
Diversity and Community Structure of Typhlocybinae in the Typical Karst Rocky Ecosystem, Southwest China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sample Setup and Sampling
2.3. Sample Identification
2.4. Species Diversity Analysis
3. Results
3.1. Environmental Variables
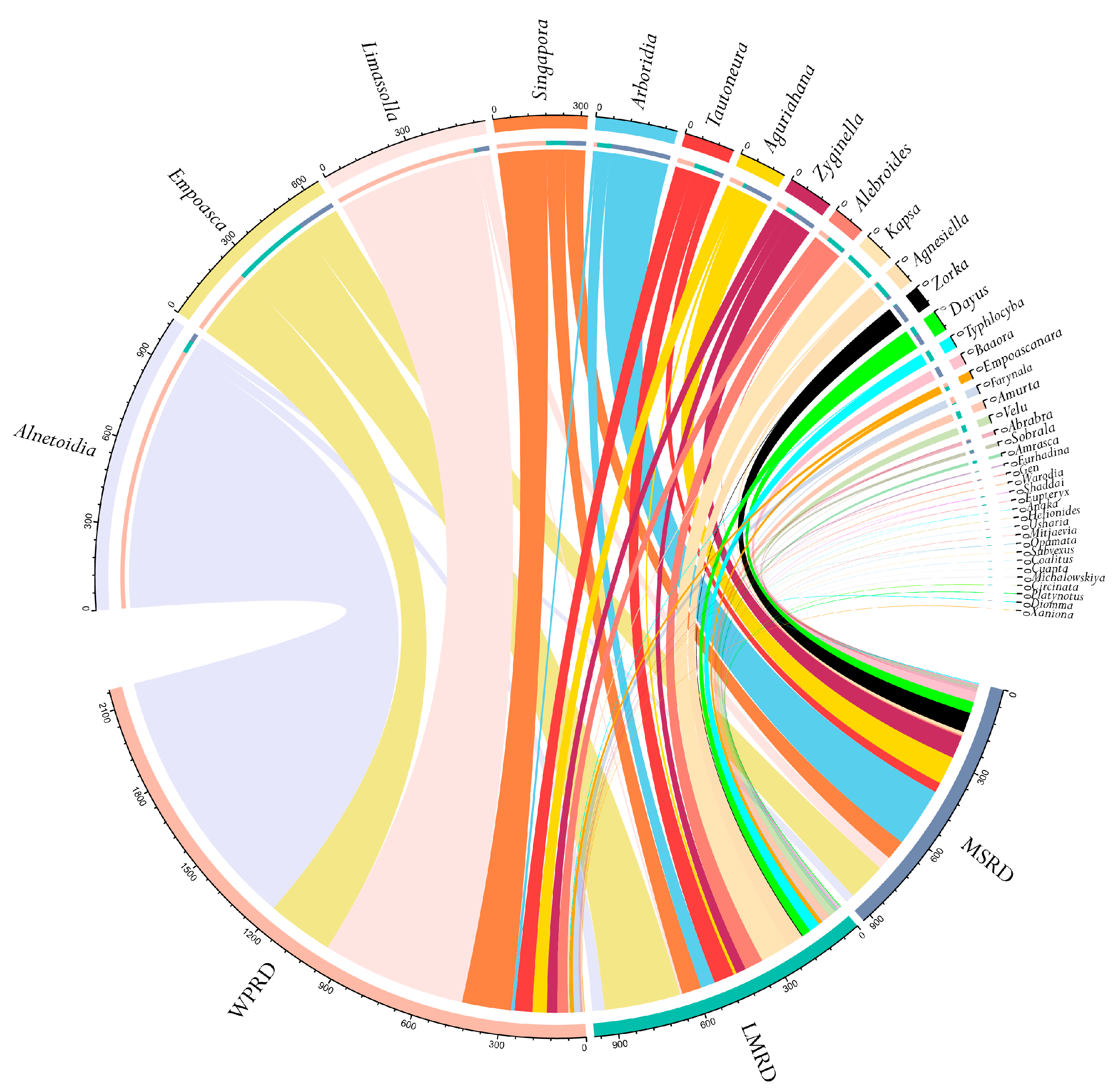
3.2. Typhlocybinae Community
3.3. Diversity Analysis
3.3.1. Community Diversity Analysis of Typhlocybinae in Different Study Areas
3.3.2. Community Diversity Analysis of Typhlocybinae in Different Host Plants
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.Z.; Qian, Y.Q. Frontiers in biodiversity science. Acta Ecol. Sin. 1997, 6, 3–10. [Google Scholar]
- Hector, A.; Bagchi, R. Biodiversity and ecosystem multifunctionality. Nature 2007, 448, 188–190. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Hillebrand, H.; Jonsson, P.R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology 2008, 89, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.X. The global perspective of karst rocky desertification and the countermeasures and experience of China’s governance. Pratacultural Sci. 2008, 25, 7. [Google Scholar]
- Gu, Y.; Chen, F.; Li, K.; Wu, H. Ecological control of land rocky desertification and vegetation recovery in karst region of Yunnan province. Sci. Technol. Rev. 2009, 27, 75–80. [Google Scholar]
- Yang, X.Y.; Zhou, Z.F.; Zou, C.H. Spatial pattern analysis of karst rocky desertification based on landscape level index. J. Guizhou Norm. Univ. (Nat. Sci.) 2010, 28, 23–27. [Google Scholar]
- Cao, J.H.; Yuan, D.X.; Tong, L.Q.; Malik, A.; Hui, Y.; Fen, H. An overview of karst ecosystem in southwest China: Current state and future management. J. Resour. Ecol. 2015, 6, 247–256. [Google Scholar]
- Wang, S.J. The Most Serious Ecogeologically environmental problem in southwestern China—Karst Rocky Desertification. Bull. Mineral. Petrol. Geochem. 2003, 2, 120–126. [Google Scholar]
- Wang, S.J. Concept deduction and its connotation of karst rocky desertification. Carsologica Sin. 2002, 21, 31–35. [Google Scholar]
- Sheng, M.Y.; Xiong, K.N.; Cui, G.Y.; Gao, Y. Plant diversity and soil physical-chemical properties in karst rocky desertification ecosystem of Guizhou, China. Acta Ecol. Sin. 2015, 35, 15. [Google Scholar]
- Zhou, Y.; Wei, Q.; Xiao, N.; Huang, J.; Gong, T.; Fei, Y.; Shi, Z.; Chen, H. Characteristics of soil Mites communities structure under vegetation vertical gradient in the Shibing World Natural Heritage Property, China. Forests 2022, 13, 598. [Google Scholar] [CrossRef]
- Cheng, C.; Gao, M.; Zhang, M.; Long, Y.D.; Wu, M.Z.; Li, Y.J.; Xiao, N. Effects of disturbance to moss biocrusts on soil nutrients, enzyme activities, and microbial communities in degraded karst landscapes in southwest China. Soil Biol. Biochem. 2021, 152, 108065. [Google Scholar] [CrossRef]
- Conrad, K.F.; Warren, M.S.; Fox, R. Rapid declines of common, widespread British moths provide evidence of an insect biodiversity crisis. Biol. Conserv. 2006, 132, 279–291. [Google Scholar] [CrossRef]
