
A Synoptic Review of the Cartilaginous Fishes (Chondrichthyes: Holocephali, Elasmobranchii) from the Upper Jurassic Konservat-Lagerstätten of Southern Germany: Taxonomy, Diversity, and Faunal Relationships
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Systematic and Taxonomic Considerations
2.2. Methodological Approaches
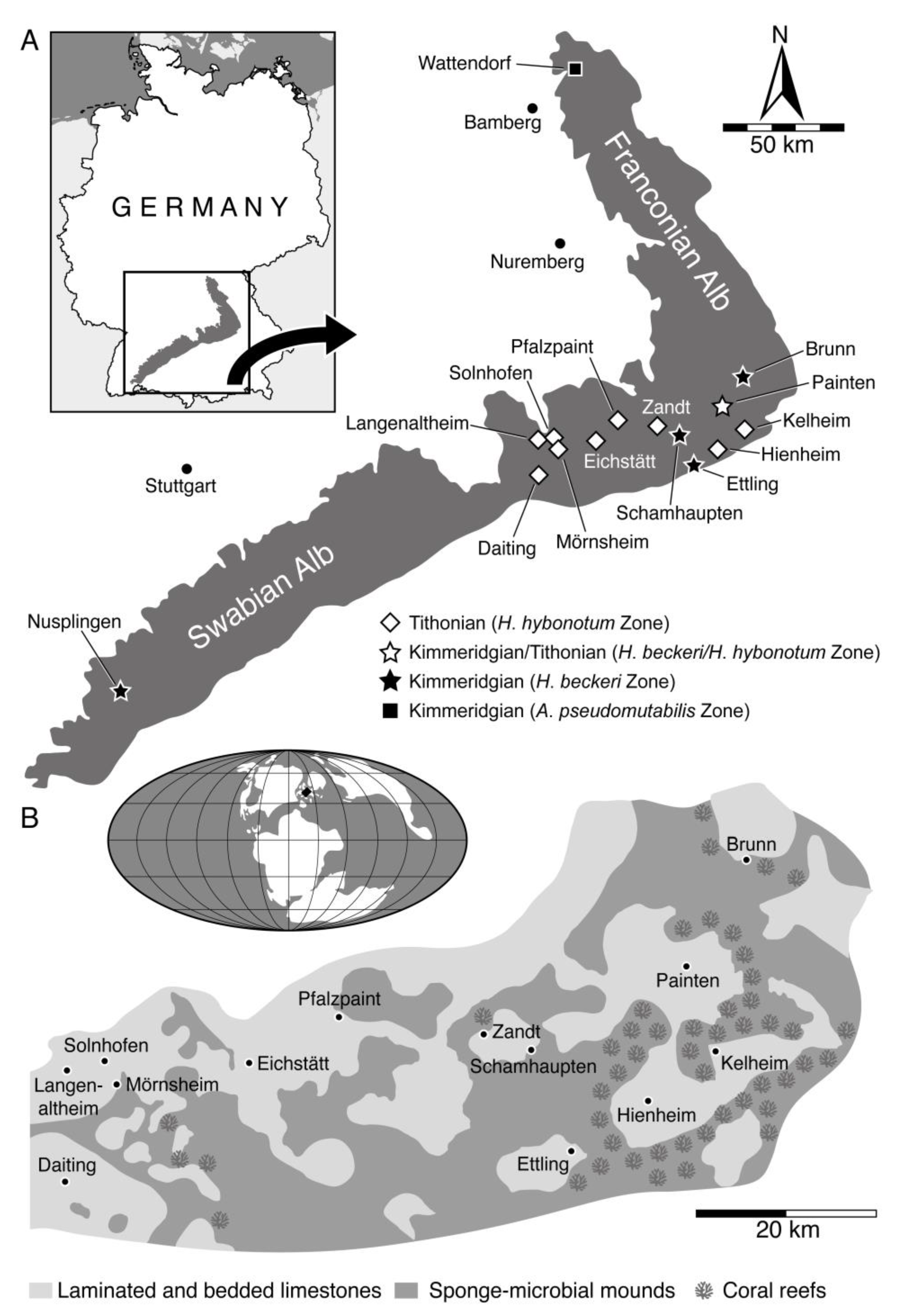
3. Geographical and Geological Setting
3.1. Wattendorf
3.2. Solnhofen Archipelago
3.3. Nusplingen

