
Parasites in Imported Edible Fish and a Systematic Review of the Pathophysiology of Infection and the Potential Threat to Australian Native Aquatic Species


Abstract
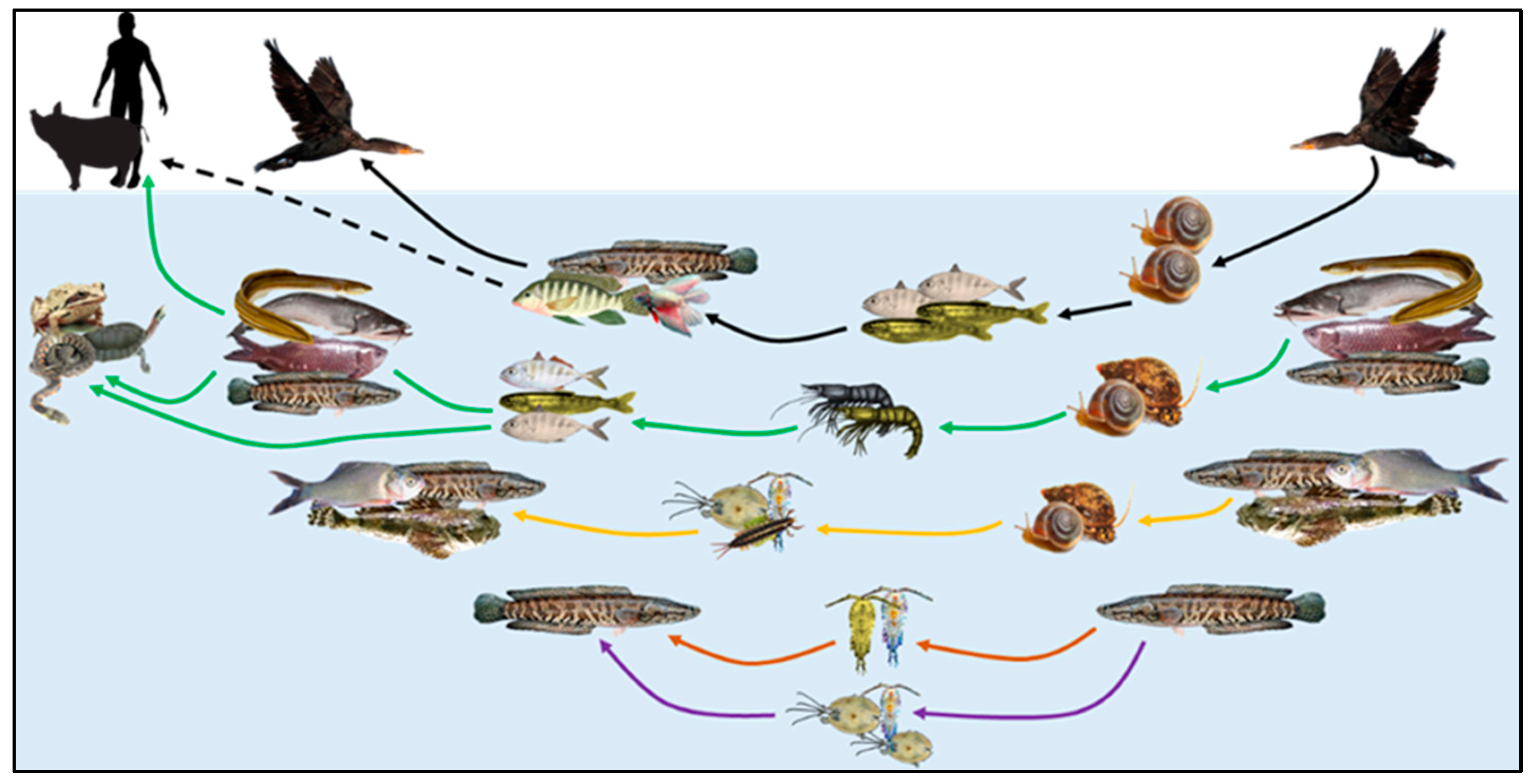
:1. Introduction
2. Materials and Method
2.1. Parasite Collection and Preparation
2.2. Morphological Identification
2.3. Parasite Population Calculations
2.4. Literature Search
3. Results
3.1. Prevalence of Helminths in Fish
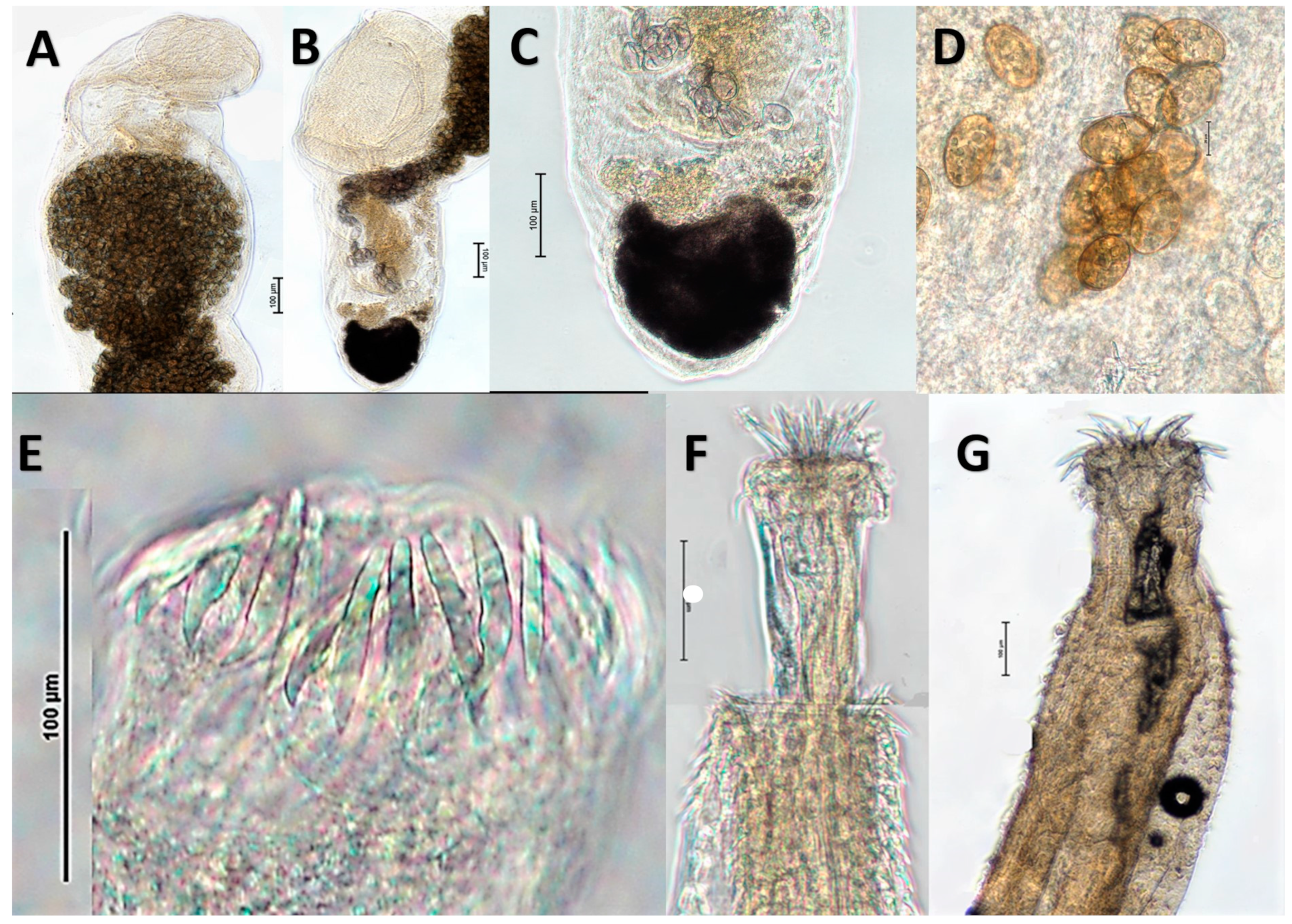
3.2. Morphological Identification of Helminths from Consumer Ready Channa Fish
3.2.1. Genarchopsis Species Ozaki, 1925, Family Derogenidae Nicoll, 1910, Class Trematoda
Genarchopsis Species General Observations
Genarchopsis paithanensis
Genarchopsis sp. 1
3.2.2. Senga sp. Dollfus, 1934, Family Bothriocephalidae Blanchard, 1849, Class Cestoda
3.2.3. Pallisentis Species Van Cleave, 1928, Family Quadrigyridae Van Cleave, 1920, Class Eoacanthocephala
Pallisentis (Brevitritospinus) sp. 1
Pallisentis (Pallisentis) gomptii
3.3. Literature Search Results
3.3.1. Euclinostomum sp., Travassos, 1928, Pathophysiology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species of Parasite | Host Infected | Site of Infection | Geographical Locality | Fish Pathophysiology | Reference |
|---|---|---|---|---|---|
| Euclinostomum ardeolae El-Naffar & Khalifa, 1981 | Nile tilapia, Oreochromis niloticus (Linnaeus, 1758) | Kidney | Egypt | Grey/black cysts | Ahmed, et al. [57] |
| Euclinostomum heterostomum (Rudolphi, 1809) Travassos, 1928 | Spotted snakehead, Channa punctata (Bloch, 1793) | Liver, kidney & viscera | India | Heavily parasitized fish lethargic | Bhargavi, et al. [58] |
| Euclinostomum heterostomum | Mozambique tilapia, Oreochromis mossambicus (Peters, 1852) | High in muscle | Multiple locations, Venda and Lebowa, southern Africa | Loss of consumer confidence. | Britz, et al. [51] |
| Euclinostomum heterostomum | Redbelly tilapia, Tilapia zillii syn. Coptodon zillii (Gervais, 1848) | Body cavity, skin, eye | Opi Lake, Nigeria | Pronounced inflammation & roughened skin. Ex-cysted metacercariae associated damage from burrowing through host organs. Fish blindness, myositis, muscle bumps. Decreased fish marketability. | Echi, et al. [21] |
| Euclinostomum heterostomum | Redbelly tilapia T. zillii syn. Coptodon zillii | Skin | Nigeria | Co-infection with other clinostomatids causes cysts, ulcers, degeneration of skin/muscle, necrosis. | Echi, et al. [59] |
| Euclinostomum heterostomum | Striped snakehead, Channa striata (Bloch, 1793) | Body cavity, muscles, liver, gill opening, intestine, kidneys & ovaries | Bhopal, India | Fish with reduced glomeruli size, severe degeneration/necrosis of hemopoietic tissue and tubule cells with hypertrophied nuclei & epithelial cells detached. Occlusion of tubular lumen. | Kaur, et al. [60] |
| Euclinostomum heterostomum | Spotted snakehead, C. punctata | Liver | Bhopal, India | Degeneration & necrosis of liver tissue with enucleated hepatocytes. | Kaur, et al. [61] |
| Euclinostomum heterostomum | Apistogramma ramirezi syn. Mikrogeophagus ramirez (Myers & Harry, 1948) | Encysted skin | Imported from Hong Kong to Purdue University, USA | Irritating effect, rubbing against rocks & self-trauma. | Kazacos and Appel [50] |
| Euclinostomum heterostomum | Guppy, Poecilia reticulata Peters, 1859 | Musculature | Chonburi Province, Thailand | Localised degeneration & necrosis where parasite present. | Laoprasert, et al. [62] |
| Euclinostomum heterostomum | Striped snakehead, C. striata and Spotted snakehead, C. punctata | Liver | N/D | Rupture & loss of hepatocyte distinct shape. Vacuolation of cytoplasm. Hypertrophy of hepatocytes. Perilobular space of liver shows vacuolation, loosening of hepatic tissue & necrosis. | Laxma Reddy, et al. [63] |
| Euclinostomum heterostomum | Redbelly tilapia, T. zillii syn. Coptodon zillii | Encapsulated mostly in peritoneum of the kidney some musculature | Nile, Giza governorate, Egypt | Parasite cyst wall merged with renal interstitium & glomerular structures. Intense inflammatory cells extending into surrounding renal tissue. Significant glomerular & interstitial congestion, tubular epithelium with haemorrhages, vacuolization & necrosis. | Mahdy, et al. [64] |
| Euclinostomum heterostomum | African catfish, Clarias gariepinus (Burchell, 1822) | Muscle | Buffeldoorn Dam & Seshego Dam Lebowa, South Africa | Heavy infestations likely to occur in ponds with high fish density if intermediate snail host is present. Loss consumer confidence. | Mashego and Saayman [65] |
| Euclinostomum heterostomum | Nile tilapia, O. niloticus | Kidney | Commercial markets Cairo & fisherman Kafr El Sheikh, Egypt | Thick fibrous area surrounding parasite. Degenerative renal tissue, tubules & congested blood vessels. Some sections showed changes to glomeruli and Bowman’s capsule. | Mohamed, et al. [66] |
| Euclinostomum clarias (Dubois, 1930) Dollfus, 1932 | African catfish, C. gariepinus | Liver | Nigeria | Hepatic degeneration, necrosis/fibrosis, inflammation of bile duct, severe damage result of larval migration. | Onucha [67] |
| Euclinostomum heterostomum | Wild caught croaking gourami, Trichopsis vittata (Cuvier, 1831), Siamese fighting fish, Betta splendens Regan, 1910 and crescent betta, Betta imbellis Ladiges, 1975 | Musculature | Southern Thailand | Tubercle-like thickened areas on skin. | Pinky, et al. [68] |
| Euclinostomum heterostomum | Spotted snakehead, C. punctata | Liver, kidney, peritoneum, muscle, and ovary | Local fish market, Aligarh, North India | Tissue damage, infiltration immune cells cyst wall, chronic inflammation, granulomas. Liver degeneration hepatocytes, cytoplasmic vacuolation, nuclear alterations, mallory body formation, fibrosis, necrosis. Kidney distortion/dilation renal tubules, vacuolar degeneration, hypertrophy/hyperplasia tubular epithelial cells, occlusion tubules, fibrosis, haemorrhage, congestion glomeruli. | Shareef and Abidi [1] |