- Dietrich, C.H. Keys to the families of Cicadomorpha and subfamilies and tribes of Cicadellidae (Hemiptera: Auchenorrhyncha). Fla. Entomol. 2005, 88, 502–517. [Google Scholar] [CrossRef]
- Oman, P.W.; Knight, W.J.; Nielson, M.W. Leafhoppers (Cicadellidae): A bibliography, generic check-list and index to the world literature 1956–1985 | Annals of the entomological society of America | Oxford academic. Ann. Entomol. Soc. Am. 1990, 51, 53–89. [Google Scholar]
- Bartlett, C.R.; Deitz, L.L.; Dmitriev, D.A.; Sanborn, A.F.; Soulier-Perkins, A.; Wallace, M.S. The diversity of the true hoppers (Hemiptera: Auchenorrhyncha). Insect Biodivers. Sci. Soc. 2018, 2, 501–590. [Google Scholar]
- Yuan, S.; Huang, M.; Wang, X.S.; Ji, L.Q.; Zhang, Y.L. Centers of endemism and diversity patterns for typhlocybine leafhoppers (Hemiptera: Cicadellidae: Typhlocybinae) in China. Insect Sci. 2014, 21, 523–536. [Google Scholar] [CrossRef]
- Nickel, H.; Hildebrandt, J. Auchenorrhyncha communities as indicators of disturbance in grasslands (Insecta, Hemiptera)—A case study from the Elbe flood plains (northern Germany). Agric. Ecosyst. Environ. 2003, 98, 183–199. [Google Scholar] [CrossRef]
- Roddee, J.; Kobori, Y.; Hanboonsong, Y. Multiplication and distribution of sugarcane white leaf phytoplasma transmitted by the leafhopper, Matsumuratettix hiroglyphicus (Matsumura) (Hemiptera: Cicadellidae), in infected sugarcane. Sugar Tech. 2018, 20, 445–453. [Google Scholar] [CrossRef]
- Yang, X.; Chen, X.X.; Yuan, Z.W.; Su, D.; Song, Y.H. Community structure and dynamics of leafhopper in different land restoration types in the light-moderate rocky desertification areas. Acta Ecol. Sin. 2022, 42, 6790–6800. [Google Scholar]
- Gu, W.; Ma, L.; Sun, H.; Wang, L.; Zhang, Z. Variation in structure and dynamics of insect community in larch plantations under different soil conditions. Sci. Silvae Sin. Acta Ecol. Sin. 2017, 53, 97–106. [Google Scholar]
- Chen, X.X.; Jiang, J.; Zhang, N.; Yang, X.; Chi, Y.K.; Song, Y.H. Effects of habitat fragmentation on the population structure and genetic diversity of Erythroneurini in the typical karst rocky ecosystem, Southwest China. Insects 2022, 13, 499. [Google Scholar] [CrossRef] [PubMed]
- Xiong, K.N.; Li, J.; Long, M.Z. Features of soil and water loss and key issues in demonstration areas for combating karst rocky desertification. Acta Geogr. Sin. 2012, 67, 878–888. [Google Scholar]
- Cui, L.; Xiong, K.N.; Guan, Z.H.; Chen, Y.B.; Liu, Z.Q. The model of karst land management based on plant diversity restoration and protection: Examples from Salaxi and Huajiang areas of Guizhou. Carsologica Sin. 2016, 35, 513–524. [Google Scholar]
- Gu, Z.F.; Liu, Q.; Lu, Y.R. Relationships between the rock and soil chemical element contents and rocky desertification difference in karst rocky desertification process: A case study on the Zhenfeng-Guanling Huajiang area of Guizhou Province. Carsologica Sin. 2016, 35, 533–538. [Google Scholar]
