beaver, R.A.; Cognato, A.I. A monograph of the Xyleborini (Coleoptera, Curculionidae, Scolytinae) of the Indochinese Peninsula (except Malaysia) and China. Zookeys 2020, 983, 1–442. [Google Scholar] [CrossRef]
- Cognato, A.I.; Smith, S.M.; Li, Y.; Pham, T.H.; Hulcr, J. Genetic variability among Xyleborus glabratus populations native to Southeast Asia (Coleoptera: Curculionidae: Scolytinae: Xyleborini) and the description of two related species. J. Econom. Entomol. 2019, 112, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Xu, M.F.; Gao, L.; Ruan, Y.Y.; Lai, S.C.; Xu, Y.; Li, Y. New records of two invasive ambrosia beetles (Curculionidae: Scolytinae: Xyleborini) to mainland China. BioInvas. Records 2021, 10, 74–80. [Google Scholar] [CrossRef]
- Yin, H.F.; Huang, F.S.; Li, Z.L. Economic Insect Fauna of China. Fasc. 29. Coleoptera: Scolytidae; Science Press: Beijing, China, 1984. [Google Scholar]
- Brockerhoffff, E.G.; Liebhold, A.M. Ecology of forest insect invasions. Biol. Invas. 2017, 19, 3141–3159. [Google Scholar] [CrossRef]
- Kirkendall, L.R.; Faccoli, M. Bark beetles and pinhole borers (Curculionidae, Scolytinae, Platypodinae) alien to Europe. Zookeys 2010, 56, 227–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haack, R.A.; Rabaglia, R.J. Non-native bark and ambrosia beetles in the USA: Potential and current invaders. In Potential Invasive Pests of Agricultural Crops; Peña, J.E., Ed.; CAB International: Wallingford, UK, 2013; pp. 48–74. [Google Scholar]
- Brockerhoff, E.G.; Bain, J.; Kimberley, M.O.; Knizek, M. Interception frequency of non-native bark and ambrosia beetles (Coleoptera: Scolytinae) and relationship with establishment in New Zealand and worldwide. Can. J. Forest Res. 2006, 36, 289–298. [Google Scholar] [CrossRef]
- Chang, H.; Liu, Q.; Hao, D.; Liu, Y.; An, Y.; Qian, L.; Yang, X. DNA barcodes and molecular diagnostics for distinguishing introduced Xyleborus (Coleoptera: Scolytinae) species in China. Mito. DNA 2013, 25, 63–69. [Google Scholar] [CrossRef]
- Zhou, G.Y.; Wei, X.H.; Wu, Y.P.; Liu, S.G.; Huang, Y.H.; Yan, J.H.; Zhang, D.Q.; Zhang, Q.M.; Liu, J.X.; Meng, Z.; et al. Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China. Glob. Chang. Biol. 2011, 17, 3736–3746. [Google Scholar] [CrossRef]
- Qiao, Y.; Xiong, Y.M.; Wu, Q.H. Structure and diversity of soil macrofauna and mesofauna communities in the Baiyun Mountain, Guangzhou. Guangdong Landsc. Archit. 2018, 5, 17–20. [Google Scholar]
- Yang, G.; Liao, K.Y. Analysis on the development law of couapse and landslide disaster in zhongshan city. Quat. Sci. 2019, 39, 1246–1251. [Google Scholar]
- Liu, P.L.; Chen, L.; Liu, X.D.; Dai, Y.H.; Feng, Y.J.; Zhang, Q.M.; Chu, G.W.; Meng, Z. Temporal and spatial variability of soil moisture in a forest succession series in Dinghushan. Acta Ecol. Sin. 2021, 41, 1798–1807. [Google Scholar]
- Zhang, J.Q. The Characteristics of the Vegetation and It’s Stand Reforms in Bai Yun Mountain Scenic Spots and Historical Sites, Guangzhou. Ecol. Sci. 1995, 1, 31–39. [Google Scholar]