| Euclinostomum heterostomum | Guppy, P. reticulata cultured | Muscle | Kidchakan Supamattaya Aquatic Animal Health Research Center, Songkhla, southern Thailand | Abnormal swimming behaviour. Fish death severe infection. | Suanyuk, et al. [52] |
| Euclinostomum ardeolae | Nile tilapia, O. niloticus | Kidney | The Nile, Egypt | Cysts embedded kidney exerting pressure on tissue, black discolouration. | Tayel, et al. [69] |
| Isoparorchis hypselobagri (Billet, 1898) Ejsmont, 1932 (probably Isoparorchis trisimilitubis) | Wallago, Wallago attu (Bloch & Schneider, 1801) | Swim bladder | India | Infected fish unsuitable for human consumption. Patches of black pigments in the muscles and viscera of its hosts. Causes mortality and great economic loss. Adult parasites excrete poisonous metabolic substances within swim bladder. Ammonia is converted to urea. Urea high depending on parasite number. | Adak and Manna [70] |
| Isoparorchis hypselobagri (probably Isoparorchis trisimilitubis) | Wallago, W. attu | Swim bladder | India | Ammonia major excretory product. Amount of excreted ammonia differs depending on parasite number. Ammonotelic and ammonia can be formed by the action of several enzymes in Schistosoma mansoni as well. | Adak and Manna [71] |
| Isoparorchis hypselobagri (probably Isoparorchis trisimilitubis or Isoparorchis sp. 3) | Wallago, W. attu | Swim bladder | Dhaka, Bangladesh | Juvenile forms caused massive tissue damage, resulting erosions and tunnels in musculature, exudate, discoloration connective tissue, extreme melanisation, mixed inflammatory responses. | Alam [72] |
| Isoparorchis hypselobagri (probably Isoparorchis trisimilitubis) | Wallago, W. attu | Air bladder | Kakraiya lake, Jahangirabad, India | Inkspot disease. | Choudhary, et al. [73] |
| Isoparorchis hypselobagri (probably Isoparorchis trisimilitubis) | Long-whiskered catfish, Mystus aor syn. Sperata aor (Hamilton, 1822)Day’s mystus, Mystus bleekeri (Day, 1877) | Muscles, swim bladder, visceral organs, body cavity, viscera, some in the mouth, urinary system, biliary system, ovaries | Kuliarchar & Upazila rivers, India | Extensive tissue damage including inflammation, necrosis, and empty spaces with fragmented blood capillaries, tissue debris, lymphocytes and fluids. Infected liver, swim bladder and kidney showed vacuolation and massive melanisation. | Farhana and Khanum [74] |
| Isoparorchis hypselobagri (probably Isoparorchis trisimilitubis) | Spotted snakehead, C. punctata | Fins, liver, ovaries, abdominal cavity | Khookas bundh, Jaipur, India | Necrosis of fin tissues, scale loss. Necrotic areas with extensive inflammatory exudate formation were seen throughout the viscera. Liver reduced in size. Haemorrhage of intestinal wall. | Mahajan, et al. [18] |
| Isoparorchis hypselobagri (probably Isoparorchis trisimilitubis) | Red-crowned roofed turtle Kachuga kachuga syn. Batagur kachuga (Gray, 1831) | Body cavity | Hyderabad, India | N/D however this turtle is critically endangered in India and likely extinct Bangladesh. | Simha [75], Praschag, et al. [76] |
| Isoparorchis hypselobagri | Pungtungia herzi, Herzenstein, 1892 Acheilognathus koreensis syn. Tanakia koreensis (Kim & Kim, 1990), Squalidus japonicus coreanus syn. Squalidus japonicus (Sauvage, 1883) and Odontobutis platycephala Iwata & Jeon, 1985 | Muscle & lesions skin | Saengbiryang-myeon, Sancheong-gun, Gyeongsangnam-do, Korea | Inkspot disease, muscle and skin swellings and lesions. | Sohn and Na [77] |
| Isoparorchis hypselobagri (probably Isoparorchis trisimilitubis) | Mystus seenghala, syn. Sperata seenghala (Sykes, 1839) | Swim bladder | River Godavari, Rajahmundry, India | ‘‘Ink spot disease’’. | Vankara, et al. [78] |
| Genarchopsis goppo Ozaki, 1925 | Striped snakehead, C. striata | Intestine | Warangal, India | Histopathological changes include shortening and destruction of villi, vacuolation of sub mucous cells, dilation of blood vessels thickening of muscles and necrosis. In the infected fish carbohydrates, glycogen, protein and lipid contents are increased significantly to compensate for parasite presence. | Laxmareddy and Benarjee [17] |
| Genarchopsis paithanensis Pardeshi & Hiware, 2012 | Zig-zag eel, Mastacembelus armatus (Lacepède, 1800) | Intestine | India | Damage sub and mucosal layer and dilation blood vessels, destruction and extrusion of intestinal villi, inframammary and hyperplastic fibrosis. | Pardeshi and Hiware [79] |
| Genrachopsis goppo | Striped snakehead, C. striata | Intestine | Warangal, India | Severe damage to villi and other layers of intestine. Infections interfere with digestion and absorption of food material causing metabolic disturbances. Excretory products and metabolic end products excreted into intestine produce toxicity, interfere with protein metabolism of host. Host tissue may show decrease in protein content. | Reddy and Benarjee [16] |
| Genrachopsis goppo | Spotted snakehead, C. punctata | Intestine | Kakatiya, India | Glycogen content increased during infections to compensate for parasite needs. | Vinatha, et al. [80] |
| Pallisentis (P.) nagpurensis Bhalerao, 1931 | Gibelion catla, Catla catla syn. Labeo catla (Hamilton, 1822) and roho labeo, Labeo rohita (Hamilton, 1822) | N/D | Hyderabad, India | Overall protein in liver and intestine by 17%–26%. Amino acids increased by 14%–48.8% with highest increase in liver. | Kumar [81] |
| Pallisentis punctatin (misspelling likely Pallisentis (Brevitritospinus) punctati) | Spotted snakehead, C. punctata | Digestive tract | Hyderabad, India | Metabolic enzymes, succinate dehydrogenase and lactate dehydrogenase activity higher in infected fish. Oxidative stress enzymes lipid peroxidation, glutathione peroxidase and superoxide dismutase increased in infected fish. | Latha, et al. [82] |
| Pallisentis (P) nagapurensis | Striped snakehead, C striata | Intestine | Warangal district, Andhra Pradesh, India | Complete disruption intestinal mucosa and submucosa, thickened lamina propria, damage epithelial cells, mucosal folding & clumps. Villi shrunken, infected intestine enlarged and slightly inflamed. | Laxma Reddy and Benarjee [83] |