- Yin, L.J.; Zhou, Z.F.; Zhu, C.L.; Shang, M.J. Geomorphological differentiation characteristics of land use change in karst rocky desertification area-A case of Guanling-Zhenfeng Huajiang rocky desertification comprehensive demonstration zone in Guizhou Province. Bull. Soil Water Conserv. 2020, 40, 37–46. [Google Scholar]
- Liu, P.P.; Xiao, H.; Chen, H. Temporal and spatial evolution characteristics of rocky desertification control landscape pattern in Salaxi demonstration area in Bijie city. Southwest China J. Agric. Sci. 2020, 33, 2316–2324. [Google Scholar]
- Zhang, Y.Y.; Chen, Q.W. On ecological benefit monitoring of rocky desertification control project in typical demonstration areas: A case study of Salaxi in Bijie and Huajiang in Guanling. J. Leshan Norm. Univ. 2018, 33, 58–67. [Google Scholar]
- Li, Y.; Liu, Z.Q.; Liu, X.X.; Cao, Y.; Li, K.P. Effect of overlying soils and bedrock on drip water elements in rocky desertification area in Bijie, Guizhou. Environ. Chem. 2016, 35, 1894–1902. [Google Scholar]
- Cai, Y.; Luo, X.J.; Wang, X.S.; Wang, Y.Q.; Wang, S.; Liu, Y.L. Evolution characteristics of plant communities in the Karst area of Dolomites, Southeastern Guizou province. Northwest J. Bot. 2019, 39, 2064–2071. [Google Scholar]
- Wu, X.Z. Floristic analysis of pteridophyte in karst forest area of Guizhou Shibing Yuntaishan Nature Reserve. J. Anhui Agric. Sci. 2012, 40, 869–870. [Google Scholar]
- Yang, S.L.; Luo, Q.H. New records and species richness of the family cerambycidae in Yuntaishan Area of Shibing, Guizhou. Guizhou Agric. Sci. 2013, 41, 103–106. [Google Scholar]
- Zhang, Z.; Huang, X.; Zhou, Y. Factors influencing the evolution of human-driven rocky desertification in karst areas. Land Degrad. Dev. 2021, 32, 817–829. [Google Scholar] [CrossRef]
- Dmitriev, D.A. 3i Typhlocybinae: 3i Interactive Keys and Taxonomic Databases, Subfamily Typhlocybinae-Oct 2011 Version. Species 2000 & ITIS Catalogue of Life. Species 2000; Reading: Berkshire, UK, 2014. [Google Scholar]
- Li, Z.Z.; Li, H.; Xing, J.C. Insects (Auchenorrhyncha) Type Specimens in the Collection of Guizhou University; Guizhou Science and Technology Press: Guiyang, China, 2014. [Google Scholar]
- Meng, J. Systematic Study on the Tribe Alebrini and Dikraneurini from China (Hemiptera: Cicadellidae; Typhlocybinae); The University of Guizhou Press: Guiyang, China, 2017. [Google Scholar]
- Song, Y.H.; Li, Z.Z. Erythroneurini and Zyginellini from China (Hemiptera: Cicadellidae: Typhlocybinae); Science and Technology Publishing House: Guiyang, China, 2014; pp. 12–209. [Google Scholar]
- Luo, F.F.; Zhou, R.; Su, W.H. Effects of drought on the water content of typical plants in the central Yunnan plateau. J. Anhui Agric. Sci. 2012, 40, 12745–12747. [Google Scholar]
- Hong, X.M.; Xin, Y.; Li, J.L. Butterfly diversity and its influencing factors in Saihanwula Nature Reserve. Biodivers. Sci. 2018, 6, 590–600. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Urbana, IL, USA, 1947; pp. 10–98. [Google Scholar]
- Margalef, D.R. Information theory in ecology. Gen. Syst. 1957, 3, 36–71. [Google Scholar]
