
Impact of No-Tillage on Soil Invertebrate Communities in the Southern Forest Steppe of West Siberia: Preliminary Research
 , , and
, , and
Abstract
:1. Introduction
2. Object of Study, Material and Methods
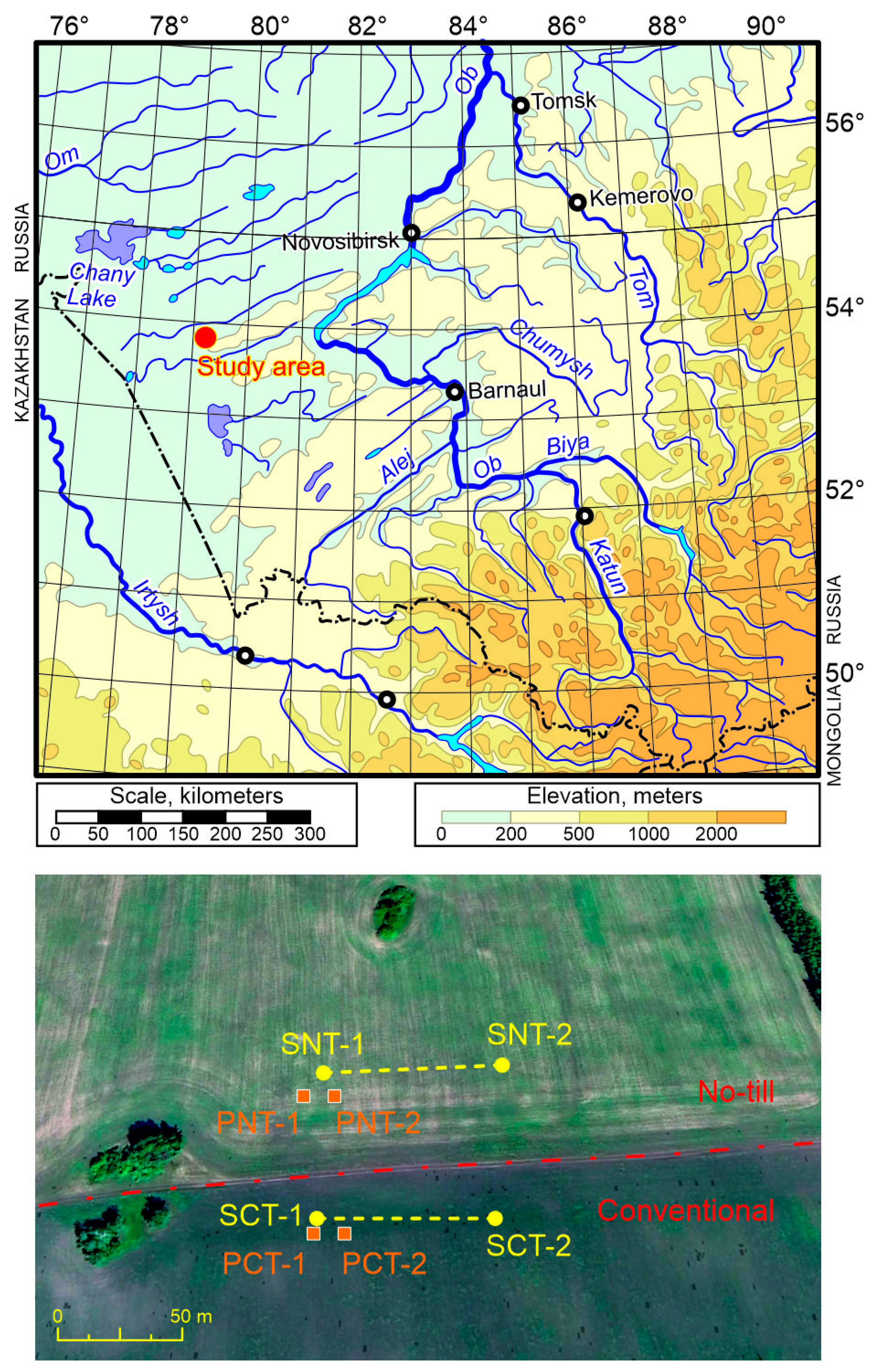
2.1. Experimental Sites and Soils of the Agrocenoses Studied
2.2. Sampling of Soil Invertebrates and Processing
- Taking soil samples to assess communities of soil-inhabiting invertebrates (earthworms, soil arthropods, etc.). We took soil monoliths 50 cm ∗ 25 cm in size (area 1/8 m2) to a depth of 15 cm in 10-fold replication. Soil samples were taken along transects about 70 m long, passing at a distance of 20–40 m from the field boundaries; the distance between individual samples was about 5–8 m (Figure 2). Samples were taken twice (22–24 May 2017 and 15 May 2018). Samples were placed in ventilated bags made of synthetic fabric, delivered to the laboratory, and disassembled by hand a short time after being obtained. During manual sorting of samples, if possible, all representatives of the macrofauna larger than 1–2 mm were extracted from them. Springtails and small soil mites such as Oribatida (soil mesofauna) were not taken into account.
- 2.
