
Geographically Isolated Wetlands as a Reserve for the Conservation of Amphibian Biodiversity at the Edge of Their Range


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Wetlands
2.2. Amphibian Recording Methods
2.2.1. Capturing Newt Larvae Using a Hand Net
2.2.2. Visual Counting of Green Water Frogs While Netting Newt Larvae
2.2.3. Bycatch Capturing Amphibians with Fyke Traps
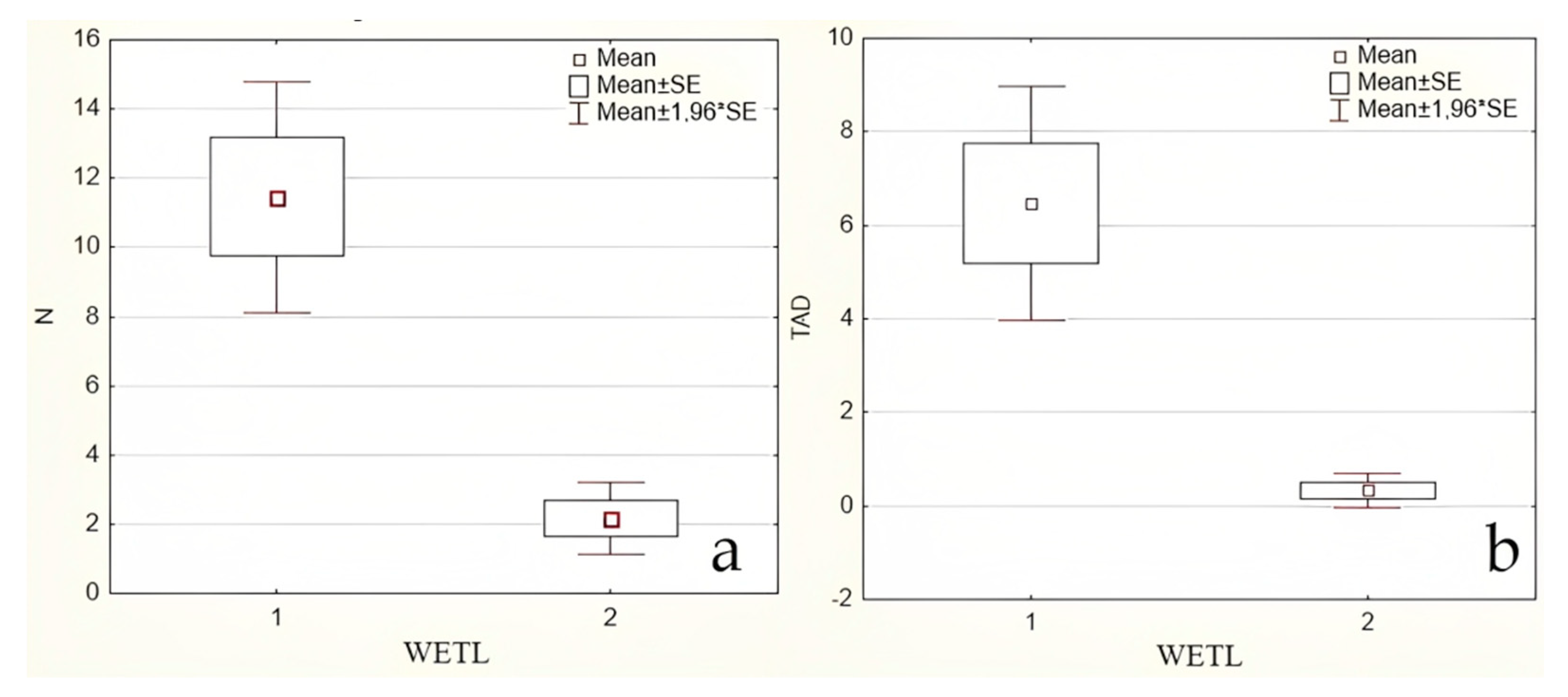
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Muñoz, E.; Gilbert, J.D.; Parra, G.; Guerrero, F. Wetlands classification for amphibian conservation in Mediterranean landscapes. Biodivers. Conserv. 2010, 19, 901–911. [Google Scholar] [CrossRef]
- Beebee, T.J.; Griffiths, R.A. The amphibian decline crisis: A watershed for conservation biology? Biol. Conserv. 2005, 125, 271–285. [Google Scholar] [CrossRef]
- Grant, E.H.C.; Muths, E.; Schmidt, B.R.; Petrovan, S.O. Amphibian conservation in the Anthropocene. Biol. Conserv. 2019, 236, 543–547. [Google Scholar] [CrossRef]
- Pupina, A.; Pupins, M.; Nekrasova, O.; Tytar, V.; Kozynenko, I.; Marushchak, O. Species distribution modelling: Bombina bombina (Linnaeus, 1761) and its important invasive threat Perccottus glenii (Dybowski, 1877) in Latvia under global climate change. J. Environ. Res. Eng. Manag. 2018, 74, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Fernández-Beaskoetxea, S.; Garner, T.W.; Carrascal, L.M. Long-term monitoring of an amphibian community after a climate change-and infectious disease-driven species extirpation. Glob. Chang. Biol. 2018, 24, 2622–2632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tytar, V.; Nekrasova, O.; Pupins, M.; Skute, A.; Marushchak, O.; Čeirāns, A.; Kozynenko, I. Identifying Environmental Refuges (“Coldspots”) from Infection by Batrachochytrium dendrobatidis of Amphibians in Eastern Europe. Biol. Life Sci. Forum 2021, 2, 36. [Google Scholar] [CrossRef]
- Tytar, V.; Nekrasova, O.; Pupins, M.; Čeirāns, A.; Skute, A. Modelling the range expansion of pumpkinseed Lepomis gibbosus across Europe, with a special focus on Ukraine and Latvia. North-West. J. Zool. 2022, 18, 143–150. [Google Scholar]
- Marushchak, O.Y.; Nekrasova, O.D.; Tytar, V.M.; Smirnov, N.A.; Korshunov, O.V.; Pupins, M.; Mykytynets, G.; Skute, A.; Henle, K.; Kaiser, H.A. GIS approach to the study of colour anomalies in amphibians of Ukraine reveals the deleterious effect of human impacts. Herpetol. Notes 2021, 14, 1239–1251. Available online: https://www.biotaxa.org/hn/article/view/62048 (accessed on 1 January 2023).