- Pielou, E.C. Shannon’s formula as a measure of specific diversity: Its use and misuse. Am. Nat. 1966, 100, 463–465. [Google Scholar] [CrossRef]
- Peet, R.K. The measurement of species of diversity. Annu. Rev. Ecol. Evol. Syst. 2014, 5, 285. [Google Scholar] [CrossRef]
- Baho, D.L.; Futter, M.N.; Johnson, R.K.; Angeler, D.G. Assessing temporal scales and patterns in time series: Comparing methods based on redundancy analysis. Ecol. Complex. 2015, 22, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, K.N.; Gokhale, L.A. Advanced Charting Techniques of Microsoft Excel 2016 Aiming Visualization. Int. J. Comput. Sci. Eng. 2019, 7, 198–207. [Google Scholar] [CrossRef]
- Cronk, B. How to Use IBM SPSS Statistics: A Step-By-Step Guide to Analysis and Interpretation; Routledge: London, UK, 2016. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Zhang, M.; Zhao, Y.; Zhang, L.; Wu, J.; Li, S.; Zhu, G.; Sun, L. Comparison of Characteristics of Microbial Community Structure in Sewage Treatment Plants of High Altitude Area and Low Altitude Area. Environ. Eng. Sci. 2022, 40, 66–73. [Google Scholar]
- El-Hady, R.M.; El-Hawagry, M.S.A.; Soliman, M.M. Diversity and temporal variations of the leafhopper fauna (Cicadellidae, Auchenorrhyncha, Hemiptera) in two ecological zones of Egypt. J. Nat. Hist. 2020, 54, 2869–2887. [Google Scholar] [CrossRef]
- Hollier, J.A.; Maczey, N.; Masters, G.J.; Mortimer, S.R. Grassland leafhoppers (Hemiptera: Auchenorrhyncha) as indicators of habitat condition–A comparison of between-site and between-year differences in assemblage composition. J. Insect Conserv. 2005, 9, 299–307. [Google Scholar] [CrossRef]
- Jin, Q.; Ruan, Q.; Chen, F.; Liu, B.; Cai, W.J.; Zhang, A.B. Species diversity and phylogenetic diversity of moth species on the Qinghai-Tibet Plateau. J. Environ. Entomol. 2018, 40, 102–110. [Google Scholar]
- Yuan, C.J.; Xiong, K.N.; Rong, L.; Weng, Y.F. Research progress on the biodiversity during the ecological restoration of larst rocky desertification. Earth Environ. 2021, 49, 336–345. [Google Scholar]
- Diekötter, T.; Billeter, R.; Crist, T.O. Effects of landscape connectivity on the spatial distribution of insect diversity in agricultural mosaic landscapes. Basic Appl. Ecol. 2008, 9, 298–307. [Google Scholar] [CrossRef]
- Edwards, D.P.; Tobias, J.A.; Sheil, D.; Meijaard, E.; Laurance, W.F. Maintaining ecosystem function and services in logged tropical forests. Trends Ecol. Evol. 2014, 29, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.S. Diversity, disturbance, and sustainable use of Neotropical forests: Insects as indicators for conservation monitoring. J. Insect Conserv. 1997, 1, 25–42. [Google Scholar] [CrossRef]
- Schmitz, O.J.; Rosenblatt, A.E.; Smylie, M. Temperature dependence of predation stress and the nutritional ecology of a generalist herbivore. Ecology 2016, 97, 3119–3130. [Google Scholar] [CrossRef]
- Geppert, C.; La-Bella, G.; Boscutti, F.; Sanna, F.; Lorenzo, M. Effects of temperature and plant diversity on orthopterans and leafhoppers in calcareous dry grasslands. J. Insect Conserv. 2021, 25, 287–296. [Google Scholar] [CrossRef]