- Installation of pitfall traps for the study of the soil-surface arthropods (carabids, spiders, etc.). A total of 10 plastic cups with a diameter of 6.5 cm (1/3 filled with a 3% solution of acetic acid used as a fixing liquid) were exhibited at both plots from 22–30 May 2017 (before sowing) and 13–20 June 2018 (after emergence of crops, but before insecticidal treatment, see Table 1). Short-duration pitfall trapping was used due to the rapid change in fauna and the developmental stages of ground beetles and other soil invertebrates during spring and summer. A long recording period contributes to the registration of species random for the habitat [15,16]. The change in fauna on agricultural fields is determined not only by the peculiarities of the phenology of specific species, but also by the sequence of agrotechnical measures (for example, the use of pesticides), which radically change the structure of soil animal communities. Traps were placed about 20–40 m from the edge of the field in two groups of five in the form of a square “envelope” with a central trap (five-spot pattern). The length of one side of the square was about 4 m. The distance between the groups of traps was about 7–10 m (see Figure 2). Thus, the location of the traps was a rectangle approximately 4 ∗ 15 m, inside which 10 traps were placed in a checkerboard pattern. We did not use trap covers in order to make the results comparable to our previous studies. Our 25 years of experience shows that traps of such a small diameter, standing on a flat surface, are almost never flooded, even with heavy rain. The traps’ short exposure period also avoids flooding. If traps are shaded with covers, they become more attractive in the open field as a refuge for soil-dwelling animals and thus distort the counting results.
- 3.
- All invertebrates from soil samples were placed in 70% alcohol. They were determined in order or (in the case of Coleoptera) to families in the laboratory. Invertebrates of the most numerous groups (earthworms (Lumbricidae), spiders (Aranei), ground beetles (Carabidae), and click beetles (Elateridae)) were determined at the species level.
- 4.
- After exposure, pitfall traps were placed in individual zip bags and delivered to the laboratory, where spiders and beetles were extracted from them. Ground beetles and spiders were identified according to species.
- 5.
- Earthworm species were identified by E.V. Golovanova according to the key to the earthworms of Russia [17]. DNA was extracted from ethanol-fixed specimens. A sample of the body wall (about 100 µg) was dissolved in guanidiniumisothiocyanate and transferred to silica columns (BioSilica, Russia). After several rounds of washing with guanidiniumisothiocyanate and ethanol, DNA was transferred to a clean tube and used as a PCR matrix. Amplification of the mitochondrial cox1 gene fragment was conducted using universal primers LCO1490m (5′-TACTC-AACAA-ATCAC-AAAGA-TATTG-G-3′) and HCO2198 (5′-TAAAC-TTCAG-GGTGA-CCAAA-AAATC-A-3′) [18] with the Biomaster HS-Taq PCR Mix (Biolabmix, Russia). Sequencing was performed in the SB RAS Genomics Core Facility (ICBFM SB RAS, Novosibirsk, Russia). Sequences were edited using Chromas 2.6.6 (http://technelysium.com.au, accessed on 6 March, 2023). Unique haplotypes were submitted to GenBank under accession numbers OQ271318 and OQ271319.