- Pupins, M.; Nekrasova, O.; Marushchak, O.; Tytar, V.; Theissinger, K.; Čeirāns, A.; Skute, A.; Georges, J.-Y. Potential Threat of an Invasive Fish Species for Two Native Newts Inhabiting Wetlands of Europe Vulnerable to Climate Change. Diversity 2023, 15, 201. [Google Scholar] [CrossRef]
- Nekrasova, O.; Tytar, V.; Pupins, M.; Čeirāns, A. Range expansion of the alien red-eared slider Trachemys scripta (Reptilia, Testudines) in Eastern Europe, with special reference to Latvia and Ukraine. BioInvasions Rec. 2022, 11, 287–295. [Google Scholar] [CrossRef]
- Dubrovsky, Y.V.; Nekrasova, O.D. Vertebrate species richness analysis in pond complexes. In Proceedings of the ZOOCENOSIS–2007: Biodiversity and Role of Animals in Ecosystems. The IV International Conference, Dnipropetrovsk, Ukraine, 9–12 October 2007; Dnipropetrovsk University Press: Dnipropetrovsk, Ukraine; pp. 17–19. (In Russian). [Google Scholar]
- Burrow, A.K.; Lance, S. Restoration of Geographically Isolated Wetlands: An Amphibian-Centric Review of Methods and Effectiveness. Diversity 2022, 14, 879. [Google Scholar] [CrossRef]
- Siffert, O.; Pellet, J.; Ramseier, P.; Tobler, U.; Bergamini, A.; Schmidt, B.R. Where Land and Water Meet: Making Amphibian Breeding Sites Attractive for Amphibians. Diversity 2022, 14, 834. [Google Scholar] [CrossRef]
- Nekrasova, O.D.; Marushchak, O.Y.; Vasyliuk, O.V.; Oskyrko, O.S. Herpetofauna of habitats from Bern Convention Resolution № 4 in Ukraine. In Proceedings of the Conference Proceedings: Classification of Vegetation and Biotopes of Ukraine: The Third Ukrainian Scientific-Theoretical, Kyiv, Ukraine, 19–21 April 2018; Didukh, Y.P., Dubyna, D.V., Eds.; Kholodny Institute of Botany NAS: Kyiv, Ukraine, 2018; pp. 55–63. (In Ukrainian). [Google Scholar]
- Marushchak, O.; Nekrasova, O.; Pupins, M.; Tytar, V.; Ceirans, A. The role and importance of the protected areas’ (Emerald Network) development for amphibians and reptiles on the example of Ukraine in the context of various factors’ influence. J. Environ. Res. Eng. Manag. 2019, 1, 154–158. [Google Scholar] [CrossRef]
- Leibowitz, S.G.; Nadeau, T.L. Isolated wetlands: State-of-the-science and future directions. Wetlands 2003, 23, 663–684. [Google Scholar] [CrossRef]
- Tiner, R.W. Geographically isolated wetlands of the United States. Wetlands 2003, 23, 494–516. [Google Scholar] [CrossRef]