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
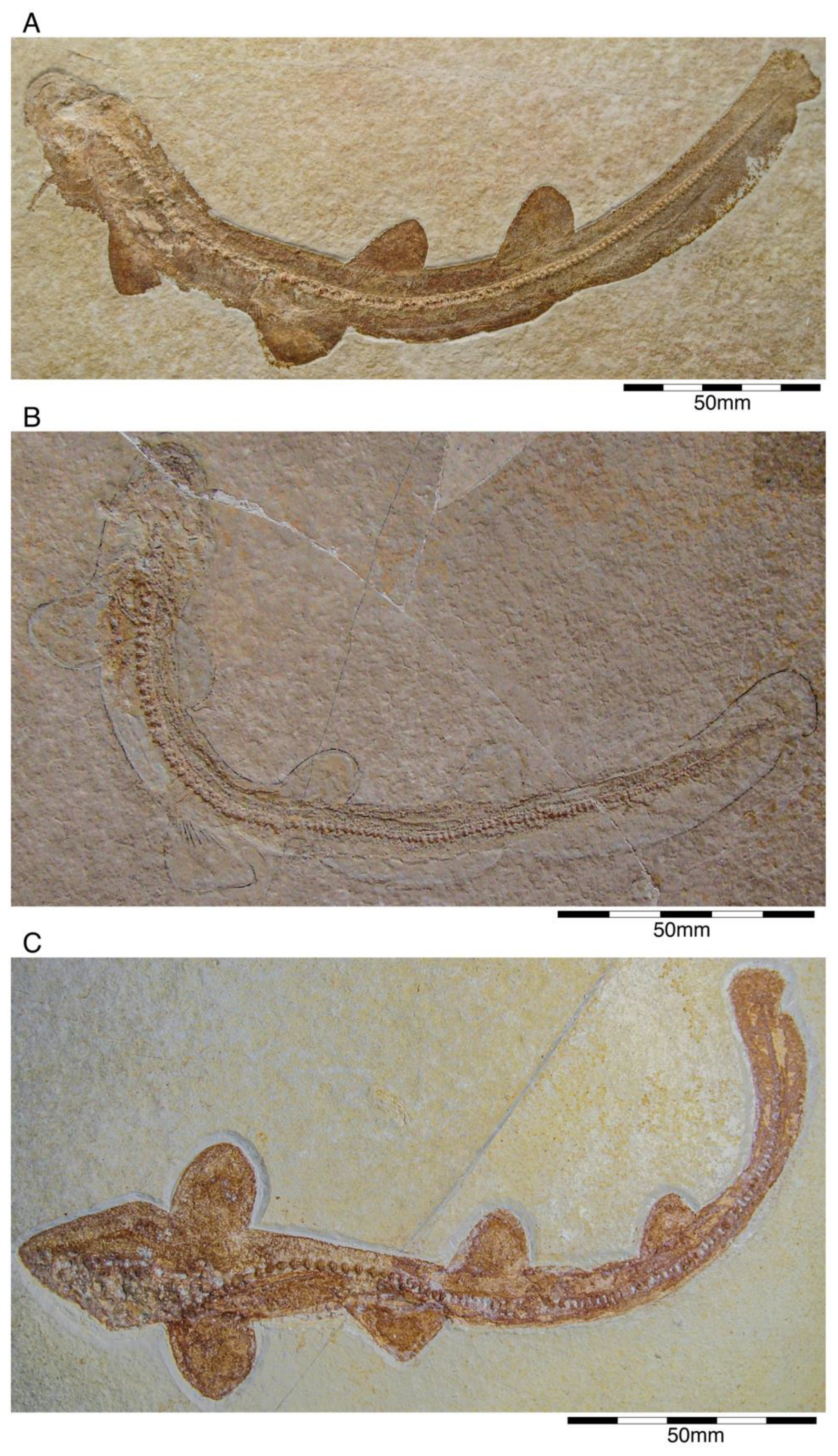{kind=link}
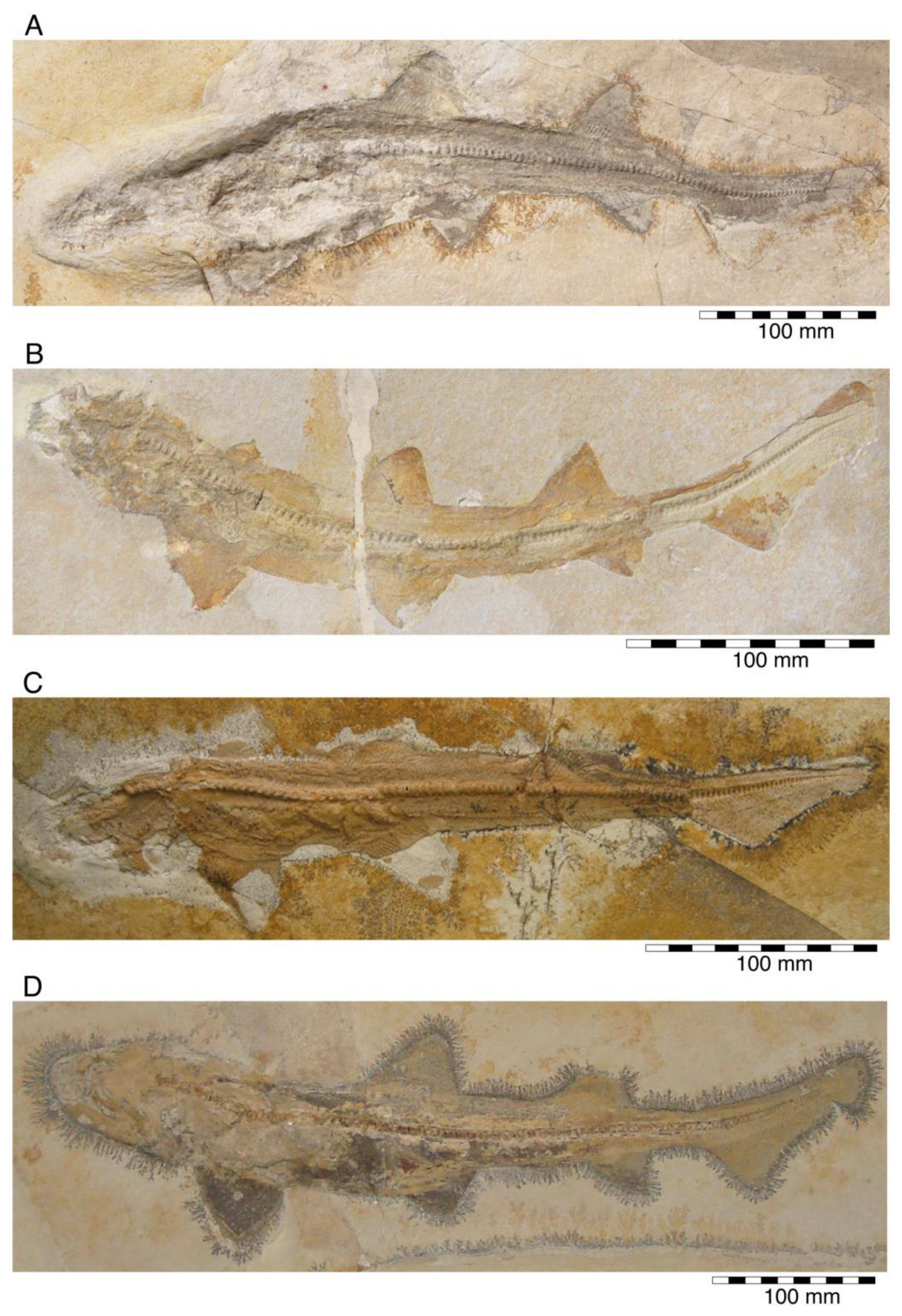{kind=link}
{kind=link}