2.3. Statistics
3. Results
3.1. Soils
3.2. Common Features of Soil Invertebrate Communities
3.3. Analysis of the Diversity and Abundance of Soil Invertebrate Communities
3.4. Carabidae
3.5. Spiders
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kutovaya, O.V.; Nikitin, D.A.; Geraskina, A.P. No-till technology as a factor of activity of soil invertebrate in agricultural chernozems of Stavropol region. Sel’skokhozyaistvennaya Biol. Agric. Biol. 2021, 56, 199–210. (In Russian) [Google Scholar] [CrossRef]
- House, G.J.; Parmelee, R.W. Comparison of soil arthropods and earthworms from conventional and no-tillage agroecosystems. Soil Tillage Res. 1985, 5, 351–360. [Google Scholar] [CrossRef]
- Errouissi, F.; Ben Moussa-Machraoui, S.; Ben-Hammouda, M.; Nouira, S. Soil invertebrates in durum wheat (Triticum durum L.) cropping system under Mediterranean semi arid conditions: A comparison between conventional and no-tillage management. Soil Tillage Res. 2011, 112, 122–132. [Google Scholar] [CrossRef]
- Winter, J.P.; Voroney, R.P.; Ainsworth, D.A. Soil microarthropods in long-term no-tillage and conventional tillage corn production. Can. J. Soil Sci. 1990, 70, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Bedano, J.C.; Cantú, M.P.; Doucet, M.E. Influence of three different land management practices on soil mite (Arachnida: Acari) densities in relation to a natural soil. Appl. Soil Ecol. 2006, 32, 293–304. [Google Scholar] [CrossRef]
- Warburton, D.B.; Klimstra, W.D. Wildlife use of no-till and conventionally tilled corn fields. J. Soil Water Conserv. 1984, 39, 327–330. [Google Scholar]
- Robertson, L.; Kettle, B.; Simpson, G. The influence of tillage practices on soil macrofauna in a semi-arid agroecosystem in northeastern Australia. Agric. Ecosyst. Environ. 1994, 48, 149–156. [Google Scholar] [CrossRef]
- Hendrix, P.F.; Parmelee, R.W.; Crossley, D.A.; Coleman, D.C.; Odum, E.P.; Groffman, P. Detritus Food Webs in Conventional and No-Tillage Agroecosystems. Bioscience 1986, 36, 374–380. [Google Scholar] [CrossRef]
- Lins, V.S.; Santos, H.R.; Gonçalves, M.C. The effect of the glyphosate, 2,4-D, atrazine e nicosulfuron herbicides upon the Edaphic collembola (Arthropoda: Ellipura) in a no tillage system. Neotropical Entomol. 2007, 36, 261–267. [Google Scholar] [CrossRef] [Green Version]
- House, G.J.; Worsham, A.D.; Sheets, T.J.; Stinner, R.E. Herbicide effects on soil arthropod dynamics and wheat straw decom-position in a North Carolina no-tillage agroecosystem. Biol. Fertil. Soils 1987, 4, 109–114. [Google Scholar] [CrossRef]
- Tabaglio, V.; Gavazzi, C.; Menta, C. Physico-chemical indicators and microarthropod communities as influenced by no-till, conventional tillage and nitrogen fertilisation after four years of continuous maize. Soil Tillage Res. 2009, 105, 135–142. [Google Scholar] [CrossRef]