- Nekrasova, O.; Marushchak, O.; Pupins, M.; Tytar, V.; Georges, J.-Y.; Theissinger, K.; Ceirans, A.; Skute, A. Modeling the influence of invasive fish species Perccottus glenii (Dybowski, 1877) on the distribution of newts in Eastern Europe, exemplified by Lissotriton vulgaris (Linnaeus, 1758) and Triturus cristatus (Laurenti, 1768), using a GIS approach. In Proceedings of the 2nd International Electronic Conference on Diversity (IECD 2022)—New Insights into the Biodiversity of Plants, Animals and Microbes, 15–31 March 2022; MDPI: Basel, Switzerland, 2022. [Google Scholar] [CrossRef]
- Rannap, R.; Lõhmus, A.; Briggs, L. Restoring Ponds for Amphibians: A Success Story. Hydrobiologia 2009, 634, 87–95. [Google Scholar] [CrossRef]
- Catenazzi, A. State of the world's amphibians. Annu. Rev. Environ. Resour. 2015, 40, 91–119. [Google Scholar] [CrossRef]
- Heyer, R.; Donnelly, M.A.; Foster, M.; Mcdiarmid, R. (Eds.) Measuring and Monitoring Biological Diversity: Standard Methods for Amphibians; Smithsonian Institution: Washington, DC, USA, 2014; p. 320. [Google Scholar]
- Čeirāns, A.; Pupins, M. Amphibian and Reptile Background Monitoring Manual, 2nd ed.; Latgale Ecological Society; Daugavpils University: Daugavpils, Latvia, 2020; 34p. (In Latvian) [Google Scholar]
- Čeirāns, A.; Pupins, M. Amphibian and Reptile Natura 2000 Monitoring Manual, 2nd ed.; Latgale Ecological Society; Daugavpils University: Daugavpils, Latvia, 2020; 29p. (In Latvian) [Google Scholar]
- Pupins, M.; Čeirāns, A.; Nekrasova, O.; Theissinger, K.; Georges, J.-Y. Method of collecting green frogs for scientific and environmental studies by hand net catching. Daugavpils Latg. Ecol. Soc. 2022, 1–11. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Calderon, M.R.; González, S.P.; Pérez-Iglesias, J.M.; Jofré, M.B. Anthropogenic impacts on rivers: Use of multiple indicators to assess environmental quality status. Hydrobiologia 2022, 850, 469–487. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wetland Number | Coordinates | Dimensions | Vegetation (% of Area) | Natural Wetland Destroying History (WD) and the Method Used to Restore the Wetland (WR) |
|---|---|---|---|---|
| LV.1.1. | 55.692844; 26.786556 | L = 140 m; B = 9 m; S = 1383 m2, Dmax = 1.2 m | E = 40; SE = 60; F = 70. | WD: Natural wetland has been drained, its catchment basin was used for agriculture and polluted with fertilizers and overgrown with shrubs. WR: Pond LV.1.1. was reconstructed by excavation in 2013 in a wetland natural area; ponds LV.1.2.–LV.2.4. in 2018. |
| LV.1.2. | 55.693658; 26.788077 | L = 19 m; B = 12 m; S = 207 m2; Dmax = 1 m | E = 20; SE = 60; F = 100. | WD, WR: See LV.1.1. |
| LV.1.3. | 55.694307; 26.789198 | L = 6 m; B = 5 m; S = 28 m2; Dmax = 1 m | E = 5; SE = 10; F = 5. | WD, WR: See LV.1.1. |
| LV.1.4. | 55.695204; 26.790489 | L = 23 m; B = 5 m; S = 79 m2; Dmax = 1 m | E = 30; SE = 20; F = 10. | WD, WR: See LV.1.1. |