| Pallisentis (P) celatus (Van Cleave, 1928) Baylis, 1933 | Asian swamp eel, Monopterus albus (Zuiew, 1793) | Intestine | N/D | Mechanical damage to intestinal epidermis & muscle layer. | LI Chun-tao, et al. [84] |
| Acanthocephalan species not specified | Spotted snakehead, C. punctata | Intestine | River Gomti, Lucknow, India | Damage of intestinal tissues, shortening of villi, granuloma site of attachment. Erosion villi tip, necrosis & hyperplasia. | Verma and Saxena [85] |
| Senga sp. Dollfus, 1934 | Zig-zag eel, M. armatus & Snakehead, Channa sp. Scopoli, 1777 | Intestine | Maharashtra State, India | Damage intestinal villi, granuloma site of attachment. | Bhure and Nanware [86] |
| Senga mastacembelusae sp. nov. (not a valid species but Senga sp. likely) | Zig-zag eel, M. armatus | Intestine | Godavari Basin, India | Significant mechanical damage. Scolex deeply penetrating intestinal layers & damage mucosa, submucosa, muscularis mucosa. Intestinal villi architecture destruction & granuloma at scolex attachment. | Fartade and Fartade [87] |
| Senga rostellarae (probably Senga pahangensis or Senga filiformis) | Indonesian snakehead, Channa micropeltes (Cuvier, 1831) | Intestine | Kenyir Lake, Malaysia | Intestine with severe villus damage, destruction of villi epithelium and necrosis. Cross section of cestode showed increase of goblet cells and generated necrosis and severe damage to fish intestine. Conditions likely cause of death in fish due to haemorrhage and malabsorption of nutrients. | Hassan, et al. [88] |
| Senga species (probably Senga malayana for C. striata and Senga vishakapatnamensis for C. punctata) | Striped snakehead, C. striata & spotted snakehead, C. punctata | Intestine | Unknown | Excess mucus secretion, severe degeneration and necrosis in mucosal, submucosal, serosa layer and muscular layers at attachment. Ruptured serosa layer, vacuolization in tunica muscularis and lamina propria, shortened, fused and irregular shaped villous processes. | Kaur [89] |
| Senga sp. | Zig-zag eel, M. armatus | Intestine | India | Shortening, flattening and damage of villi and cyst formation in the intestine of fish. | Nanware and Bhure [90] |
| Senga sp. | Striped snakehead, C. striata | Intestine | Kaigaon Toka, India | Mechanical damage to intestinal tissue including shortening & damage to villi, thickening of the muscle layer, destruction of villi, hold fast penetration of the mucosa & damage to mucous & submucous membranes. | Shirsat, et al. [91] |
| Senga sp. | Siamese fighting fish, Betta splendens | Intestine | Aurangabad district, India | Destruction & extrusion of intestinal villi, fibroblast cell & plasma cell. | Wankhede, et al. [92] |
3.3.2. Genarchopsis sp. Pathophysiology
3.3.3. Isoparorchis sp., Southwell, 1913, Pathophysiology
3.3.4. Pallisentis (P.) and Pallisentis (B.) Pathophysiology
3.3.5. Senga Species Pathophysiology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shareef, P.; Abidi, S.M. Studies on the epidemiology and histopathology of Euclinostomum heterostomum (Trematoda; Digenea) infection in Channa punctata from North India. Arch. Pol. Fish 2015, 23, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Torrissen, O.; Jones, S.; Asche, F.; Guttormsen, A.; Skilbrei, O.T.; Nilsen, F.; Horsberg, T.E.; Jackson, D. Salmon lice–impact on wild salmonids and salmon aquaculture. J. Fish Dis. 2013, 36, 171–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares-Dias, M.; Martins, M.L. An overall estimation of losses caused by diseases in the Brazilian fish farms. J. Parasit. Dis. 2017, 41, 913–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, T. Parasites in cultured and feral fish. Vet. Parasitol. 1999, 84, 317–335. [Google Scholar] [CrossRef]
- Trujillo-González, A.; Becker, J.A.; Vaughan, D.B.; Hutson, K.S. Monogenean parasites infect ornamental fish imported to Australia. Parasitol. Res. 2018, 117, 995–1011. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.F.; Britton, J.R.; Turnbull, J.F. A risk assessment for managing non-native parasites. Biol. Invasions 2013, 15, 1273–1286. [Google Scholar] [CrossRef]
- Peeler, E.J.; Oidtmann, B.C.; Midtlyng, P.J.; Miossec, L.; Gozlan, R.E. Non-native aquatic animals introductions have driven disease emergence in Europe. Biol. Invasions 2011, 13, 1291–1303. [Google Scholar] [CrossRef] [Green Version]
- Dove, A.; Cribb, T.; Mockler, S.; Lintermans, M. The Asian fish tapeworm, Bothriocephalus acheilognathi, in Australian freshwater fishes. Mar. Freshw. Res. 1997, 48, 181–183. [Google Scholar] [CrossRef]
- Dove, A.D.M.; Ernst, I. Concurrent invaders—Four exotic species of Monogenea now established on exotic freshwater fishes in Australia. Int. J. Parasitol. 1998, 28, 1755–1764. [Google Scholar] [CrossRef]
- Hassan, M.; Beatty, S.; Morgan, D.; Doupé, R.; Lymbery, A. An introduced parasite, Lernaea cyprinacea L., found on native freshwater fishes in the south west of Western Australia. J. R. Soc. West. Aust. 2008, 91, 149–153. [Google Scholar]
- Zhu, X.; Barton, D.P.; Wassens, S.; Shamsi, S. Morphological and genetic characterisation of the introduced copepod Lernaea cyprinacea Linnaeus (Cyclopoida: Lernaeidae) occurring in the Murrumbidgee catchment, Australia. Mar. Freshw. Res. 2020, 72, 876–886. [Google Scholar]
- Whittington, R.J.; Chong, R. Global trade in ornamental fish from an Australian perspective: The case for revised import risk analysis and management strategies. Prev. Vet. Med. 2007, 81, 92–116. [Google Scholar] [PubMed]
- Quartey, S.H.; Wells, S. The fishing industry’s perception of its contribution towards the sustainability of the Eyre Peninsula region, Australia. Australas. J. Reg. Stud. 2019, 25, 135–176. [Google Scholar]
- BICON. Case: Finfish (Excluding Salmonid) for Human Consumption Effective: 07 Apr 2020. Available online: https://bicon.agriculture.gov.au/BiconWeb4.0/ImportConditions/Questions/EvaluateCase?elementID=0000067916&elementVersionID=360 (accessed on 15 April 2020).