- Bokina, I.G. Bugs (Heteroptera, Nabidae) in agrocenoses of cereals in the forest-steppe of Western Siberia. Zool. Zhurnal 2013, 92, 450–456. [Google Scholar] [CrossRef]
- Bokina, I.G. Lacewing insects (Neuroptera: Chrysopidae, Hemerobiidae) Neuroptera insects (Chrysopidae, Hemerobiidae) according to different spring wheat cultivation practices in the northern forest-steppe of Western Siberia. Euroasian Entomol. J. 2020, 19, 67–74. [Google Scholar] [CrossRef]
- IUSS Working Group. WRB, World Reference Base for Soil Resources 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2015. [Google Scholar]
- Matalin, A.V. Peculiarities of Spatial and Temporal Differentiation of Carabids (Coleoptera, Carabidae) in the Steppe Zone. Entomol. Rev. 1997, 77, 1155–1166. [Google Scholar]
- Matalin, A.V.; Makarov, K.V. Using demographic data to better interpret pitfall trap catches. Zookeys 2011, 100, 223–254. [Google Scholar] [CrossRef]
- Vsevolodova-Perel, T.S. Dozhdevye Chervi Fauny Rossii. Kadastr i Opredelitel [The Earthworms of the Fauna of Russia. Cadaster and Key]; Nauka: Moscow, Russia, 1997; 102p. [Google Scholar]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. Conserved primers for PCR amplification of mitochondrial DNA from different invertebrate phyla. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Mamaev, B.M. , Medvedev, L.N., Pravdin, F.N. Opredelitel Nasekomykh Evropeiskoi Chasti SSSR [Identification Keys to the insects of the European part of the USSR]; Prosveschenie: Moscow, Russia, 1976; 304p. (In Russian) [Google Scholar]
- Dolin, V.G. Opredelitel Lichinok Zhukov-Stchelkunov Fauny SSSR [Identification Keys to the Elateridae Larvae of the Fauna of the USSR]; Urozhai: Kiev, Russia, 1978; 124p. [Google Scholar]
- Byzova, Y.B.; Ghilyarov, M.S.; Dunger, V.; Zakharov, A.A.; Kozlovskaya, L.S.; Korganova, G.A.; Mazantseva, G.P.; Meletsis, V.I.; Prasse, I.; Puzachenko, J.G.; et al. Kolichestvennye Metody v Pochvennoi Zoologii [Quantitative Methods in Soil Zoology]; Nauka: Moscow, Russia, 1987; 288p. (In Russian) [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer: Dordrecht, The Netherlands, 1988. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Carter, M.R.; Gregorich, E.G. (Eds.) Soil Sampling and Methods of Analysis, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Shekhovtsov, S.; Golovanova, E.; Peltek, S. Cryptic diversity within the Nordenskiold's earthworm, Eisenia nordenskioldi subsp. nordenskioldi (Lumbricidae, Annelida). Eur. J. Soil Biol. 2013, 58, 13–18. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Golovanova, E.; Peltek, S.E. Genetic diversity of the earthworm Eisenia nordenskioldi (Lumbricidae, Annelida). Vavilov J. Genet. Breed. 2017, 21, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Shekhovtsov, S.V.; Golovanova, E.V.; Peltek, S.E. Mitochondrial DNA variation in Eisenia n. nordenskioldi (Lumbricidae) in Europe and Southern Urals. Mitochondrial DNA Part A 2015, 27, 4643–4645. [Google Scholar] [CrossRef] [PubMed]