- Xiao, J. Present situation of Natural forest and ecological problem in Wugui Mountain. Trop. Forest. 1985, 4, 25–29. [Google Scholar]
- Wang, Y.H.; Chen, B.G.; Yao, S.Z. Vegetation landscape types and characteristics of Baiyunshan Scenic Spot, Guangzhou. J. South China Agric. Univ. 2003, 24, 56–59. [Google Scholar]
- Xu, C.Y.; Feng, Z.J.; Li, Z.K. An investigation of vascular plant resources at the Recreation Landscape Greenbelt on Western Side of Baiyun Mountain, Guangzhou. J. South China Agric. Univ. 2002, 23, 56–59. [Google Scholar]
- Wang, J.B.; Tan, Z.J.; Jiang, Q.C.; Gu, J.M.; Liu, C.K.; Zhang, Y.S. A catalogue of insects investigation from Wugui Mountain of Zhongshan Nature Reserve (I). Modern Agr. Sci. Technol. 2011, 13, 304–306. [Google Scholar]
- Wood, S.L. A reclassification of the genera of Scolytidae (Coleoptera). Great Basin Nat. Mem. 1986, 10, 1–126. [Google Scholar]
- Ugland, K.I.; Gray, J.S.; Ellingsen, K.E. The species-accumulation curve and estimation of species richness. J. Animal Ecol. 2003, 72, 888–897. [Google Scholar] [CrossRef] [Green Version]
- Li, Q. Species accumulation curves and its application. Chin. J. Appl. Entomol. 2011, 48, 1882–1888. [Google Scholar]
- Magurran, E.A. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004. [Google Scholar]
- Jaccard, P. The distribution of the flora in the alpine zone.1. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monographs 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Hulcr, J.; Mogia, M.; Isua, B.; Novotny, V. Host specifificity of ambrosia and bark beetles (Col., Curculionidae: Scolytinae and Platypodinae) in a New Guinea rainforest. Ecol. Entomol. 2007, 32, 762–772. [Google Scholar] [CrossRef]
- Raffa, K.F.; Grégoire, J.C.; Lindgren, B.S. Natural History and Ecology of Bark Beetles. Bark Beetles. In Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 1–40. [Google Scholar]
- Tanner, L.D.; Kanga, L.H.; Haseeb, M.; Whilby, L.; Onokpise, O.U. Efficacy of selected attractants for monitoring the populations of the redbay ambrosia beetle, Xyleborus glabratus Eichhoff (Coleoptera: Scolytidae) and other bark beetles in the Florida Panhandle. Curr. Investig. Agr. Curr. Res. 2018, 1, 1–7. [Google Scholar]
- Cognato, A.I.; Sari, G.; Smith, S.; Beaver, R.A.; Li, Y.; Hulcr, J.; Jordal, B.H.; Kajimura, H.; Lin, C.S.; Pham, T.H.; et al. The essential role of taxonomic expertise in the creation of DNA databases for the identifification and delimitation of Southeast Asian ambrosia beetle species (Curculionidae: Scolytinae: Xyleborini). Front. Ecol. Evol. 2020, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Beaver, R.A. Host specifificity of temperate and tropical animals. Nature 1979, 281, 139–141. [Google Scholar] [CrossRef]