- FRDC. Seafood Import and Export by Volume. In Seafood Trade Data; The Fisheries Research and Development Corporation: Canberra, Australia, 2020. [Google Scholar]
- Reddy, B.L.; Benarjee, G. Histochemical studies on Genarchopsis goppo from fresh water murrel, Channa striatus. Asian J. Anim. Sci. 2011, 6, 23–26. [Google Scholar]
- Laxmareddy, B.; Benarjee, G. Intestinal histopathology of trematode infected fish, Channa striatus. BioLIfe 2013, 1, 29–31. [Google Scholar]
- Mahajan, C.; Agrawal, N.; John, M.; Katta, V. Effect of the digenean Isoparorchis hypselobagri (Billet, 1898) on an airbreathing fish Channa punctatus (Bloch) with particular reference to biochemical and haematological changes. J. Fish Dis. 1979, 2, 519–528. [Google Scholar] [CrossRef]
- Pinto, R.; Barros, L.; Tortelly, L.; Teixeira, R.; Gomes, D. Prevalence and pathology of helminths of ciconiiform birds from the Brazilian swamplands. J. Helminthol. 2004, 78, 259. [Google Scholar] [CrossRef]
- Mihalca, A.; Fictum, P.; Škorič, M.; Sloboda, M.; Kärvemo, S.; Ghira, I.; Carlsson, M.; Modrý, D. Severe granulomatous lesions in several organs from Eustrongylides larvae in a free-ranging dice snake, Natrix tessellata. Vet. Pathol. 2007, 44, 103–105. [Google Scholar]
- Echi, P.; Eyo, J.; Okafor, F.; Onyishi, G.; Ivoke, N. First record of Co–infection of three clinostomatid parasites in cichlids (Osteichthyes: Cichlidae) in a tropical freshwater lake. Iran. J. Public Health 2012, 41, 86–90. [Google Scholar]
- Fan, P. Viability of metacercariae of Clonorchis sinensis in frozen or salted freshwater fish. Int. J. Parasitol. 1998, 28, 603–605. [Google Scholar] [CrossRef]
- Grabda, J.; Bier, J.W. Cultivation as an estimate for infectivity of larval Anisakis simplex from processed herring. J. Food Prot. 1988, 51, 734–736. [Google Scholar] [CrossRef]
- Sirisinha, S.; Chawengkirttikul, R.; Sermswan, R. Immunodiagnosis of opisthorchiasis. Southeast Asian J. Trop. Med. Public Health 1991, 22, 179–183. [Google Scholar] [PubMed]
- Mas, C.S.; Bargues, M.D. Human liver flukes: A review. Res. Rev. Parasitol. 1997, 57, 145–218. [Google Scholar]
- Yossepowitch, O.; Gotesman, T.; Assous, M.; Marva, E.; Zimlichman, R.; Dan, M. Opisthorchiasis from imported raw fish. Emerg. Infect. Dis. 2004, 10, 2122–2126. [Google Scholar] [CrossRef] [PubMed]
- Knibb, W.; Le, C.; Katouli, M.; Bar, I.; Lloyd, C. Assessment of the origin of white spot syndrome virus DNA sequences in farmed Penaeus monodon in Australia. Aquaculture 2018, 494, 26–29. [Google Scholar] [CrossRef]
- Loynes, K. White Spot Disease in Australia up to 2017: A Chronology; Science Technology and Natural Resources: Canberra, Australia, 2017. [Google Scholar]
- FRDC. Summary Overview: Economic Impact of 2016 White Spot Disease Outbreak; Fisheries Research and Development Corporation: Deacon West, Australia, 2016. [Google Scholar]
- Williams, M.; Hernandez-Jover, M.; Williams, T.; Shamsi, S. A risk scoring system for seafood supply chain breaches and examination of freshwater fish imported to Australia. FQS Food Qual. Saf. 2021, 5, fyab004. [Google Scholar] [CrossRef]
- Williams, M.; Hernandez-Jover, M.; Shamsi, S. Parasites of zoonotic interest identified in selected imported freshwater fish. Food Waterborne Parasitol. 2021, 26, e00138. [Google Scholar] [CrossRef] [PubMed]
- Conte-Grand, C.; Britz, R.; Dahanukar, N.; Raghavan, R.; Pethiyagoda, R.; Tan, H.H.; Hadiaty, R.K.; Yaakob, N.S.; Rüber, L. Barcoding snakeheads (Teleostei, Channidae) revisited: Discovering greater species diversity and resolving perpetuated taxonomic confusions. PLoS ONE 2017, 12, e0184017. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.; Moore, J.W.; Herborg, L.-M.; Murray, C.C.; Serrao, N.R. A non-native snakehead fish in British Columbia, Canada: Capture, genetics, isotopes, and policy consequences. Manag. Biol. Invasions 2013, 4, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.J.; Andriyono, S.; Sektiana, S.P.; Rahman, M.M.; Kim, H.-W. The molecular characterization of complete mitochondrial genome of spotted snakehead fish, Channa punctata (Bloch 1793). Mitochondrial DNA Part B 2019, 4, 547–548. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.Z.; Hossain, M.M.M. Isolation and characterization of internal parasites in snakehead. Int. J. Fish. Aquat. Stud. 2015, 2, 17–22. [Google Scholar] [CrossRef]
- Shamsi, S.; Suthar, J. A revised method of examining fish for infection with zoonotic nematode larvae. Int. J. Food Microbiol. 2016, 227, 13–16. [Google Scholar]
- Bier, J.W.; Jackson, G.J.; Adams, A.M.; Rude, R.A. Bacteriological Analytical Manual, Chapter 19, Parasitic Animals in Foods. Available online: https://www.fda.gov/Food/FoodScienceResearch/LaboratoryMethods/ucm071468.htm (accessed on 20 February 2018).