- Shekhovtsov, S.V.; Berman, D.I.; Bulakhova, N.A.; Vinokurov, N.N.; Peltek, S.E. Phylogeography of Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) from the north of Asia. Polar Biol. 2018, 41, 237–247. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Shipova, A.A.; Poluboyarova, T.V.; Vasiliev, G.V.; Golovanova, E.V.; Geraskina, A.P.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Species Delimitation of the Eisenia nordenskioldi Complex (Oligochaeta, Lumbricidae) Using Transcriptomic Data. Front. Genet. 2020, 11, 598196. [Google Scholar] [CrossRef] [PubMed]
- Perel, T.S. Distribution and Patterns of Distribution of Earthworms in the Fauna of the USSR (with Key Tables); Nauka: Moscow, Russia, 1979. [Google Scholar]
- Golovanova, E.V.; Kniazev, S.Y.; Babiy, K.A.; Tsvirko, E.I.; Karaban, K.; Solomatin, D.V. Dispersal of earthworms from the Rudny Altai (Kazakhstan) into Western Siberia. Ecol. Montenegrina 2021, 45, 48–61. [Google Scholar] [CrossRef]
- Kniazev, S.Y.; Kislyi, A.A.; Bogomolova, I.N.; Golovanova, E.V. Territorial Heterogeneity of the Earthworm Population (Opisthopora, Lumbricidae) of Omsk Oblast and Environmental Factors: A Quantitative Assessment of the Relationship. Contemp. Probl. Ecol. 2022, 15, 484–493. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Conventional Farming System | No-Till Farming System |
|---|---|---|
| Geographical coordinates | Soil samples: SCT1—53.81663° N, 79.24866° E; SCT2—53.81663° N, 79.24980° E. Pitfall traps: PCT1—53.81660° N, 79.24866° E; PCT2—53.81659° N, 79.24885° E. | Soil samples: SNT1—53.81732° N, 79.24862° E; SNT2—53.81731° N, 79.24992° E. Pitfall traps: PNT1—53.81718° N, 79.24850° E; PNT2—53.81718° N, 79.24871°E. |
| Soil tillage | Dump plowing to a depth of 18–20 cm, moisture nailing with harrows and spring pre-sowing cultivation to a depth of 10 cm | Complete absence of tillage, starting from 2007 |
| Cultivated crop in 2017 | Spring wheat | Spring Wheat |
| Cultivated crop in 2018 | Peas | Rape |
| Fertilization, annually in the period (more than 3 years) prior to sampling | Without fertilizers | 35–42 kgN/ha (ammoniumnitrate) and 10 kgN/ha + 26 kgP2O5/ha + 26 K2O/ha (di-ammoniumphosphate) |
| Application of pesticides immediately before sampling in 2017 | Was not applied | Was not applied |
| Application of pesticides in 2017 | Elant herbicides 0.6 L/ha + Stalker 0.02 kg/ha + Taipan 0.5 L/ha during the growing season | Kernell herbicides 2 L/ha + Esteron 0.5 L/ha before sowing; herbicides Agrokson 0.6 L/ha + Trizlak 0.02 kg/ha + Foxtrot Extra 0.5 L/ha during the growing season; fungicide Abakus Ultra 1 L/ha; insecticide Fastak 0.1 L/ha |
| Application of pesticides immediately before sampling in 2018 | Was not applied | Was not applied until the end of May, at the beginning of June Kernell herbicides 2 L/ha + Esteron 0.5 L/ha |
| Year | 2017 | 2018 | ||
|---|---|---|---|---|
| Type of Soil Treatment | No-Till | Conventional | No-Till | Conventional |
| Diversity Parameter | ||||