- Iqbal, J.; Ashfaq, M.; Hasan, M.; Sagheer, M.; Nadeem, M. Influence of abiotic factors on population fluctuation of leaf hopper, Amrasca biguttula biguttula (Ishida) on Okra. Pak. J. Zool. 2010, 42, 5. [Google Scholar]
- Helbing, F.; Fartmann, T.; Löffler, F.; Poniatowski, D. Effects of local climate, landscape structure and habitat quality on leafhopper assemblages of acidic grasslands. Agric. Ecosyst. Environ. 2017, 246, 94–101. [Google Scholar] [CrossRef]
- Lawton, J.H. Plant architecture and the diversity of phytophagous insects. Annu. Rev. Entomol. 1983, 28, 23–39. [Google Scholar] [CrossRef]
- Zhang, K.; Lin, S.L.; Ji, Y.Q.; Yang, C.X.; Wang, X.Y. Plant diversity accurately predicts insect diversity in two tropical landscapes. Mol. Ecol. 2016, 25, 4407–4419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauker, F.; Diekotter, T.; Schwarzbach, F.; Wolters, V. Pollinator dispersal in an agricultural matrix: Opposing responses of wild bees and hoverflies to landscape structure and distance from main habitat. Landsc. Ecol. 2009, 24, 547–555. [Google Scholar] [CrossRef]
- Taft, J.B.; Dietrich, C.H. A test of concordance in community structure between leafhoppers and grasslands in the central Tien Shan Mountains. Arthropod Plant Interact 2017, 11, 843–859. [Google Scholar] [CrossRef]
- Rösch, V.; Tscharntke, T.; Scherber, C.; Batáry, P. Landscape composition, connectivity and fragment size drive effects of grassland fragmentation on insect communities. J. Appl. Ecol. 2013, 50, 387–394. [Google Scholar] [CrossRef]
- Scherber, C.; Heimann, J.; Köhler, G.; Mitschunas, N.; Weisser, W.W. Functional identity versus species richness: Herbivory resistance in plant communities. Oecologia 2010, 163, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, E.; Murillo, V.; Orwat, J. Leafhopper food plants in a Neotropical forest in Panama (Hemiptera: Cicadellidae). An. Da Acad. Bras. Ciências 2021, 93, e20190171. [Google Scholar] [CrossRef]
- Branquart, E.; Hemptinne, J.L. Selectivity in the exploitation of floral resources by hoverflies (Diptera: Syrphinae). Ecography 2000, 23, 732–742. [Google Scholar] [CrossRef]
- Jovii, S.; Burgio, G.; Diti, I.; Duanka, K.; Ante, V. Influence of landscape structure and land use on Merodon and Cheilosia (Diptera: Syrphidae): Contrasting responses of two genera. J. Insect Conserv. 2017, 21, 53–64. [Google Scholar]
- Shen, M.W.; Chen, S.B.; Bi, M.J.; Chen, W.D.; Zhou, K.X. Relationship between geographic patterns of ant species richness and environmental factors in China. J. Ecol. 2016, 36, 7732–7739. [Google Scholar]
- Harison, S.; Cornell, H. Toward a better understanding of the regional causes of local community richness. Ecol. Lett. 2008, 11, 969–979. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, G. Distribution pattern and mechanism of insect species diversity in Inner Mongolia. Biodivers. Sci. 2018, 26, 701–706. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Areas | Host Plant | Altitude (m) | Latitude | Longitude |