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, T.; Cribb, T.H.; Miller, T.L.; Urabe, M.; Van Ha, N.; Binh, T.T.; Shed’ko, M.B. Revision of Isoparorchis Southwell, 1913 (Digenea, Hemiuroidea, Isoparorchiidae), parasites of the air bladder of freshwater catfishes: A molecular and morphological study. Bull. Natl. Mus. Nat. Sci. Ser. A 2014, 40, 15–51. [Google Scholar]
- Urabe, M.; Nishimura, T.; Shimazu, T. Taxonomic revision of three species of the genus Genarchopsis (Digenea: Hemiuroidea: Derogenidae) in Japan by molecular phylogenetic analyses. Parasitol. Int. 2012, 61, 554–560. [Google Scholar] [PubMed]
- Pardeshi, P.; Hiware, C. Comparative Study on Species of Genarchosis Parasitizing Mastacembelus armatus Lecepede (1800). BIOINFOLET Q. J. Life Sci. 2012, 9, 79–83. [Google Scholar]
- Bhadauria, S.; Dandotia, M. Studies on the trematode parasites of fresh water fishes with special reference to Gwalior region. II: On one new genus and some unknown and known species. Riv. Parassitol. 1984, 45, 341–383. [Google Scholar]
- Koiri, R.; Roy, B. The occurrence of a new piscine tapeworm Senga Dollfus, 1934 (Pseudophyllidea: Bothriocephalidae) in Monopterus cuchia (F. Hamilton, 1822) from Tripura. Imp. J. Interdiscip. Res. 2017, 3, 864–870. [Google Scholar]
- Majid, M.; Shinde, G. Two new species of the genus Senga Dollfus, 1934 (Cestoda: Pseudophyllidea) from freshwater fish at Jagannathpuri, Orissa. Indian J. Parasitol. 1984, 8, 169–172. [Google Scholar]
- Pardeshi, P.; Hiware, C. A new Pseudophyllidean Senga rupchandensis n. sp. from Channa striatus (Bloch, 1793) at Jalna District (MS), India. Recent Res. Sci. Technol. 2011, 3, 17–22. [Google Scholar]
- Amin, O.M.; Heckmann, R.A.; Nguyen, V.H.; Pham, V.L.; Pham, N.D. Revision of the genus Pallisentis (Acanthocephala: Quadrigyridae) with the erection of three new subgenera, the description of Pallisentis (Brevitritospinus) vietnamensis subgen. et sp. n., a key to species of Pallisentis, and the description of a new quadrigyrid genus, Pararaosentis gen. n. Comp. Parasitol. 2000, 67, 40–50. [Google Scholar]
- Gupta, S.P.; Verma, S. On three new Acanthocephala parasites of the genus Pallisentis Van Cleave, 1928 from fresh water fishes of Lucknow. Helminthologia 1980, 17, 269–282. [Google Scholar]
- Gautam, N.K.; Misra, P.K.; Saxena, A.M. Four new species of the genus Pallisentis (Quadrigyridae, Van Cleave, 1920) from freshwater fish in Uttar Pradesh, India. Acta Parasitol. 2019, 64, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Shareef, P.A.; Abidi, S.M.A. Cysteine protease is a major component in the excretory/secretory products of Euclinostomum heterostomum (Digenea: Clinostomidae). Parasitol. Res. 2014, 113, 65–71. [Google Scholar] [CrossRef]
- Kazacos, K.R.; Appel, G.O. Euclinostomum heterostomum metacercariae. PHSWA 1983, 50, 103–107. [Google Scholar]
- Britz, J.; Van As, J.; Saayman, J. Occurrence and distribution of Clinostomum tilapiae Ukoli, 1966 and Euclinostomum heterostomum (Rudolphi, 1809) metacercarial infections of freshwater fish in Venda and Lebowa, Southern Africa. J. Fish Biol. 1985, 26, 21–28. [Google Scholar] [CrossRef]
- Suanyuk, N.; Mankhakhet, S.; Soliman, H.; Saleh, M.; El-Matbouli, M. Euclinostomum heterostomum infection in guppies Poecilia reticulata cultured in southern Thailand. Dis. Aquat. Org. 2013, 104, 121–127. [Google Scholar] [CrossRef]
- Ikisa, K.; Babatunde, B.; Hart, A. Histopathological variations in gills, liver and kidney of nile tilapia-Oreochromis niloticus exposed to benzalkoniumbhloride mixture with treated produced water. J. Appl. Sci. Environ. Manag. 2019, 23, 1181–1187. [Google Scholar] [CrossRef]
- Akiyoshi, H.; Inoue, A. Comparative histological study of teleost livers in relation to phylogeny. Zool. Sci. 2004, 21, 841–850. [Google Scholar] [CrossRef] [Green Version]
- Witeska, M. Anemia in teleost fishes. Bull. Eur. Assoc. Fish Pathol. 2015, 35, 149–160. [Google Scholar]
- Roychowdhury, P.; Aftabuddin, M.; Pati, M.K. Thermal stress altered growth performance and metabolism and induced anaemia and liver disorder in Labeo rohita. Aquac. Res. 2020, 51, 1406–1414. [Google Scholar] [CrossRef]
- Ahmed, M.; El-Ganainy, S.; El-Aziz, A.; Shaban, H. Morphological and Electrophoretic Differentiation of Two Clinostomatid Metacercariae Infecting Oreochromis niloticus from the River Nile at El-Minia District, Egypt. Egypt. Acad. J. Biol. Sci. E Med. Entomol. Parasitol. 2018, 10, 105–115. [Google Scholar] [CrossRef]
- Bhargavi, G.; Prasanna, D.; Rani, R.J. Invitro studies on antihelminthic activity of seeds of Butea frondosa on the metacercaria of Euclenostomum heterostomum a parasite of fresh water fish Channa punctatus. In Proceedings of the 22nd National Congress on Parasitology, Kalyani, India, 30 October–1 November 2010. [Google Scholar]
- Echi, P.; Iyaji, F.; Ejere, V.; Abuh, S. Dynamics of synchronized clinostomatids infections in cichlids. Environ. Conserv. J. 2014, 15, 49–54. [Google Scholar]
- Kaur, P.; Shrivastav, R.; Qureshi, T. Pathological effect on naturally infected kidneys of freshwater murrels by Euclinostomum heterostomum metacercariae. J. Parasit. Dis. 2016, 40, 157–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, P.; Qureshi, T.; Shrivastav, R. Histo-architectural alterations induced by Euclinostomum heterostomum (metacercariae) infected liver of Channa punctata (Bloch). J. Parasit. Dis. 2012, 36, 197–199. [Google Scholar] [CrossRef] [Green Version]
- Laoprasert, T.; Naksuwon, S.; Maneepitaksanti, W.; Chinabut, S. Parasitic digenean Euclinostomum sp.(Trematoda: Clinostomatidae) in guppy (Poecilia reticulata). Thai Fisheries Gazette 2006, 59, 323–331. [Google Scholar]
- Reddy, B.L.; Benarjee, G.; Pradesh, A. Comparative liver histopathology of fresh water murrels due to trematode, Euclinostomum heterostomum. BioLife 2014, 2, 369–399. [Google Scholar]
- Mahdy, O.A.; Abdel-Maogood, S.Z.; Mohammed, F.F.; Salem, M.A. Effect of Verbesina Alternifolia and Mentha Piperita Oil Extracts on Newly Excysted Metacercaria of Euclinostomum heterostomum (Rudolphi, 1809) (Digenea: Clinostomatidae) from Naturally Infected Kidneys of Tilapia Zillii in Egypt. J. Egypt. Soc. Parasitol. 2017, 47, 513–521. [Google Scholar]
- Mashego, S.N.; Saayman, J.E. Digenetic trematodes and cestodes of Clarias gariepinus (Burchell. 1822) in Lebowa, South Africa with taxonomic notes. S. Afr. J. Wildl. Res. 1989, 19, 17–20. [Google Scholar]
- Mohamed, I.R.; Wafaa, I.I.; Zakaria, S.M.; Rabab, M.A.; Fekry, M. Histological and Histochemical Changes in Liver and Kidney of Some Fishes Induced by Natural Larval Trematodes Infection. Researchgate 2008, 1–19. Available online: https://www.researchgate.net/publication/335967409_Histological_and_Histochemical_Changes_in_Liver_and_Kidney_of_Some_Fishes_Induced_by_Natural_Larval_Trematodes_Infection (accessed on 19 February 2023).
- Onucha, E. Liver pathology of the african catfish, Clarias gariepinus infected with Euclinostomum clarias metacercariae in Anambra river basin, Nigeria. Anim. Res. Int. 2010, 7, 1151–1155. [Google Scholar]
- Pinky, K.; Rekha, S.; Dar, B.A.; Jha, G.N.; Qureshi, T.A. Suppression of ovarian development in freshwater fishes due to endo-helminthic infection. Indian J. Fish. 2015, 62, 128–132. [Google Scholar]
- Tayel, I.S.; Ahmed, M.N.; Ramadan, A.E. Histopathological alterations and parasitic infection in Oreochromis niloticus fish inhabiting the River Nile water. Egypt. J. Aquat. Biol. Fish. 2020, 24, 233–247. [Google Scholar] [CrossRef]
- Adak, S.; Manna, B. Urea Excretion in Isoparorchis hypselobagri (Billet, 1898). Proc. Zool. Soc. 2013, 66, 100–104. [Google Scholar] [CrossRef]
- Adak, S.; Manna, B. Study on the rate of ammonia excretion in Isoparorchis hypselobagri (Billet, 1898). Zool. Res. Hum. Welf. 2008, 27, 279–283. [Google Scholar]
- Alam, S. Parasite Diversity and Their Effects on Histological and Biochemical Components of Wallago attu (Bloch-Schneider, 1801) and Rita rita (Hamilton-Buchanan, 1822). Ph.D. Thesis, University of Dhaka, Dhaka, Bangladesh, 2019; p. 285. [Google Scholar]
- Choudhary, K.; Ray, S.; Agrawal, N. Record of Some Digeneans, Infecting Freshwater Fishes, Snails, and Amphibians in India, using 28S Ribosomal DNA Marker. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2019, 90, 501–513. [Google Scholar] [CrossRef]
- Farhana, R.; Khanum, H. The effect of parasitic infestation on tissues and organs of Mystus aor (Hamilton) and Mystus bleekeri (Day). In Advances in Parasitology: A Novel Approach towards a Disease-Free World, Proceedings of the 22nd National Congress of Parasitology, West Bengal, India, 30 October–1 November 2011; Department of Zoology, University of Kalyani: Kalyani, West Bengal, India, 2011; pp. 223–227. [Google Scholar]
- Simha, S.S. Studies on the trematode parasites of reptiles found in Hyderabad State. Z. Parasitenkd. 1958, 18, 161–218. [Google Scholar] [CrossRef]
- Praschag, P.; Ahmed, M.F.; Das, I.; Singh, S. Batagur kachuga (Errata Version Published in 2019). Available online: https://www.iucnredlist.org/ (accessed on 15 November 2020).