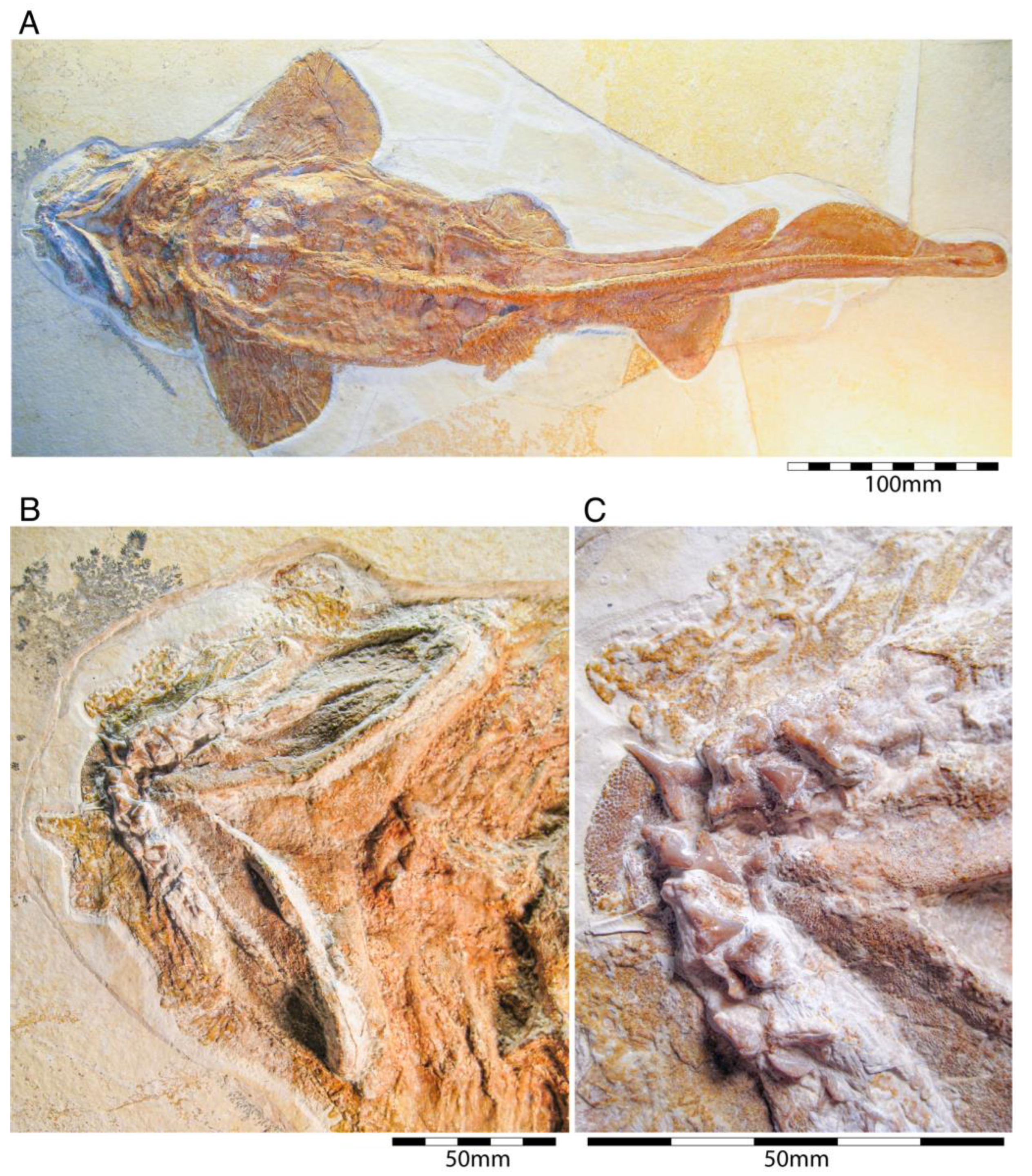{kind=link}
{kind=link}
{kind=link}
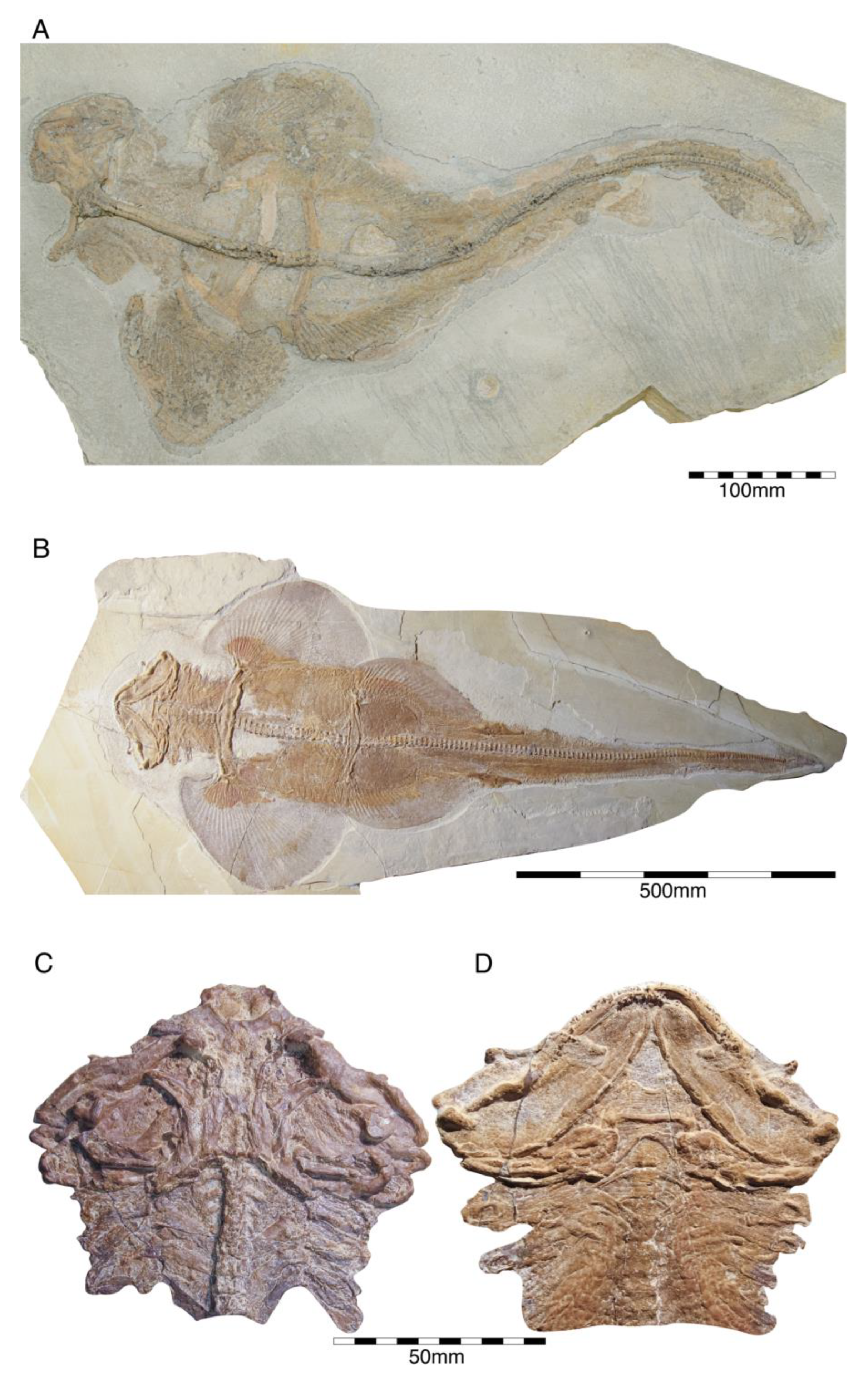{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4. Taxonomic Review