| Simpson_1-D | 0.79 | 0.80 | 0.75 ** | 0.46 ** |
| Shannon_H | 1.87 | 1.99 | 1.74 ** | 1.06 ** |
| Evenness_e^H/S | 0.50 | 0.49 | 0.44 | 0.36 |
| Menhinick | 1.02 | 1.38 | 0.90 | 1 |
| Margalef | 2.36 | 2.94 | 2.24 | 1.68 |
| Equitability_J | 0.73 | 0.74 | 0.68 * | 0.51 * |
| Fisher_alpha | 3.33 | 4.56 | 3.06 | 2.41 |
| Berger–Parker | 0.35 | 0.35 | 0.42 ** | 0.72 ** |
| Year | 2017 | 2018 | ||
|---|---|---|---|---|
| Type of Soil Treatment | No-Till | Conventional | No-Till | Conventional |
| Taxa, Functional Group | ||||
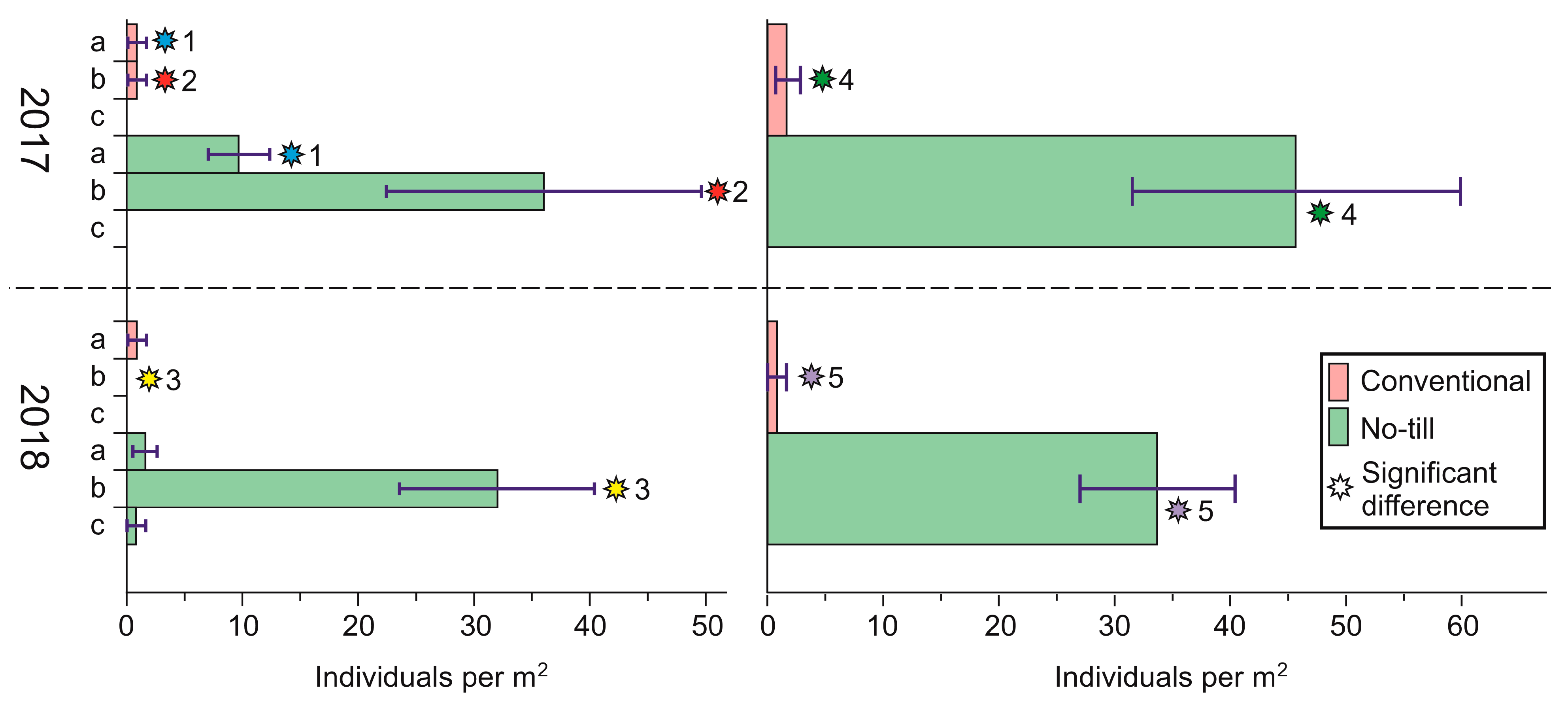
| Enchytraeidae | 8.80 ± 4.21 | 10.40 ± 7.16 | 71.20 ± 34.51 * | 6.40 ± 1.07 * |
| Lumbricidae | 45.60 ± 14.16 ** | 1.60 ± 1.07 ** | 33.60 ± 6.73 ** | 0.80 ± 0.80 ** |
| Trombidiformes | 1.60 ± 1.07 | 4.80 ± 2.13 | 0 | 0 |
| Aranei | 21.60 ± 5.73 ** | 0.80 ± 0.80 ** | 0.80 ± 0.80 | 0 |
| Chilopoda | 0.80 ± 0.80 | 0 | 5.60 ± 2.93 | 0 |
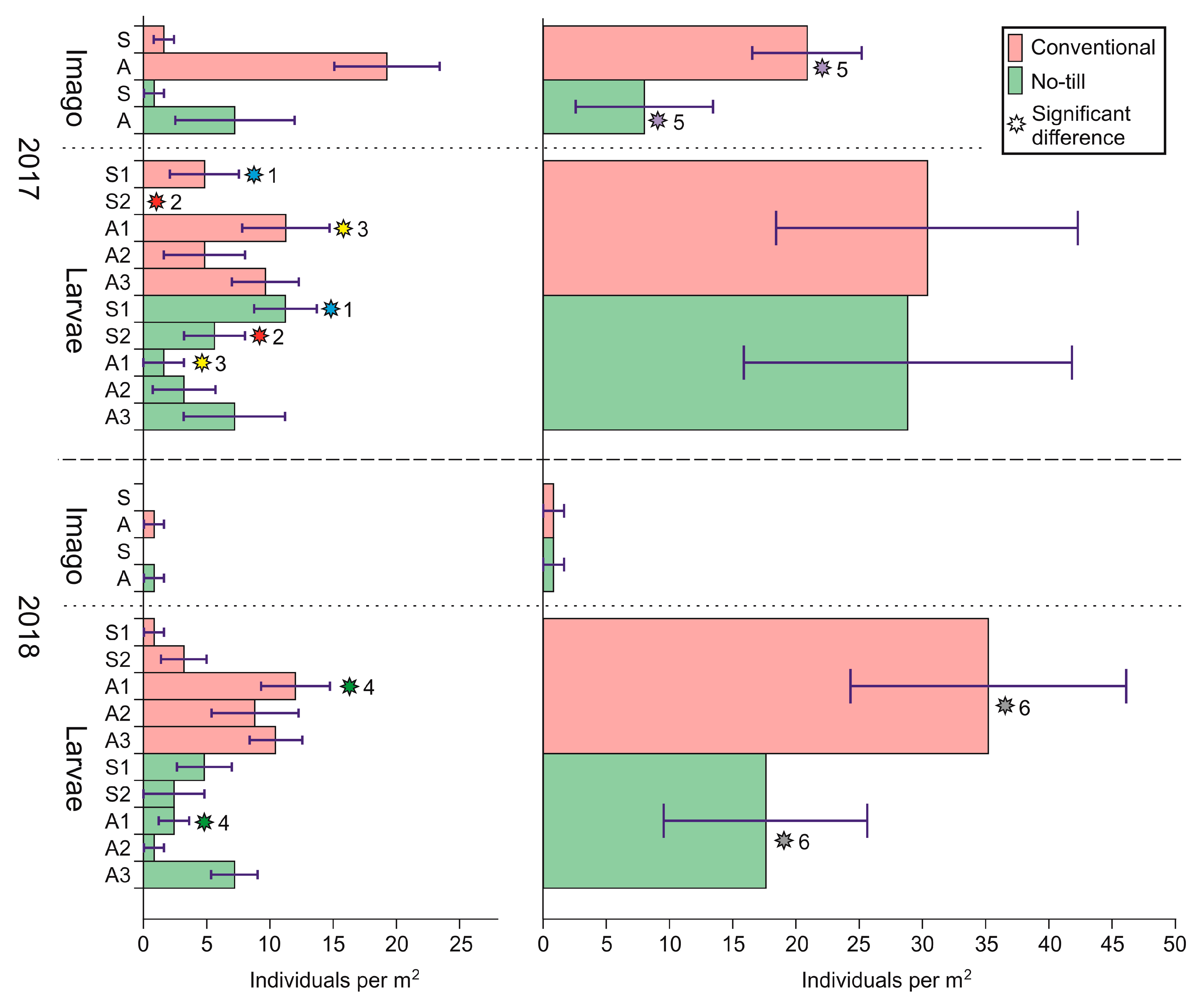
| Carabidae | 8.80 ± 3.26 | 11.20 ± 6.10 | 6.40 ± 2.61 | 1.60 ± 1.07 |
| Carabidae. l | 1.60 ± 1.07 | 0 | 16.00 ± 4.77 ** | 1.60 ± 1.07 ** |
| Elateridae | 8.00 ± 5.47 * | 20.80 ± 4.33 * | 0.80 ± 0.80 | 0.80 ± 0.80 |
| Elateridae. l | 31.20 ± 8.55 | 32.80 ± 10.22 | 17.60 ± 4.89 * | 36.80 ± 5.49 * |
| Staphylinidae | 3.20 ± 1.77 | 2.40 ± 1.71 | 0 | 0 |
| Curculionidae | 2.40 ± 1.71 | 2.40 ± 1.22 | 0.80 ± 0.80 | 1.60 ± 1.07 |
| Curculionidae. l | 0 | 1.60 ± 1.07 | 0 | 0 |
| Other Coleoptera | 0 | 0.80 ± 0.80 | 1.60 ± 1.07 | 0 |
| Other Coleoptera. l | 0 | 2.40 ± 1.22 | 0.80 ± 0.80 | 0 |
| Hymenoptera | 0.8 ± 0.8 | 0.80 ± 0.80 | 12.00 ± 7.93 | 0 |
| Hemiptera | 0.8 ± 0.8 | 0 | 0.80 ± 0.80 | 0 |
| Diptera. l et pupae | 0.8 ± 0.8 | 0.80 ± 0.80 | 0 | 1.60 ± 1.60 |
| Lepidoptera. l | 0 | 0.80 ± 0.80 | 0 | 0 |
| Number of individuals per square meter | 136.00 ± 22.28 | 94.40 ± 17.72 | 168.00 ± 42.88 ** | 51.2 ± 6.00 ** |
| Number of taxa per one soil sample | 5.30 ± 0.56 | 4.90 ± 0.48 | 5.10 ± 0.48 ** | 2.70 ± 0.33 ** |
| Year | 2017 | 2018 | ||
|---|---|---|---|---|
| Type of Soil Treatment | No-Till | Conventional | No-Till | Conventional |
| Diversity Parameter | ||||
| Simpson_1-D | 0.81 ** | 0.49 ** | 0.54 | 0.44 |
| Shannon_H | 1.80 ** | 0.68 ** | 1.13 | 0.64 |
| Evenness_e^H/S | 0.86 | 0.99 | 0.52 | 0.94 |
| Menhinick | 2.11 ** | 0.53 ** | 1.11 | 1.16 |
| Margalef | 2.50 ** | 0.38 ** | 1.49 | 0.91 |
| Equitability_J | 0.92 | 0.99 | 0.63 | 0.92 |
| Fisher_alpha | 8.29 * | 0.64 * | 2.30 | 2.62 |
| Berger–Parker | 0.27 ** | 0.57 ** | 0.66 | 0.67 |
| Year | 2017 | 2018 | ||
|---|---|---|---|---|
| Type of Soil Treatment | No-Till | Conventional | No-Till | Conventional |
| Carabid Species | ||||
| Amara consularis | 2.40 ± 2.40 | 0 | 2.40 ± 1.71 | 0 |
| Amara consularis, l | 0 | 0 | 15.20 ± 4.69 ** | 0 ** |
| Bembidion properans | 0.80 ± 0.80 | 4.80 ± 3.20 | 0.80 ± 0.80 | 0.80 ± 0.80 |
| Bembidion varium | 0.80 ± 0.80 | 0 | 0 | 0 |
| Bembidion quadrimaculatum | 0 * | 6.40 ± 3.33 * | 0 | 0 |
| Calathus melanocephalus | 0.80 ± 0.80 | 0 | 0 | 0 |
| Harpalus rufipes | 0.80 ± 0.80 | 0 | 0 | 0 |