- Sohn, W.-M.; Na, B.-K. Morphological Characteristics and Fish Hosts of Isoparorchis sp. (Digenea: Isoparorchiidae) in Korea. Korean J. Parasitol. 2018, 56, 501. [Google Scholar] [CrossRef] [Green Version]
- Vankara, A.P.; Mani, G.; Vijayalakshmi, C. A report on various digenetic metacercariae from the freshwater fishes of River Godavari, Rajahmundry. J. Parasit. Dis. 2011, 35, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Pardeshi, P.; Hiware, C. Histopathological studies on the intestine of the Mastacembelus armatus parasitized bytrematode, Genarchopsis paithanensis n. sp. BIOINFOLET Q. J. Life Sci. 2014, 11, 1209–1211. [Google Scholar]
- Vinatha, N.; Bikshapathi, V.; Babu, S.R. Post Helminthes infection changes in AAT and AIAT activity in the myotomal muscles of Channa punctatus. Sci. Res. Report. 2013, 3, 167–170. [Google Scholar]
- Kumar, P.A. Biochemical effects on protein and free amino acid metabolism in Catla catla and Labeo rohita due to Pallisentis nagpurensis infection. AIJRFANS Am. Int. J. Res. Form. Appl. Nat. Sci. 2014, 6, 82–85. [Google Scholar]
- Latha, J.N.; Ramesh, K.; Vanithadas, V. Studies on Enzymatic Activities by Gastro-Intestinal Helminthic Parasites in Channa Punctata (Snake Headed or Bloch). IJIAR Int. J. Res. Anal. Rev. 2019, 6, 807–812. [Google Scholar]
- Reddy, B.L.; Benarjee, G. Parasitic adaptation of fresh water fish, Channa striatus. Int. J. Fish. Aquat. Stud. 2015, 2, 69–71. [Google Scholar]
- Li, C.-T.; Li, X.; Zhao, Y.-Q.; Zeng, B.-P. Morphological characteristics of Pallisentis celatus and pathological changes of the intestine in Monopterus allbus infected by Pallisentis celatus. J. South. Agric. 2014, 45, 1881–1885. [Google Scholar]
- Verma, S.K.; Saxena, A. Histopathological study of freshwater fish Channa punctatus (bloch, 1793) infected with acanthocephalan parasites from river Gomti, Lucknow, Uttar Pradesh (India). Trends Fish. Res. 2018, 7, 57–64. [Google Scholar]
- Bhure, D.B.; Nanware, S.S. Hold-Fast Organs of Piscean and Avian Cestode Parasites with Special Emphasis on Histopathology. WSN World Sci. News 2016, 34, 109–120. [Google Scholar]
- Fartade, A.M.; Fartade, M.M. Histopathological study of fresh water fishes infected with ptychobothridean tapeworms parasite from Godavri basin MS (India). Int. J. Res. BioSci. 2016, 5, 39–42. [Google Scholar]
- Hassan, M.; Hadi, M.S.I.A.; Aziz, M.F.H.A.; Wahab, W.; Abdullah, F.; Deraman, S.; Zakariah, M.I. Histopathological changes in the intestine of Channa micropeltes infected with the cestode Senga rostellarae. IOP Conf. Ser. Earth Environ. Sci. 2019, 370, 012055. [Google Scholar] [CrossRef]
- Kaur, P. Histo-pathological effect of Senga species (Cestode: Pseudophyllidea) in intestine of piscian hosts. WJPPS World J. Pharm. Pharm. Sci. 2014, 3, 1506–1513. [Google Scholar]
- Nanware, S.S.; Bhure, D.B. Intestinal histopathology of Mastacembelus armatus parasitized by pseudophyllidean cestodes. Flora Fauna 2016, 22, 125–130. [Google Scholar]
- Shirsat, J.; More, V.; Wankhede, S. Histopathological changes caused by Senga sp. In Channa straitus. IJCRLS Int. J. Curr. Res. Life Sci. 2018, 7, 2659–2660. [Google Scholar]
- Wankhede, H.; Aade, U.; Kaldate, K. Histopathological study of ptychobothriidan tapeworm in freshwater fish Mastacembelus armatus (Lecepede, 1800) from Aurangabad district (ms). ISRJ Indian Streams Res. J. 2012, 2, 1–4. [Google Scholar]
- Urabe, M. Life cycle of Genarchopsis goppo (Trematoda: Derogenidae) from Nara, Japan. J. Parasitol. 2001, 87, 1404–1408. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose metabolism in fish: A review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef]
- Arts, M.T.; Kohler, C.C. Health and condition in fish: The influence of lipids on membrane competency and immune response. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 237–256. [Google Scholar]
- Randall, D.J.; Tsui, T.K.N. Ammonia toxicity in fish. Mar. Pollut. Bull. 2002, 45, 17–23. [Google Scholar] [CrossRef]
- Randall, D.J.; Wright, P.A. Ammonia distribution and excretion in fish. Fish Physiol. Biochem. 1987, 3, 107–120. [Google Scholar] [CrossRef]
- Taraschewski, H. Acanthocephala. In Encyclopedia of Parasitology, 3rd ed.; Taraschewski, H., Mehlhorn, H., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2008; pp. 3–39. [Google Scholar]
- Taraschewski, H. Host-parasite interactions in Acanthocephala: A morphological approach. Adv. Parasitol. 2000, 46, 1–179. [Google Scholar]
- Szalai, A.J.; Danell, G.; Dick, T.A. Intestinal leakage and precipitating antibodies in the serum of quillback, Carpiodes cyprinus (Lesueur), infected with Neoechinorhynchus carpiodi Dechtiar, 1968 (Acanthocephala: Neoechinorhynchidae). J. Parasitol. 1988, 74, 415–420. [Google Scholar] [CrossRef]
- Taraschewski, H. Host-parasite interface of fish acanthocephalans. I. Acanthocephalus anguillae (Palaeacanthocephala) in naturally infected fishes: LM and TEM investigations. Dis. Aquat. Org. 1988, 4, 109–119. [Google Scholar] [CrossRef]
- Bristol, J.R.; Mayberry, L.F.; Huber, D.; Ehrlich, I. Endoparasite fauna of trout in the Plitvice Lakes National Park. Vet. Arh. 1984, 54, 5–11. [Google Scholar]
- Buchmann, K. On the infection of Baltic cod (Gadus morhua L.) by the acanthocephalan Echinorhynchus gadi (Zoega) Müller. Nord. Vet. 1986, 38, 308–314. [Google Scholar]
- Kuchta, R.S.T. Bothriocephalidea In Planetary Biodiversity Inventory (2008–2017): Tapeworms from the Bowels of the Earth; Caira, J.N., Jensen, K., Eds.; Natural History Museum, The University of Kansas: Lawrence, KS, USA, 2017; Volume 25, pp. 29–47. [Google Scholar]
- Brabec, J.; Waeschenbach, A.; Scholz, T.; Littlewood, D.T.J.; Kuchta, R. Molecular phylogeny of the Bothriocephalidea (Cestoda): Molecular data challenge morphological classification. Int. J. Parasitol. 2015, 45, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Shaharom, F. Fish Parasites of Lake Kenyir, Peninsular Malaysia; Penerbit UMT, Universiti Malaysia: Kuala Terengganu, Malaysia, 2012; Volume 1, pp. 1–173. [Google Scholar]
- Dawood, M.A. Nutritional immunity of fish intestines: Important insights for sustainable aquaculture. Rev. Aquac. 2020, 13, 642–663. [Google Scholar] [CrossRef]
- Begam, M.; Sengupta, M. Immunomodulation of intestinal macrophages by mercury involves oxidative damage and rise of pro-inflammatory cytokine release in the fresh water fish Channa punctatus Bloch. Fish Shellfish Immunol. 2015, 45, 378–385. [Google Scholar] [CrossRef]
- Okamura, B.; Feist, S.W. Emerging diseases in freshwater systems. Freshw. Biol. 2011, 56, 627–637. [Google Scholar] [CrossRef]
- Jones, S.R. The occurrence and mechanisms of innate immunity against parasites in fish. Dev. Comp. Immunol. 2001, 25, 841–852. [Google Scholar] [CrossRef]
- Britton, J.R.; Pegg, J.; Williams, C.F. Pathological and ecological host consequences of infection by an introduced fish parasite. PLoS ONE 2011, 6, e26365. [Google Scholar] [CrossRef]
- Rowley, J.J.L.; Gleason, F.H.; Andreou, D.; Marshall, W.L.; Lilje, O.; Gozlan, R. Impacts of mesomycetozoean parasites on amphibian and freshwater fish populations. Fungal Biol. Rev. 2013, 27, 100–111. [Google Scholar] [CrossRef]
- Kuchta, R.; Radačovská, A.; Bazsalovicsová, E.; Viozzi, G.; Semenas, L.; Arbetman, M.; Scholz, T. Host switching of zoonotic broad fish tapeworm (Dibothriocephalus latus) to salmonids, Patagonia. Emerg. Infect. Dis. 2019, 25, 2156. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, C.; Fitch, D. Colonization, larval survival and epidemiology of the nematode Anguillicola crassus, parasitic in the eel, Anguilla anguilla, in Britain. J. Fish Biol. 1990, 36, 117–131. [Google Scholar] [CrossRef]
- NESP. 22 Australian Freshwater Fish at Risk of Extinction. Available online: https://www.nespthreatenedspecies.edu.au/news-and-media/media-releases/22-australian-freshwater-fish-at-risk-of-extinction (accessed on 11 March 2023).