| LV.2.1. | 55.691701; 26.790466 | L = 120 m; B = 2 m; S = 1587 m2; Dmax = 1.5 m | E = 70; SE = 60; F = 30. | WD: Natural wetland has been drained; its catchment basin was used for agriculture and heavily polluted with fertilizers; therefore, the drained wetland was overgrown with reed. WR: Ponds LV.2.1.–LV.2.8. were reconstructed by excavation in 2013 in a wetland natural area. |
| LV.2.2. | 55.691000; 26.789041 | L = 30 m; B = 10 m; S = 297 m2; Dmax = 1 m | E = 20; SE = 70; F = 10. | WD, WR: See LV.2.1. |
| LV.2.3. | 55.690848; 26.788735 | L = 25 m; B = 12 m; S = 272 m2; Dmax = 1 m | E = 10; SE = 70; F = 10. | WD, WR: See LV.2.1. |
| LV.2.4. | 55.690598; 26.788258 | L = 24 m; B = 11 m; S = 272 m2; Dmax = 1 m | E = 20; SE = 60; F = 5. | WD, WR: See LV.2.1. |
| LV.2.5. | 55.690268; 26.787862 | L = 50 m; B = 11 m; S = 678 m2; Dmax = 1 m | E = 10; SE = 40; F = 5. | WD, WR: See LV.2.1. |
| LV.2.6. | 55.689822; 26.787572 | L = 34 m; B = 12 m; S = 472 m2; Dmax = 1 m | E = 30; SE = 80; F = 30. | WD, WR: See LV.2.1. |
| LV.2.7. | 55.689392; 26.787220 | L = 54 m; B = 12 m; S = 754 m2; Dmax = 1 m | E = 20; SE = 40; F = 10. | WD, WR: See LV.2.1. |
| LV.2.8. | 55.688899; 26.786974 | L = 20 m; B = 10 m; S = 196 m2; Dmax = 1 m | E = 40; SE = 80; F = 40. | WD, WR: See LV.2.1. |
| LV.3.1. | 55.684114; 26.773356 | L = 8 m; B = 7 m; S = 66 m2; Dmax = 1 m | E = 40; SE = 70; F = 5. | WD: Natural wetland with a stream connected with Lake Sita was drained and overgrown with shrubs due to pollution from a nearby (~200 m) pig farm. WR: Ponds LV.3.1. and LV.3.7. were excavated in 2013 in a wetland natural area; ponds LV.3.8.–LV.3.10. in 2018. |
| LV.3.2. | 55.684200; 26.773139 | L = 29 m; B = 5 m; S = 121 m2; Dmax = 1 m | E = 20; SE = 80; F = 100. | WD, WR: See LV.3.1. |
| LV.3.3. | 55.684408; 26.771872 | L = 30 m; B = 6 m; S = 197 m2; Dmax = 1.5 m | E = 30; SE = 70; F = 80. | WD, WR: See LV.3.1. |
| LV.3.4. | 55.684552; 26.770932 | L = 11 m; B = 7 m; S = 70 m2; Dmax = 1 m | E = 90; SE = 80; F = 70. | WD, WR: See LV.3.1. |
| LV.3.5. | 55.684549; 26.770327 | L = 8 m; B = 7 m; S = 49 m2; Dmax = 0.6 m | E = 70; SE = 100; F = 80. | WD, WR: See LV.3.1. |
| LV.3.6. | 55.684718; 26.769991 | L = 25 m; B = 8 m; S = 170 m2; Dmax = 1.5 m | E = 30; SE = 60; F = 30. | WD, WR: See LV.3.1. |
| LV.3.7. | 55.684942; 26.769197 | L = 58 m; B = 6 m; S = 339 m2; Dmax = 1,5 m | E = 30; SE = 80; F = 90. | WD, WR: See LV.3.1. |
| LV.3.8. | 55.685713; 26.767474 | L = 18 m; B = 9 m; S = 226 m2; Dmax = 1 m | E = 5; SE = 10; F = 20. | WD, WR: See LV.3.1. |