- Abreu, R.L.S.; de Ribeiro, G.A.; Vianez, B.F.; Sales-Campos, C. Insects of the subfamily Scolytinae (Insecta: Coleoptera, Curculionidae) collected with pitfall and ethanol traps in primary forests of central Amazonia. Psyche 2015, 2012, 480520. [Google Scholar]
- Macedo-Reis, L.E.; de Novais, S.M.A.; Monteiro, G.F.; Flechtmann, C.A.H.; de Faria, M.L.; de Siqueira Neves, F. Spatio-Temporal distribution of bark and ambrosia beetles in a Brazilian tropical dry forest. J. Insect Sci. 2016, 16, 48. [Google Scholar] [CrossRef] [Green Version]
- Sittichaya, W.; Permkam, S.; Cognato, A.I. Species composition and flight pattern of Xyleborini ambrosia beetles (Col.: Curculionidae: Scolytinae) from agricultural areas in southern Thailand. Environ. Entomol. 2012, 41, 776–784. [Google Scholar] [CrossRef]
- Freedman, B. Environmental ecology: The impact of pollution and other stresses on ecosystem structure and function. Ecol. Econom. 1989, 4, 168–169. [Google Scholar]
- Perry, J.; Lojka, B.; Ruiz, L.G.Q.; Damme, P.V.; Houška, J.; Cusimamani, F.E. How natural forest conversion affects insect biodiversity in the Peruvian Amazon: Can agroforestry help? Forests 2016, 7, 82. [Google Scholar] [CrossRef] [Green Version]
- Noriega, J.A.; March-Salas, M.; Castillo, S.; García-Q, H.; Hortal1, J.; Santos, A.M.C. Human perturbations reduce dung beetle diversity and dung removal ecosystem function. Biotropica 2021, 3, 753–766. [Google Scholar] [CrossRef]
- Zhao, J.L. Study on Tourism Carrying Capacity Management in Bai-Yun Mountain Scenic Area. Master’s Thesis, Guangzhou University, Guangzhou, China, 2011. [Google Scholar]
- Senzaki, M.; Kadoya, T.; Francis, C.D. Direct and indirect effects of noise pollution alter biological communities in and near noise-exposed environments. Proc. R. Soc. B 2020, 287, 20200176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.S.; Fang, Y.; Li, K. Impacts of urbanization process on insect diversity. Biodivers. Sci. 2013, 21, 260–268. [Google Scholar]
- Renwick, J.A.A. Nonpreference mechanisms: Plant characteristics influencing insect behavior. In Plant Resistance to Insects: A Fundamental Approach; Smith, C.M., Ed.; Wiley-Interscience: New York, NY, USA, 1983; pp. 199–213. [Google Scholar]
- Peng, Y.; Zhou, K.; He, B.H.; Wang, J.L.; Wei, H. Analysis on Dynamic Changes of Forest Landscape in Different Zones of Dinghu Mountain Nature Reserve. For. Resour. Manag. 2011, 3, 76–81. [Google Scholar]
- Kuang, Y.W.; Wen, Z.D.; Zhou, G.Y.; Li, Z.A. Light atmospheric Nonmethane Hydrocarbons at areas around Guangzhou, China. J. Agro-Environ. Sci. 2003, 22, 570–573. [Google Scholar]
- Ouyang, X.J.; Zhou, G.Y.; Huang, Z.L.; Huang, M.H. Analysis on Runoff Water Quality in Dinghushan Biosphere Reserve. Acta Ecol. Sin. 2002, 22, 1373–1379. [Google Scholar]
- Noriega, J.A.; Zapata-Prisco, C.; García, H.; Hernández, E.; Hernández, J.; Martínez, R.; Santos-Santos, J.R.; Pablo-Cea, J.D.; Calatayud, J. Does ecotourism impact biodiversity? An assessment using dung beetles (Coleoptera: Scarabaeinae) as bioindicators in a tropical dry forest natural park. Ecol. Indic. 2020, 117, 106580. [Google Scholar] [CrossRef]