|---|---|---|---|---|
| Juglans regia | 1200 | 25°42′29″ N | 105°36′50″ E | |
| Rhus chinensis | 1280 | 25°41′36″ N | 105°37′46″ E | |
| Huajiang | Prunus persica | 1193 | 25°40′43″ N | 105°38′57″ E |
| Prunus salicina | 1190 | 25°37′43″ N | 105°41′8″ E | |
| Debregeasia orientalis | 1240 | 25°39′56″ N | 105°38′1″ E | |
| Juglans regia | 1960 | 27°14′5″ N | 105°5′51″ E | |
| Rhus chinensis | 1760 | 27°14′43″ N | 105°6′16″ E | |
| Bijie | Prunus persica | 1959 | 27°15′49″ N | 105°5′13″ E |
| Prunus salicina | 1766 | 27°14′51″ N | 105°5′52″ E | |
| Debregeasia orientalis | 1700 | 27°13′25″ N | 105°5′47″ E | |
| Juglans regia | 950 | 27°2′39″ N | 108°7′23″ E | |
| Rhus chinensis | 952 | 27°2′53″ N | 108°7′25″ E | |
| Shibing | Prunus persica | 1000 | 27°12′48″ N | 108°11′5″ E |
| Prunus salicina | 1140 | 27°13′35″ N | 108°1′54″ E | |
| Debregeasia orientalis | 912 | 27°7′51″ N | 108°2′27″ E |
| Month | Host Plants | Huajiang | Bijie | Shibing | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Humidity (%) | Precipitation Days | Temperature (°C) | Humidity (%) | Precipitation Days | Temperature (°C) | Humidity (%) | Precipitation Days | ||
| September | JR | 22.70 | 74.68 | 22 | 19.79 | 80.57 | 24 | 24.16 | 78.46 | 22 |
| RC | 24.94 | 69.47 | 22 | 19.14 | 82.00 | 24 | 24.03 | 77.57 | 22 | |
| PP | 23.05 | 72.58 | 22 | 19.78 | 80.26 | 24 | 23.4 | 77.15 | 22 | |
| PS | 23.00 | 72.59 | 22 | 19.69 | 80.97 | 24 | 23.74 | 77.10 | 22 | |
| DO | 24.25 | 70.86 | 22 | 19.48 | 81.46 | 24 | 23.36 | 77.05 | 22 | |
| May | JR | 21.48 | 70.97 | 13 | 16.71 | 79.62 | 24 | 19.96 | 83.48 | 23 |
| RC | 23.21 | 69.46 | 13 | 16.23 | 78.48 | 24 | 19.25 | 82.66 | 23 | |
| PP | 22.55 | 69.88 | 13 | 16.01 | 78.37 | 24 | 19.00 | 83.30 | 23 | |
| PS | 22.51 | 69.72 | 13 | 16.77 | 78.24 | 24 | 19.21 | 82.42 | 23 | |
| DO | 22.74 | 69.11 | 13 | 16.50 | 77.73 | 24 | 19.24 | 82.45 | 23 | |
| July | JR | 25.29 | 80.78 | 19 | 22.82 | 81.37 | 12 | 27.92 | 78.68 | 10 |
| RC | 27.21 | 77.52 | 19 | 20.12 | 79.48 | 12 | 27.12 | 78.02 | 10 | |
| PP | 24.11 | 78.63 | 19 | 20.11 | 80.25 | 12 | 26.12 | 77.67 | 10 | |
| PS | 24.14 | 78.35 | 19 | 21.67 | 79.24 | 12 | 26.37 | 77.24 | 10 | |
| DO | 24.51 | 77.56 | 19 | 20.45 | 79.14 | 12 | 27.00 | 77.38 | 10 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Jiang, J.; Chi, Y.; Su, D.; Song, Y. Diversity and Community Structure of Typhlocybinae in the Typical Karst Rocky Ecosystem, Southwest China. Diversity 2023, 15, 387. https://doi.org/10.3390/d15030387
Wang J, Jiang J, Chi Y, Su D, Song Y. Diversity and Community Structure of Typhlocybinae in the Typical Karst Rocky Ecosystem, Southwest China. Diversity. 2023; 15(3):387. https://doi.org/10.3390/d15030387
Chicago/Turabian StyleWang, Jinqiu, Jia Jiang, Yongkuan Chi, Di Su, and Yuehua Song. 2023. "Diversity and Community Structure of Typhlocybinae in the Typical Karst Rocky Ecosystem, Southwest China" Diversity 15, no. 3: 387. https://doi.org/10.3390/d15030387