4.1. Holocephali
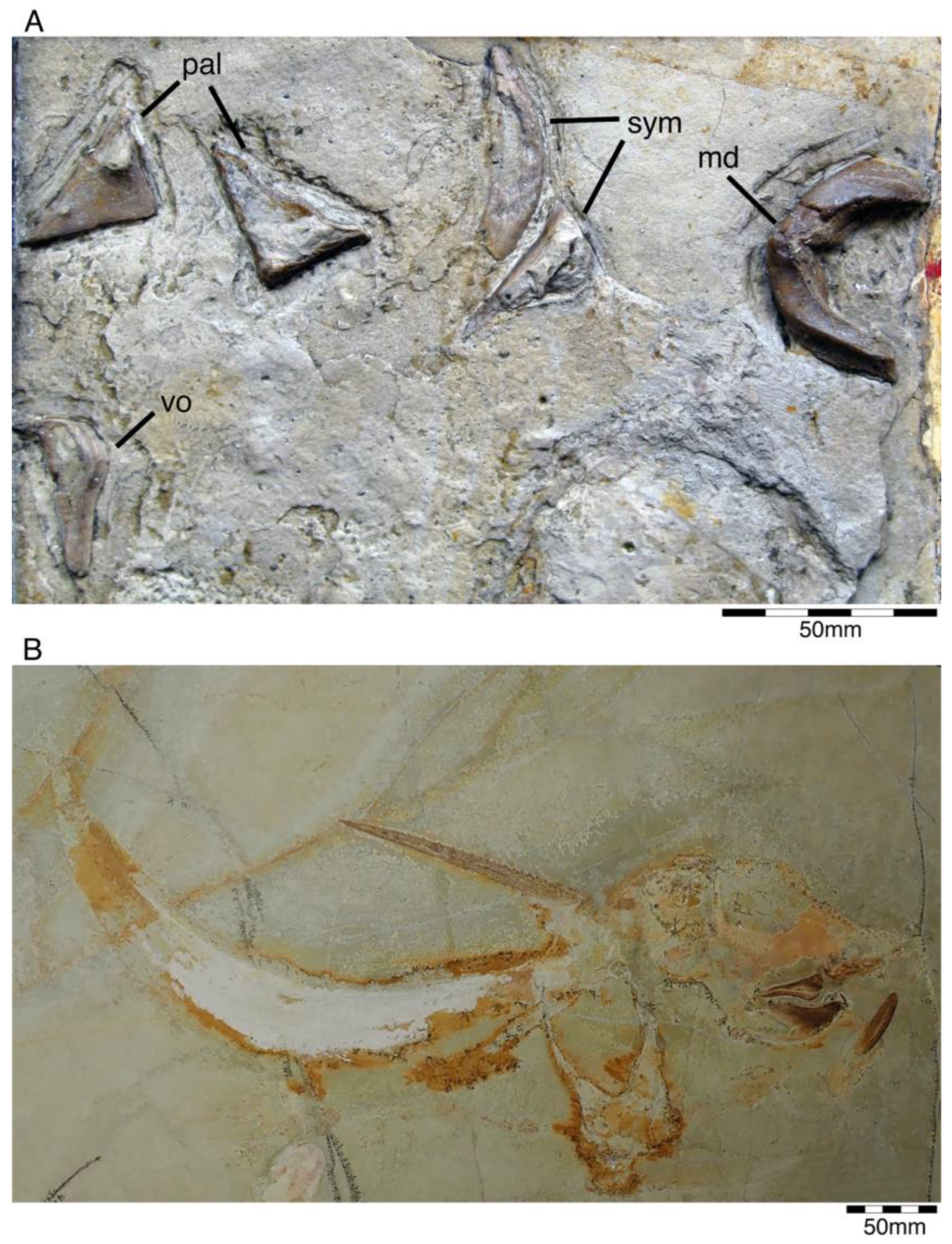
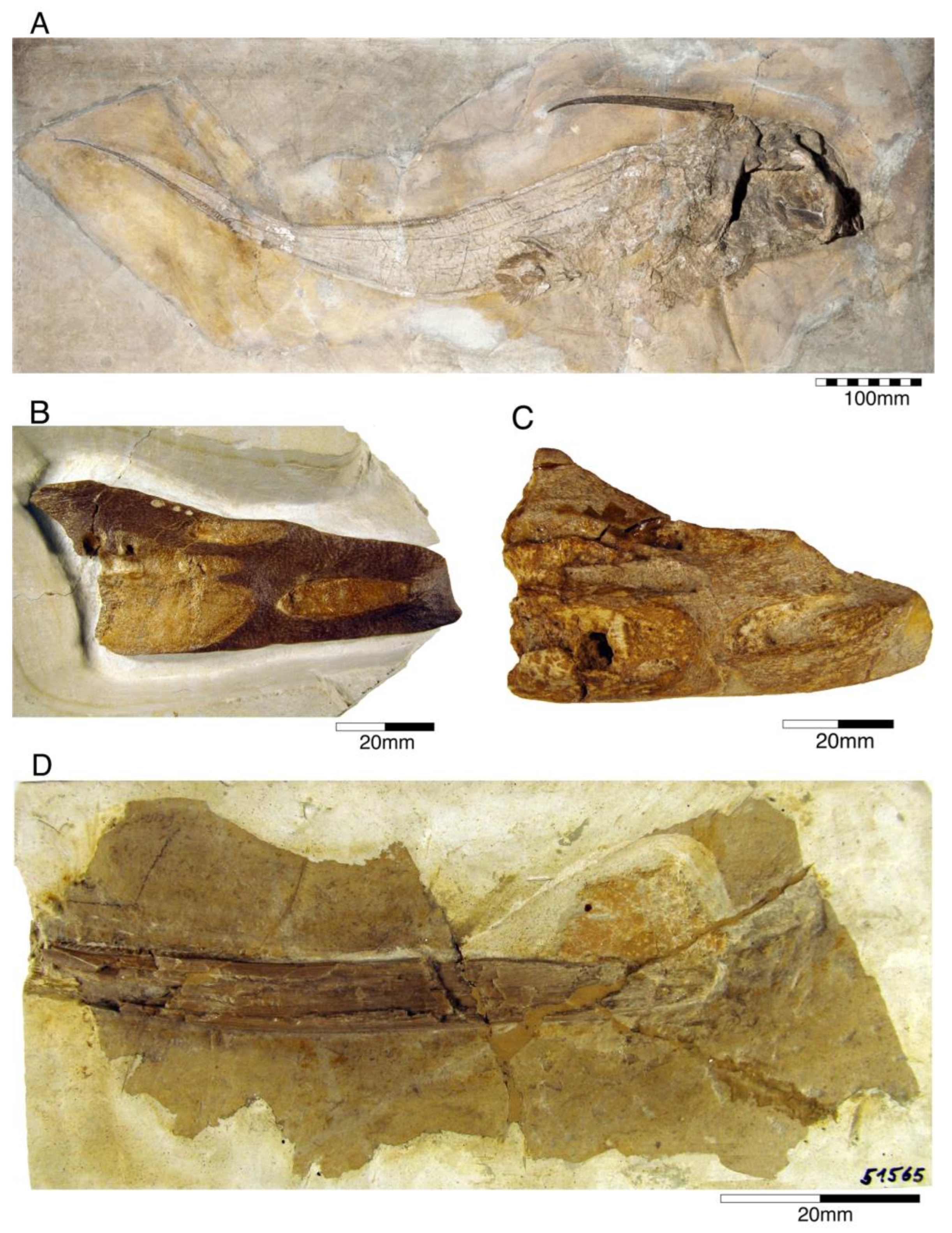
4.1.1. Chimaeriformes
- Class Chondrichthyes Huxley, 1880 [84]