- Hlásny, T.; Krokene, P.; Liebhold, A.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.; Schelhaas, M.J.; Seidl, R.; Svoboda, M.; et al. Living with Bark Beetles: Impacts, Outlook and Management Options; European Forest Institute: Joensuu, Finland, 2019; Volume 8. [Google Scholar] [CrossRef]
- Wollmann, J.; Garcia, M.S.; Flechtmann, C.A.H.; Finkenauer, E.; Garcia, F.R.M. Scolytinae assemblage structure (Coleoptera: Curculionidae) in forested areas with Eucalyptus spp. in southern Rio Grande do Sul state. Cien. Florest. 2007, 27, 1167–1177. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.A.; Andreiv, J. Caracterização da família Scolytidae (Insecta: Coleoptera) em três ambientes florestais. Cerne 2004, 10, 39–45. [Google Scholar]
- Ulyshen, M.D.; Hanula, J.L.; Horn, S.; Kilgo, J.C.; Moorman, C.E. Spatial and temporal patterns of beetles associated with coarse woody debris in managed bottomland hardwood forests. Forest Ecol. Manag. 2004, 199, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.Y.; Song, B.; Zhao, S.; Ding, S.Y. Effect of agro-landscape heterogeneity as affected by scale on diversity of Coleoptera in Fengqiu county in the lower reaches of the Yellow River. J. Ecol. Rural Environ. 2015, 31, 77–81. [Google Scholar]
- Grove, S.J. Saproxylic insect ecology and the sustainable management of forests. Ann. Rev. Ecol. Syst. 2002, 33, 1–23. [Google Scholar] [CrossRef]
- Lv, J.; Lai, S.C.; Tian, S.; Zhou, Q.; Xiao, L.F.; He, P.S.; Wang, J.G. Taxonomic studies on the Xyleborini beetles from Jiangxi Province, China. J. Environ. Entomol. 2018, 40, 840–852. [Google Scholar]
- Spennemann, D.H.R. Biology, ecology and distribution of the Date Stone Beetle, Coccotrypes dactyliperda (Coleoptera: Curculionidae). Zool. Middle East 2019, 65, 163–182. [Google Scholar] [CrossRef]
- Zhang, L. Taxonomy and Phylogeny of Xyleborini in Fujian Province; Jiangxi Agricultural University: Nangchang, China, 2021. [Google Scholar]
- Lv, J. Taxonomy and Phylogeny of Xyleborini (Coleoptera: Scolytinae) Beetles in Jiangxi Province. Master’s Thesis, Jiangxi Agricultural University, Nangchang, China, 2018. [Google Scholar]
- Zhou, Q. Species Investigation and DNA Barcoding of Rubberwood-Destroying Beetles of Scolytinae in Yunnan Province. Master’s Thesis, Jiangxi Agricultural University, Nangchang, China, 2020. [Google Scholar]
- Tian, S. Biodiversity and Molecular Identification of the Scolytinae Reinvestigation in Chongqing Area. Master’s Thesis, Jiangxi Agricultural University, Nangchang, China, 2018. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Vegetation Conditions | BY | DH | WG |
|---|---|---|---|
| Abundance of trees | 47 ± 8.46 a | 53.5 ± 18.99 a | 37.25 ± 14.87 a |
| Richness of trees | 10.75 ± 3.49 a | 18.75 ± 4.60 a | 11 ± 3.54 a |
| Shannon–Wiener diversity index of trees | 1.84 ± 0.39 a | 2.48 ± 0.29 a | 1.78 ± 0.46 a |