- Lintermans, M.; Geyle, H.M.; Beatty, S.; Brown, C.; Ebner, B.C.; Freeman, R.; Hammer, M.P.; Humphreys, W.F.; Kennard, M.J.; Kern, P. Big trouble for little fish: Identifying Australian freshwater fishes in imminent risk of extinction. Pac. Conserv. Biol. 2020, 26, 365–377. [Google Scholar] [CrossRef]
- Abd-Al-Aal, Z.; Amer, O.; Badawy, A.; El-Ashram, A. Digenetic trematodes of the little egret, Egretta garzetta, and possibility of transmission to Oreochromis niloticus at El-abbassa fish farms, Egypt. In Proceedings of the 8th International Symposium on Tilapia in Aquaculture, Cairo, Egypt, 12–14 October 2008; pp. 1351–1364. [Google Scholar]
- DPI NSW. Tilapia. Available online: https://www.dpi.nsw.gov.au/fishing/aquatic-pests-diseases/freshwater-pests/species/tilapia (accessed on 8 June 2020).
- Webb, A.C. The Ecology of Invasions of Non-Indigenous Freshwater Fishes in Northern Queensland. Ph.D. Thesis, James Cook University, Brisbane, Australia, 2003. [Google Scholar]
- Caffara, M.; Locke, S.A.; Cristanini, C.; Davidovich, N.; Markovich, M.P.; Fioravanti, M.L. A combined morphometric and molecular approach to identifying metacercariae of Euclinostomum heterostomum (Digenea: Clinostomidae). J. Parasitol. 2016, 102, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Echi, P.C.; Eyo, J.E.; Okafor, F.C. Co-parasitism and morphometrics of three Clinostomatids (Digenea: Clinostomatidae) in Sarotherodon melanotheron from a tropical freshwater lake. Anim. Res. Int. 2009, 6, 982–986. [Google Scholar] [CrossRef] [Green Version]
- Falk, R.; Smirnov, M.; Ofek, T.; Cummings, D. Parasitological Survey of Cichlidae in the Sea-of-Galilee. Isr. J. Aquac.-Bamidgeh 2020, 72, 1–6. [Google Scholar] [CrossRef]
- Gomon, M.F. Cichlidae. Available online: https://fishesofaustralia.net.au/home/family/273#moreinfo (accessed on 10 March 2023).
- Senapin, S.; Phiwsaiya, K.; Laosinchai, P.; Kowasupat, C.; Ruenwongsa, P.; Panijpan, B. Phylogenetic analysis of parasitic trematodes of the genus Euclinostomum found in Trichopsis and Betta fish. J. Parasitol. 2014, 100, 368–371. [Google Scholar] [CrossRef] [PubMed]
- DPI. Identifying Liver Fluke Snails; Department of Primary Industry: Orange, Australia, 2017; pp. 1–6. [Google Scholar]
- Kingsford, R.T.; Porter, J.L.; Brandis, K.J.; Ryall, S. Aerial surveys of waterbirds in Australia. Sci. Data 2020, 7, 172. [Google Scholar] [CrossRef]
- Abro, M.M.; Dharejo, A.M.; Khan, M.M.; Birmani, N.A. Euclinostomum heterostomum (Rud., 1809) Travassos, 1928 (Trematodes: Clinostomidae: Euclinostominae): A New Record in Avian Host Little Cormorant (Aves: Phalacrocoracidae) of Pakistan. J. Adv. Biol. 2016, 8, 1–5. [Google Scholar] [CrossRef]
- Dennis, E.A.; Sharp, M. Morphology of Euclinostomum heterostomum (Rudolphi, 1809) Trematoda: Clinostomatidae) from Bulbicus ibis. J. Helminthol. 1973, 47, 17–25. [Google Scholar] [CrossRef]
- Jhansilakshmibai, K.; Madhavi, R. Euclinostomum heterostomum (Rudolphi, 1809) (Trematoda): Life-cycle, growth and development of the metacercaria and adult. Syst. Parasitol. 1997, 38, 51–64. [Google Scholar] [CrossRef]
- Mansour, R.M. First record of Euclinostomum heterostomum from the naturally-infected heron “Ardeola ralloides” in Egypt: A light & scanning electron microscopy study. EJZ Egypt. J. Zool. 2019, 72, 22–31. [Google Scholar]
- Gomon, M.F. Glossogobius giuris Fishes of Australia. Available online: https://fishesofaustralia.net.au/home/species/129 (accessed on 27 November 2020).
- Bray, D.J. Glossogobius munroi. Available online: https://fishesofaustralia.net.au/home/species/5187 (accessed on 27 October 2020).
- Hoese, D.; Allen, G.R.; Hadiaty, R.K. Description of three new species of dwarf Glossogobius from New Guinea and northern Australia. Cybium 2017, 41, 179–193. [Google Scholar]
- Hoese, D.F.; Allen, G.R. A review of the amphidromous species of the Glossogobius celebius complex, with description of three new species. Cybium 2011, 35, 269–285. [Google Scholar]
- Hoese, D.F. The introduction of the gobiid fishes Acanthogobius flavimanus and Tridentiger trigonocephalus into Australia. Koolewong 1973, 2, 3–5. [Google Scholar]
- Abol-Munafi, A.B.; Ambak, M.A.; Ismail, P.; Tam, B. Molecular data from the cytochrome b for the phylogeny of Channidae (Channa sp.) in Malaysia. Biotechnology 2007, 6, 22–27. [Google Scholar]
- Fishtraderlau. Snakehead. Available online: http://www.aceforums.com.au/topic/46516-snakehead/ (accessed on 29 November 2020).
- Bagni, L.; Galli, C. Gabbia. Available online: https://www.molluscabase.org/aphia.php?p=taxdetails&id=818268 (accessed on 1 July 2021).
- ALA. Preserved Specimen of Stenocypris malcolmsonii. Available online: https://biocache.ala.org.au/occurrences/82178328-cbca-4117-8cfd-f9c43657f2fe (accessed on 22 February 2021).