| LV.3.9. | 55.685746; 26.768066 | L = 22 m; B = 11 m; S = 239 m2; Dmax = 1 m | E = 10; SE = 20; F = 5. | WD, WR: See LV.3.1. |
| LV.3.10. | 55.685665; 26.768568 | L = 20 m; B = 9 m; S = 209 m2; Dmax = 1 m | E = 20; SE = 40; F = 5. | WD, WR: See LV.3.1. |
| LV.4.1. | 55.690270; 26.776341 | L = 56 m; B = 10 m; S = 926 m2; Dmax = 1.5 m | E = 20; SE = 30; F = 5. | WD: Natural wetland was drained. WR: Pond LV.4.1. was excavated in 2018; pond LV.4.2. in 2013; and ponds LV.4.3–LV.4.5. in 2006. |
| LV.4.2. | 55.690796; 26.772181 | L = 41 m; B = 15 m; S = 674 m2; Dmax = 1.5 m | E = 20; SE = 90; F = 70. | WD, WR: See LV.4.1. |
| LV.4.3. | 55.690813; 26.771567 | L = 20 m; B = 12 m; S = 242 m2; Dmax = 0,5 m | E = 70; SE = 20; F = 30. | WD, WR: See LV.4.1. |
| LV.4.4. | 55.690868; 26.770978 | L = 12 m; B = 9 m; S = 106 m2; Dmax = 0.5 m | E = 80; SE = 20; F = 5. | WD, WR: See LV.4.1. |
| LV.4.5. | 55.691192; 26.770954 | L = 31 m; B = 12 m; S = 376 m2; Dmax = 1 m | E = 20; SE = 80; F = 100. | WD, WR: See LV.4.1. |
| Ponds | N Amphibians (Mean/Pond) | T. cristatus Larvae (Mean/Pond) | L. vulgaris Larvae (Mean/Pond) | Pelophylax (Mean/Pond) | N Ponds | N Ponds with Fish |
|---|---|---|---|---|---|---|
| 2018 GIW | 62 (12.4) | 21 (4.2) | 7 (1.4) | 34 (6.8) | 5 | 0 |
| 2018 nGIW | 35 (7) | 8 (1.6) | 8 (1.6) | 19 (3.8) | 5 | 1 |
| 2018 All | 97 | 29 | 15 | 53 | 10 | 1 |
| 2022 GIW | 115 (14.4) | 21 (2.6) | 64 (8) | 30 (3.75) | 8 | 0 |
| 2022 nGIW | 33 (5.5) | 1 (0.2) | 9 (1.5) | 23 (3.83) | 6 | 6 |
| 2022 All | 148 | 22 | 73 | 53 | 14 | 6 |
| All | 245 | 51 | 88 | 106 | 24 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pupins, M.; Nekrasova, O.; Tytar, V.; Garkajs, A.; Petrov, I.; Morozova, A.; Theissinger, K.; Čeirāns, A.; Skute, A.; Georges, J.-Y. Geographically Isolated Wetlands as a Reserve for the Conservation of Amphibian Biodiversity at the Edge of Their Range. Diversity 2023, 15, 461. https://doi.org/10.3390/d15030461
Pupins M, Nekrasova O, Tytar V, Garkajs A, Petrov I, Morozova A, Theissinger K, Čeirāns A, Skute A, Georges J-Y. Geographically Isolated Wetlands as a Reserve for the Conservation of Amphibian Biodiversity at the Edge of Their Range. Diversity. 2023; 15(3):461. https://doi.org/10.3390/d15030461
Chicago/Turabian StylePupins, Mihails, Oksana Nekrasova, Volodymyr Tytar, Alberts Garkajs, Iurii Petrov, Aleksandra Morozova, Kathrin Theissinger, Andris Čeirāns, Arturs Skute, and Jean-Yves Georges. 2023. "Geographically Isolated Wetlands as a Reserve for the Conservation of Amphibian Biodiversity at the Edge of Their Range" Diversity 15, no. 3: 461. https://doi.org/10.3390/d15030461