| Margalef richness index of trees | 2.52 ± 0.85 a | 4.49 ± 1.02 a | 2.78 ± 0.86 a |
| Average CBH of trees | 18.91 ± 1.80 a | 21.14 ± 6.32 a | 28.06 ± 6.09 a |
| Average height of trees | 4.72 ± 0.47 a | 5.43 ± 0.98 a | 7.76 ± 1.22 a |
| Average coverage of herbaceous layer | 0.67 ± 0.21 ab | 0.77 ± 0.13 a | 0.29 ± 0.19 b |
| Average density of trees | 0.50 ± 0.08 a | 0.54 ± 0.19 a | 0.37 ± 0.15 a |
| Species | BY | DH | WG | Total | % | Habitat Preference (IndVal) | p |
|---|---|---|---|---|---|---|---|
| Phloeosinus sp. | 0 | 0 | 1 | 1 | 0.03 | / | / |
| Cryphalus dilutus | 9 | 0 | 0 | 9 | 0.23 | / | / |
| Cryphalus itinerans | 24 | 1 | 15 | 40 | 1.02 | / | / |
| Cryphalus meridionalis | 0 | 0 | 1 | 1 | 0.03 | / | / |
| Cryphalus sp. | 1 | 2 | 0 | 3 | 0.08 | / | / |
| Hypothenemus birmanus | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Hypothenemus sp.1 | 0 | 7 | 15 | 22 | 0.56 | / | / |
| Hypothenemus sp.2 | 99 | 12 | 0 | 111 | 2.83 | BY(0.89) | 0.003 |
| Coriacephilus sp. | 1 | 14 | 18 | 33 | 0.84 | / | / |
| Eidophelus fulvipennis | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Eidophelus sp.1 | 1 | 0 | 2 | 3 | 0.08 | / | / |
| Ernoporini sp. | 0 | 2 | 0 | 2 | 0.05 | / | / |
| Cyrtogenius luteus | 0 | 1 | 1 | 2 | 0.05 | / | / |
| Coccotrypes sp.1 | 31 | 36 | 0 | 67 | 1.71 | / | / |
| Coccotrypes sp.2 | 51 | 44 | 0 | 95 | 2.42 | / | / |
| Coccotrypes sp.3 | 10 | 10 | 6 | 26 | 0.66 | / | / |
| Coccotrypes sp.4 | 38 | 17 | 51 | 106 | 2.70 | / | / |
| Coccotrypes sp.5 | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Coccotrypes sp.6 | 3 | 0 | 5 | 8 | 0.20 | / | / |
| Coccotrypes sp.7 | 0 | 0 | 10 | 10 | 0.26 | WG(1.00) | 0.007 |
| Dryocoetiops moestus | 9 | 3 | 0 | 12 | 0.31 | BY(0.75) | 0.025 |
| Dryocoetini sp. | 0 | 0 | 4 | 4 | 0.10 | / | / |
| Sueus niisimai | 2 | 23 | 10 | 35 | 0.89 | / | / |
| Amasa schlichii | 1 | 0 | 7 | 8 | 0.20 | / | / |
| Ambrosiodmus asperatus | 7 | 1 | 0 | 8 | 0.20 | BY(0.88) | 0.02 |
| Ambrosiodmus rubricollis | 27 | 7 | 3 | 37 | 0.94 | / | / |
| Ambrosiophilus atratus | 0 | 3 | 0 | 3 | 0.08 | / | / |
| Ambrosiophilus cristatulus | 4 | 3 | 1 | 8 | 0.20 | / | / |
| Ambrosiophilus osumiensis | 3 | 0 | 1 | 4 | 0.10 | / | / |
| Anisandrus ursulus | 0 | 10 | 0 | 10 | 0.26 | / | / |
| Arixyleborus cf. silvanus | 1 | 0 | 0 | 1 | 0.03 | / | / |
| Arixyleborus sp. | 0 | 0 | 1 | 1 | 0.03 | / | / |
| Beaverium magnus | 1 | 1 | 1 | 3 | 0.08 | / | / |
| Cnestus aterrimus | 4 | 1 | 1 | 6 | 0.15 | / | / |
| Cnestus mutilatus | 0 | 2 | 0 | 2 | 0.05 | / | / |
| Cyclorhipidion armiger | 0 | 4 | 0 | 4 | 0.10 | / | / |
| Cyclorhipidion beaveri | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Cyclorhipidion bodoanum | 68 | 16 | 2 | 86 | 2.19 | BY(0.79) | 0.036 |
| Cyclorhipidion distinguendum | 8 | 17 | 0 | 25 | 0.64 | / | / |
| Cyclorhipidion japonicum | 31 | 3 | 0 | 34 | 0.87 | / | / |
| Cyclorhipidion cf. xeniolum | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Diuncus haberkorni | 0 | 7 | 4 | 11 | 0.28 | / | / |