| Poecilus cupreus | 2.40 ± 1.71 | 0 | 3.20 ± 1.31 * | 0 * |
| Poecilus cupreus, l | 0 | 0 | 0.80 ± 0.80 | 1.60 ± 1.07 |
| Poecilus fortipes | 0 | 0 | 0.80 ± 0.80 | 0 |
| Poecilus versicolor | 0.80 ± 0.80 | 0 | 0 | |
| Number of individuals per square meter | 8.80 ± 3.26 | 11.20 ± 6.10 | 23.20 ± 4.21 ** | 2.40 ± 1.22 ** |
| Number of species per one soil sample | 0.80 ± 0.33 | 0.70 ± 0.21 | 1.70 ± 0.26 ** | 0.20 ± 0.13 ** |
| Year | 2017. May | 2018. June | ||
|---|---|---|---|---|
| Type of Soil Treatment | Conventional | No-Till | Conventional | No-Till |
| Species | ||||
| Agonum gracilipes | 0.10 ± 0.10 | 0.00 | 0.30 ± 0.15 | 0.00 |
| Amara consularis | 0.00 | 0.10 ± 0.10 | 0.00 | 0.56 ± 0.29 |
| Amara apricaria | 0.00 | 0.00 | 0.00 | 0.22 ± 0.15 |
| Amara bifrons | 0.00 | 0.00 | 0.10 ± 0.10 | 0.00 |
| Anisodactylus signatus | 0.00 | 0.00 | 0.20 ± 0.13 | 0.00 |
| Bembidion properans | 3.70 ± 1.15 * | 0.30 ± 0.15 * | 17.80 ± 3.97 * | 1.33 ± 0.50 * |
| Bembidion quadrimaculatum | 3.90 ± 1.05 * | 0.10 ± 0.10 * | 3.60 ± 1.09 * | 0.33 ± 0.17 * |
| Bembidion gilvipes | 0.00 | 0.00 | 0.10 ± 0.10 | 0.00 |
| Calosoma denticolle | 0.00 | 0.00 | 0.10 ± 0.10 | 0.00 |
| Carabus convexus | 0.00 | 0.00 | 0.10 ± 0.10 | 0.00 |
| Carabus sibiricus | 0.10 ± 0.10 | 0.00 | 0.00 | 0.00 |
| Curtonotus aulicus | 0.00 | 0.00 | 0.10 ± 0.10 | 0.00 |
| Harpalus cisteloides | 0.00 | 0.00 | 0.30 ± 0.21 | 0.00 |
| Microlestes minutulus | 0.20 ± 0.13 | 0.20 ± 0.13 | 0.60 ± 0.27 | 0.33 ± 0.24 |
| Notiophilus germinyi | 0.10 ± 0.10 | 0.00 | 0.00 | 0.00 |
| Poecilus cupreus | 23.00 ± 4.07 | 24.70 ± 6.85 | 216.00 ± 26.98 * | 0.11 ± 0.11 * |
| Poecilus fortipes | 1.00 ± 0.37 | 0.20 ± 0.13 | 16.20 ± 1.20 * | 0.00* |
| Poecilus lepidus | 0.00 | 0.00 | 0.30 ± 0.21 | 0.00 |
| Poecilus punctulatus | 0.00 | 0.10 ± 0.10 | 0.20 ± 0.13 | 0.00 |
| Poecilus koyi | 0.20 ± 0.13 | 0.00 | 0.60 ± 0.40 | 0.00 |
| Poecilus versicolor | 1.70 ± 0.54 * | 6.80 ± 2.41 * | 3.10 ± 0.60 * | 0.11 ± 0.11 * |
| Pseudotaphoxenus tillesii | 0.00 | 0.10 ± 0.10 | 0.00 | 0.00 |
| Pterostichus macer | 0.00 | 0.10 ± 0.10 | 0.20 ± 0.13 | 0.00 |
| Individuals per one trap per week | 34.00 ± 4.49 | 32.70 ± 9.16 | 259.90 ± 31.14 ** | 3.00 ± 0.71 ** |
| Species per one trap | 4.7 ± 0.37 * | 3.00 ± 0.42 * | 7.1 ± 0.43 ** | 1.89 ± 0.42 ** |
| Number of species | 10 | 10 | 18 | 7 |
| Type of Soil Treatment | No-Till | Conventional |
|---|---|---|
| Spider Species | ||
| Robertus arundineti | 6.40 ± 2.32 * | 0.80 ± 0.80 * |
| Robertus sp. Juv. | 6.40 ± 3.76 | 0 |
| Clubiona caerulescens | 0.80 ± 0.80 | 0 |
| Drassyllus pusillus | 0.80 ± 0.80 | 0 |
| Gnaphosa sp. Juv. | 0.80 ± 0.80 | 0 |
| Haplodrassus signifer | 0.80 ± 0.80 | 0 |
| Tibellus maritimus | 0 | 0.80 ± 0.80 |
| Haplodrassus pseudosignifer | + | 0 |
| Number of individuals per square meter | 16.80 ± 3.44 ** | 1.60 ± 1.04 ** |
| Number of species per one soil sample | 0.20 ± 0.13 ** | 1.30 ± 0.15 ** |
| Type of Soil Treatment | No-Till | Conventional |
|---|---|---|
| Diversity Parameter | ||
| Simpson_1-D | 0.69 | 0.5 |