- De Deckker, P. Ostracods of athalassic salt lakes. Dept. of Biogeography & Geomorphology, Australian National University, Canberra ACT. In Salt Lakes, Proceedings of the International Symposium on Athalassic (Inland) Salt Lakes, Adelaide, Australia, October 1979; Springer: Dordrecht, The Netherlands, 1981; p. 131. [Google Scholar]
- Martens, K.; Savatenalinton, S. A subjective checklist of the Recent, free-living, non-marine Ostracoda (Crustacea). Zootaxa 2011, 2855, 1–79. [Google Scholar] [CrossRef] [Green Version]
- Christy, M.T.; Kirkpatrick, W. Striped Snakehead (Channa striata): An Invasive Animals CRC Project; Department of Primary Industries and Regional Development: Perth, Australia, 2017; pp. 1–24. [Google Scholar]
- Bashirullah, A.K. On the occurrence of the trematode, Isoparorchis hypselobagri (Billet 1898), in fishes and notes on its life history. Nor. J. Zool. 1972, 20, 2009–2012. [Google Scholar]
- Arthur, J.R.; Ahmed, A.T.A. Checklist of the Parasites of Fishes of Bangladesh; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; pp. 1–88. [Google Scholar]
- Sahadev, K.V.; Simha, S.S. A new host record for Isoparorchis hypselobagri. Geobios 1980, 7, 272–273. [Google Scholar]
- Rao, L.N.; Kameswari, M.; Rao, G.R.H. A yet undefined host of Isoparorchis hypselobagri Billet, 1898. Curr. Sci. 1979, 48, 320. [Google Scholar]
- Van Dyke, J.U.; Ferronato, B.D.O.; Spencer, R.-J. Current conservation status of Australian freshwater turtles. Aust. J. Zool. 2018, 66, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Hero, J.-M.; Morrison, C.; Gillespie, G.; Roberts, J.D.; Newell, D.; Meyer, E.; McDonald, K.; Lemckert, F.; Mahony, M.; Osborne, W. Overview of the conservation status of Australian frogs. Pac. Conserv. Biol. 2006, 12, 313–320. [Google Scholar] [CrossRef]
- Chandler, A.C. The Prevalence and Epidemiology of Hookworm and other Helminthic Infections in India. Part IV. Assam and the Hill Areas of Eastern Bengal. IJMR Indian J. Med. Res. 1926, 14, 481–492. [Google Scholar]
- Faust, E.C. Human helminthology. In A Manual for Clinicians, Sanitarians and Medical Zoologists, 3rd ed.; Lea & Febiger: Philadelphia, PA, USA, 1930; pp. 1–744. [Google Scholar]
- Chai, J.-Y. Miscellaneous Zoonotic Species. In Human Intestinal Flukes; Springer: Dordrecht, The Netherlands, 2019; pp. 491–520. [Google Scholar]
- Varma, T.; Ahluwalia, S. An unusual record of Isoparorchis hypselobagri (Billet 1898), a trematode parasite of fishes from the bile duct of a pig. Indian Vet. J. 1980, 57, 688–689. [Google Scholar]
- Cribb, T. Two new digenetic trematodes from Australian freshwater fishes with notes on previously described species. J. Nat. Hist. 1988, 22, 27–43. [Google Scholar] [CrossRef]
- Reynolds, M.; Duarte, L.M.; Coltro, W.K.T.; Silva, M.F.; Gomez, F.J.V.; Garcia, C.D. Laser-engraved ammonia sensor integrating a natural deep eutectic solvent. Microchem. J. 2020, 157, 105067. [Google Scholar] [CrossRef]
- Smales, L.R.; Barton, D.P.; Chisholm, L.A. Acanthocephalans from Australian elasmobranchs (Chondrichthyes) with a description of a new species in the genus Gorgorhynchus Chandler, 1934 (Rhadinorhynchidae). Syst. Parasitol. 2019, 96, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Bakker, T.C.; Mazzi, D.; Zala, S. Parasite induced changes in behavior and color make Gammarus pulex more prone to fish predation. Ecology 1997, 78, 1098–1104. [Google Scholar] [CrossRef] [Green Version]
- Kaldonski, N.; Perrot-Minnot, M.-J.; Cézilly, F. Differential influence of two acanthocephalan parasites on the antipredator behaviour of their common intermediate host. Anim. Behav. 2007, 74, 1311–1317. [Google Scholar] [CrossRef]
- Perrot-Minnot, M.-J.; Kaldonski, N.; Cézilly, F. Increased susceptibility to predation and altered anti-predator behaviour in an acanthocephalan-infected amphipod. Int. J. Parasitol. 2007, 37, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Taraschewski, H.; Peters, W. Comparative investigations of the morphology and chemical composition of the eggshells of Acanthocephala. Parasitol. Res. 1992, 78, 376–381. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Lui, A.; Squerzanti, S.; Lorenzoni, M.; Shinn, A.P. Confirmation of the hosts involved in the life cycle of an acanthocephalan parasite of Anguilla anguilla (L.) from Lake Piediluco and its effect on the reproductive potential of its amphipod intermediate host. Parasitol. Res. 2012, 110, 2137–2143. [Google Scholar] [CrossRef] [PubMed]
- George, P.V.; Nadakal, A.M. Studies on the life cycle of Pallisentis nagpurensis Bhalerao, 1931 (Pallisentidae; Acanthocephala) parasitic in the fish Ophiocephalus striatus (Bloch). Hydrobiologia 1973, 42, 31–43. [Google Scholar] [CrossRef]
- Blair, D. Polyonchobothrium scleropagis n. sp. (Cestoda: Pseudophyllidea) from the Australian osteoglossid fish Scleropages leichardti Günther. J. Helminthol. 1978, 52, 147–153. [Google Scholar] [CrossRef]
- Shiel, R.; Walker, K.; Williams, W. Plankton of the lower River Murray, South Australia. Mar. Freshw. Res. 1982, 33, 301–327. [Google Scholar] [CrossRef]
- Mitchell, B.; Williams, W. Factors influencing the seasonal occurrence and abundance of the zooplankton in two waste stabilization ponds. Mar. Freshw. Res. 1982, 33, 989–997. [Google Scholar] [CrossRef]
- Geddes, M. Understanding zooplankton communities in farm dams: The importance of predation. In Limnology in Australia; Springer: Berlin/Heidelberg, Germany, 1986; pp. 387–401. [Google Scholar]
- Hołyńska, M. On species of the genus Thermocyclops (Copepoda: Cyclopidae) occurring in northern Queensland, Australia. Ann. Zool. 2006, 56, 335–367. [Google Scholar]
- Chaicharoen, R.; Sanoamuang, L.-O.; Hołyńska, M. A review of the genus Thermocyclops (Crustacea: Copepoda: Cyclopoida) in Cambodia. Zool. Stud. 2011, 50, 780–803. [Google Scholar]
- Gillespie, G.; Hero, J.-M. Potential impacts of introduced fish and fish translocations on Australian amphibians. In Declines and Disappearances of AUSTRALIAN Frogs; Campbell, A., Ed.; Environment Australia: Canberra, Australia, 1999; pp. 131–145. [Google Scholar]




| Fish and Number (N=) | Parasite Species | Site of Infection | No. of Fish Infected | Range in Infected Fish | Prevalence (%) | Total No. of Parasites Found | Mean Intensity | Mean Abundance |
|---|---|---|---|---|---|---|---|---|
| Channa species (n = 103) | Pallisentis sp. Van Cleave, 1928 in total | 96 Pallisentis (P) identified intestinal mesentery and intestinal wall. One Pallisentis (B) embedded in fish musculature | 37 | 0–11 | 35.9 | 97 | 2.62 | 0.94 |
| Pallisentis (B.) sp. 1 Amin, Heckmann, Nguyen, Pham & Pham, 2000 | 1 | 1 | ||||||
| Pallisentis (P.) gomptii Gupta & Verma, 1980 | 5 | 7 | ||||||
| Genarchopsis sp. Ozaki, 1925 total | Free in abdominal cavity | 17 | 0–4 | 16.5 | 36 | 2.1 | 0.34 | |
| Genarchopsis paithanensis Pardeshi & Hiware, 2012 | 3 | 3 | ||||||
| Genarchopsis sp. 1 | 1 | 1 | ||||||
| Senga sp. Dollfus, 1934 | Intestinal lumen | 5 | 0–1 | 04.8 | 5 | 1.0 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, M.; Hernandez-Jover, M.; Shamsi, S. Parasites in Imported Edible Fish and a Systematic Review of the Pathophysiology of Infection and the Potential Threat to Australian Native Aquatic Species. Diversity 2023, 15, 470. https://doi.org/10.3390/d15040470
Williams M, Hernandez-Jover M, Shamsi S. Parasites in Imported Edible Fish and a Systematic Review of the Pathophysiology of Infection and the Potential Threat to Australian Native Aquatic Species. Diversity. 2023; 15(4):470. https://doi.org/10.3390/d15040470
Chicago/Turabian StyleWilliams, Michelle, Marta Hernandez-Jover, and Shokoofeh Shamsi. 2023. "Parasites in Imported Edible Fish and a Systematic Review of the Pathophysiology of Infection and the Potential Threat to Australian Native Aquatic Species" Diversity 15, no. 4: 470. https://doi.org/10.3390/d15040470