| Diuncus mucronatus | 3 | 4 | 15 | 22 | 0.56 | / | / |
| Euwallacea andamanensis | 2 | 0 | 0 | 2 | 0.05 | / | / |
| Euwallacea fornicatus | 8 | 4 | 14 | 26 | 0.66 | WG(0.75) | 0.049 |
| Euwallacea interjectus | 4 | 1 | 15 | 20 | 0.51 | / | / |
| Euwallacea minutus | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Euwallacea semiermis | 0 | 0 | 1 | 1 | 0.03 | / | / |
| Euwallacea similis | 86 | 2 | 13 | 101 | 2.58 | / | / |
| Euwallacea sp. | 14 | 4 | 0 | 18 | 0.46 | / | / |
| Fraudatrix melas | 0 | 0 | 1 | 1 | 0.03 | / | / |
| Hadrodemius comans | 1 | 10 | 0 | 11 | 0.28 | DH(0.91) | 0.008 |
| Microperus alpha | 18 | 29 | 5 | 52 | 1.33 | / | / |
| Microperus cf. chrysophylli | 1 | 0 | 0 | 1 | 0.03 | / | / |
| Microperus diversicolor | 79 | 1 | 1 | 81 | 2.07 | BY(0.98) | 0.007 |
| Microperus kadoyamaensis | 11 | 62 | 0 | 73 | 1.86 | DH(0.85) | 0.008 |
| Microperus perparvus | 23 | 27 | 46 | 96 | 2.45 | / | / |
| Planiculus bicolor | 41 | 36 | 5 | 82 | 2.09 | / | / |
| Xyleborinus andrewesi | 416 | 15 | 65 | 496 | 12.65 | BY(0.84) | 0.007 |
| Xyleborinus artestriatus | 65 | 28 | 13 | 106 | 2.70 | BY(0.61) | 0.007 |
| Xyleborinus ephialtodes | 0 | 8 | 0 | 8 | 0.20 | / | / |
| Xyleborinus cf. jianghuasuni | 2 | 0 | 0 | 2 | 0.05 | / | / |
| Xyleborinus octiesdentatus | 4 | 0 | 2 | 6 | 0.15 | / | / |
| Xyleborinus saxesenii | 23 | 8 | 20 | 51 | 1.30 | / | / |
| Xyleborus affinis | 39 | 23 | 13 | 75 | 1.91 | / | / |
| Xyleborus festivus | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Xyleborus glabratus | 30 | 45 | 16 | 91 | 2.32 | / | / |
| Xyleborus perforans | 1 | 7 | 6 | 14 | 0.36 | / | / |
| Xylosandrus nr. adherescens | 0 | 1 | 0 | 1 | 0.03 | / | / |
| Xylosandrus amputatus | 0 | 8 | 0 | 8 | 0.20 | / | / |
| Xylosandrus borealis | 1 | 0 | 1 | 2 | 0.05 | / | / |
| Xylosandrus compactus | 18 | 11 | 14 | 43 | 1.10 | / | / |
| Xylosandrus crassiusculus | 111 | 57 | 74 | 242 | 6.17 | / | / |
| Xylosandrus discolor | 17 | 18 | 2 | 37 | 0.94 | / | / |
| Xylosandrus germanus | 0 | 6 | 5 | 11 | 0.28 | / | / |
| Xylosandrus mancus | 993 | 264 | 4 | 1261 | 32.16 | / | / |
| Xyleborini sp. | 0 | 0 | 10 | 10 | 0.26 | / | / |
| Scolytoplatypus sinensis | 0 | 22 | 0 | 22 | 0.56 | DH(1.00) | 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Li, L.; Yang, X.; Lin, W.; Han, Q.; Li, Z. Spatiotemporal Distributions of Scolytinae Beetles in the Subtropical Forests of Southern China. Diversity 2023, 15, 499. https://doi.org/10.3390/d15040499
Yu Y, Li L, Yang X, Lin W, Han Q, Li Z. Spatiotemporal Distributions of Scolytinae Beetles in the Subtropical Forests of Southern China. Diversity. 2023; 15(4):499. https://doi.org/10.3390/d15040499
Chicago/Turabian StyleYu, Yali, Lanfeng Li, Xingke Yang, Wei Lin, Qunxin Han, and Zhiqiang Li. 2023. "Spatiotemporal Distributions of Scolytinae Beetles in the Subtropical Forests of Southern China" Diversity 15, no. 4: 499. https://doi.org/10.3390/d15040499