| Shannon_H | 1.44 | 0.69 |
| Evenness_e^H/S | 0.60 | 1 |
| Menhinick | 1.49 | 1.41 |
| Margalef | 1.94 | 1.44 |
| Equitability_J | 0.74 | 1 |
| Fisher_alpha | 3.54 | 0 |
| Berger–Parker | 0.41 | 0.5 |
| Year | 2017. May | 2018. June | ||
|---|---|---|---|---|
| Type of Soil Treatment | Conventional | No-Till | Conventional | No-Till |
| Species | ||||
| Alopecosa albostriata | 0 | 0 | 0.20 ± 0.13 | 0 |
| Alopecosa dimidiata | 0 | 0 | 0 | 0.10 ± 0.10 |
| Alopecosa cuneata | 0 | 0.40 ± 0.16 | 0 | 0.10 ± 0.10 |
| Alopecosa cursor | 0 | 0 | 0.10 ± 0.10 | 0 |
| Alopecosa farinosa | 0 | 0.10 ± 0.10 | 0 | 0 |
| Alopecosa schmidti | 0.11 ± 0.11 * | 0 * | 0 | 0 |
| Arctosa sp. Juv. | 0 | 0.10 ± 0.10 | 0 | 0 |
| Drassyllus pusillus | 0 | 0.60 ± 0.22 | 0.30 ± 0.15 | 0.40 ± 0.22 |
| Erigone atra | 0 | 0 | 0.10 ± 0.10 | 0 |
| Gnaphosa licenti | 0 | 0.10 ± 0.10 | 0 | 0.20 ± 0.13 |
| Gnaphosa sp. Juv. | 0 | 0.10 ± 0.10 | 0 | 0 |
| Haplodrassus pseudosignifer | 0 | 0.10 ± 0.10 | 0.60 ± 0.16 | 0.20 ± 0.13 |
| Haplodrassus signifer | 0 | 0.20 ± 0.13 | 0.30 ± 0.15 | 0 |
| Pardosa agrestis | 0 | 0.20 ± 0.13 | 0.50 ± 0.17 | 0.10 ± 0.10 |
| Pardosa bifasciata | 0 | 0.20 ± 0.13 | 0 * | 0.50 ± 0.22 * |
| Pardosa fulvipes | 0 | 0 | 0.10 ± 0.10 | 0 |
| Pardosa palustris | 0 | 0 | 0.20 ± 0.13 | 0.10 ± 0.10 |
| Pardosa plumipes | 0 | 0 | 1.70 ± 0.37 * | 0.10 ± 0.10 * |
| Pardosa sp. Juv. | 0 | 0 | 0 | 0.20 ± 0.13 |
| Robertus arundineti | 0 | 0 | 0.10 ± 0.10 | 0.10 ± 0.10 |
| Talavera aequipes | 0 | 0 | 0 | 0.10 ± 0.10 |
| Thanatus striatus | 0 | 0 | 0 | 0.10 ± 0.10 |
| Trochosa robusta | 0 | 0 | 0.10 ± 0.10 | 0 |
| Xerolycosa miniata | 0 | 0 | 0.50 ± 0.17 * | 0 * |
| Xysticus cristatus | 0 | 0.40 ± 0.16 | 0.10 ± 0.10 | 0 |
| Zelotes mundus | 0 | 0.20 ± 0.13 | 0 | 0 |
| Individuals per one trap per week | 0.11 ± 0.11 * | 3.11 ± 0.68 * | 4.90 ± 0.59 * | 2.20 ± 0.63 * |
| Species per one trap | 0.11 ± 0.11 * | 2.89 ± 0.61 * | 3.70 ± 0.37 * | 2.00 ± 0.29 * |
| Number of species | 1 | 11 | 14 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyubechanskii, I.I.; Golovanova, E.V.; Dudko, R.Y.; Azarkina, G.N.; Rusalimova, O.A.; Samoylova, E.S.; Shekhovtsov, S.V.; Barsukov, P.A. Impact of No-Tillage on Soil Invertebrate Communities in the Southern Forest Steppe of West Siberia: Preliminary Research. Diversity 2023, 15, 402. https://doi.org/10.3390/d15030402
Lyubechanskii II, Golovanova EV, Dudko RY, Azarkina GN, Rusalimova OA, Samoylova ES, Shekhovtsov SV, Barsukov PA. Impact of No-Tillage on Soil Invertebrate Communities in the Southern Forest Steppe of West Siberia: Preliminary Research. Diversity. 2023; 15(3):402. https://doi.org/10.3390/d15030402
Chicago/Turabian StyleLyubechanskii, Ilya I., Elena V. Golovanova, Roman Yu. Dudko, Galina N. Azarkina, Olga A. Rusalimova, Ekaterina S. Samoylova, Sergei V. Shekhovtsov, and Pavel A. Barsukov. 2023. "Impact of No-Tillage on Soil Invertebrate Communities in the Southern Forest Steppe of West Siberia: Preliminary Research" Diversity 15, no. 3: 402. https://doi.org/10.3390/d15030402