- Subclass Subterbranchialia Zangerl, 1979 [85]
- Superorder Holocephali Bonaparte, 1832 [86]
- Order Chimaeriformes Obruchev, 1953 [87]
- Suborder Myriacanthoidei Patterson, 1965 [88]
- Family Chimaeropsidae Patterson, 1965 [88]
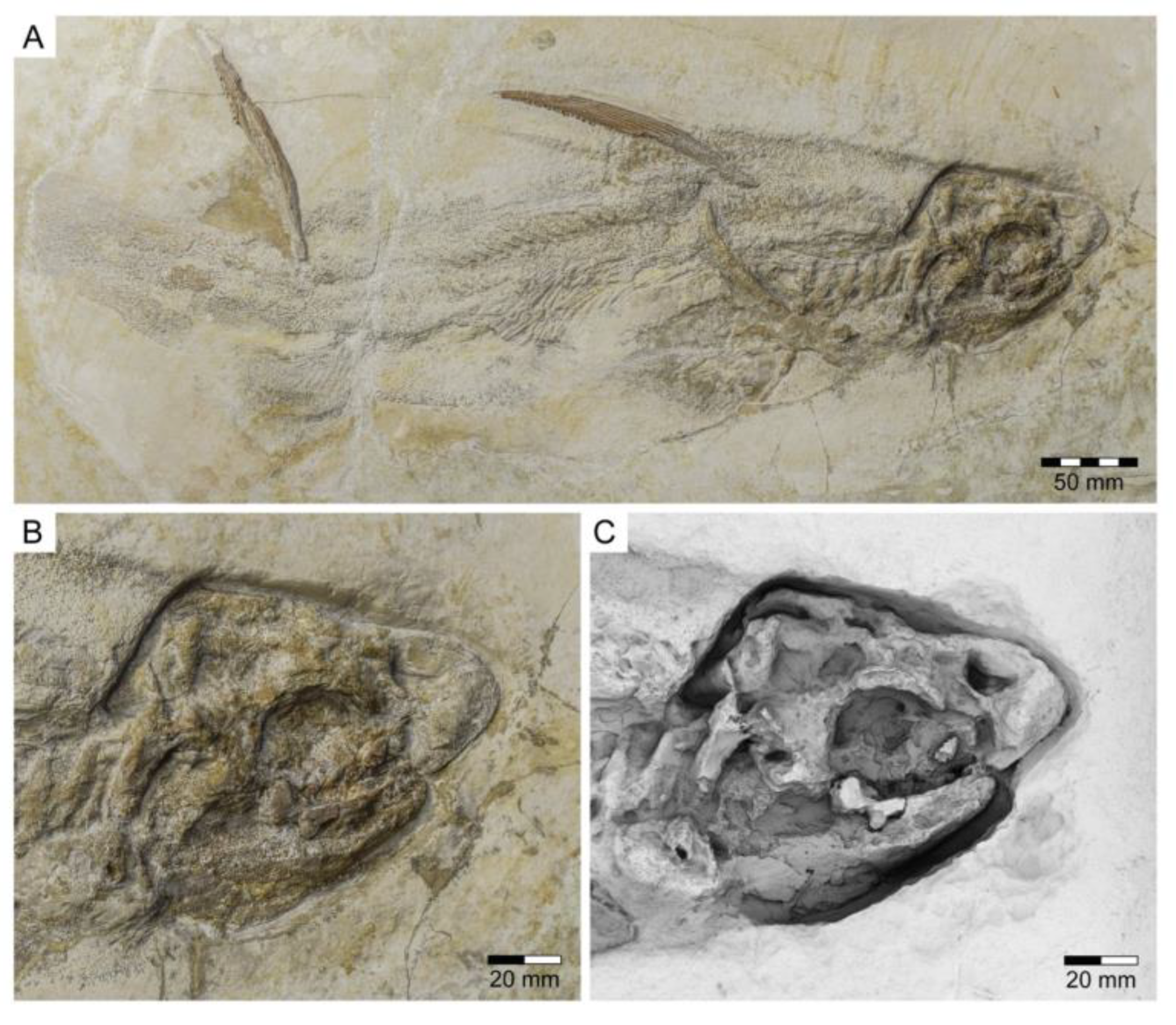
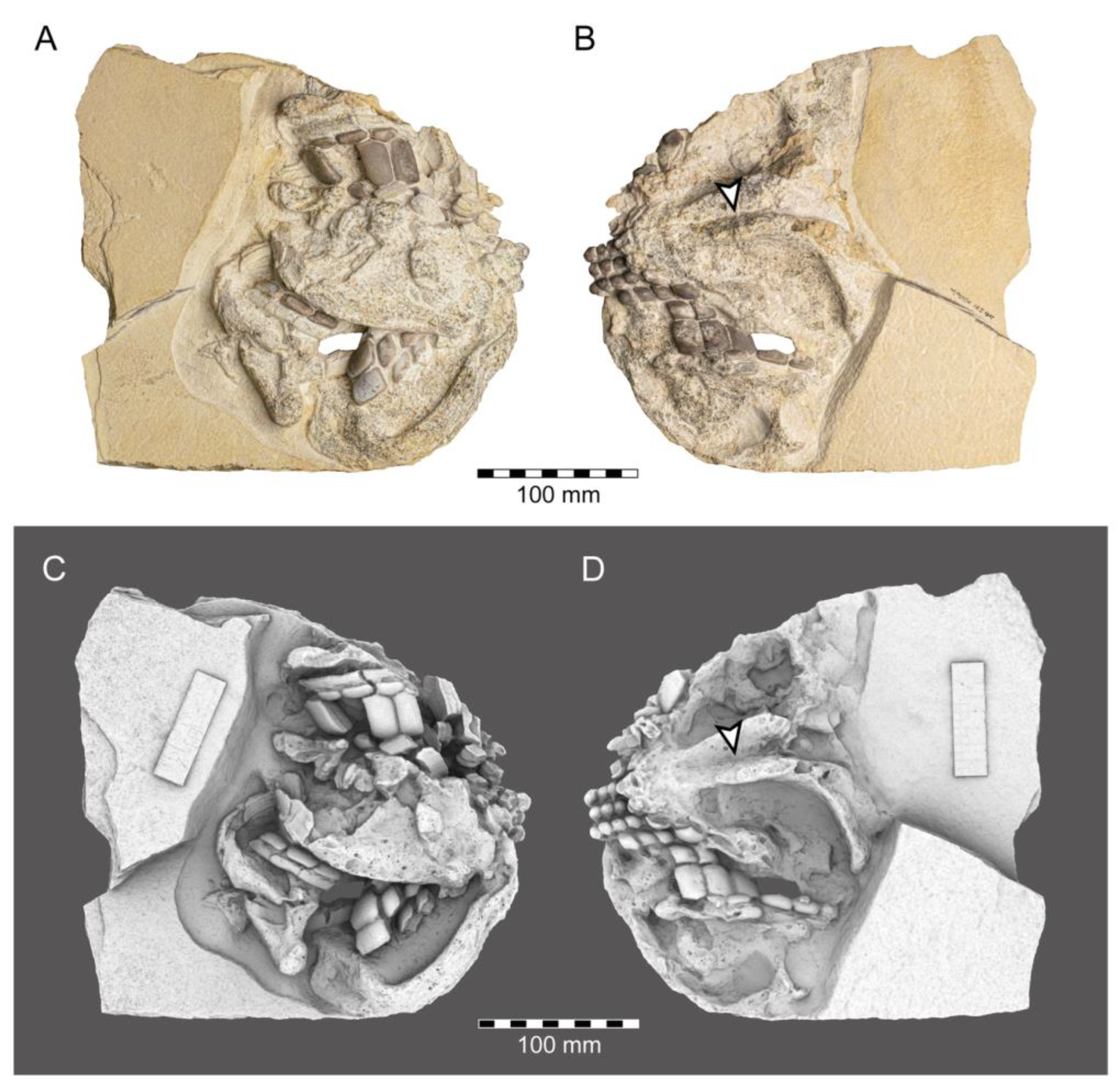
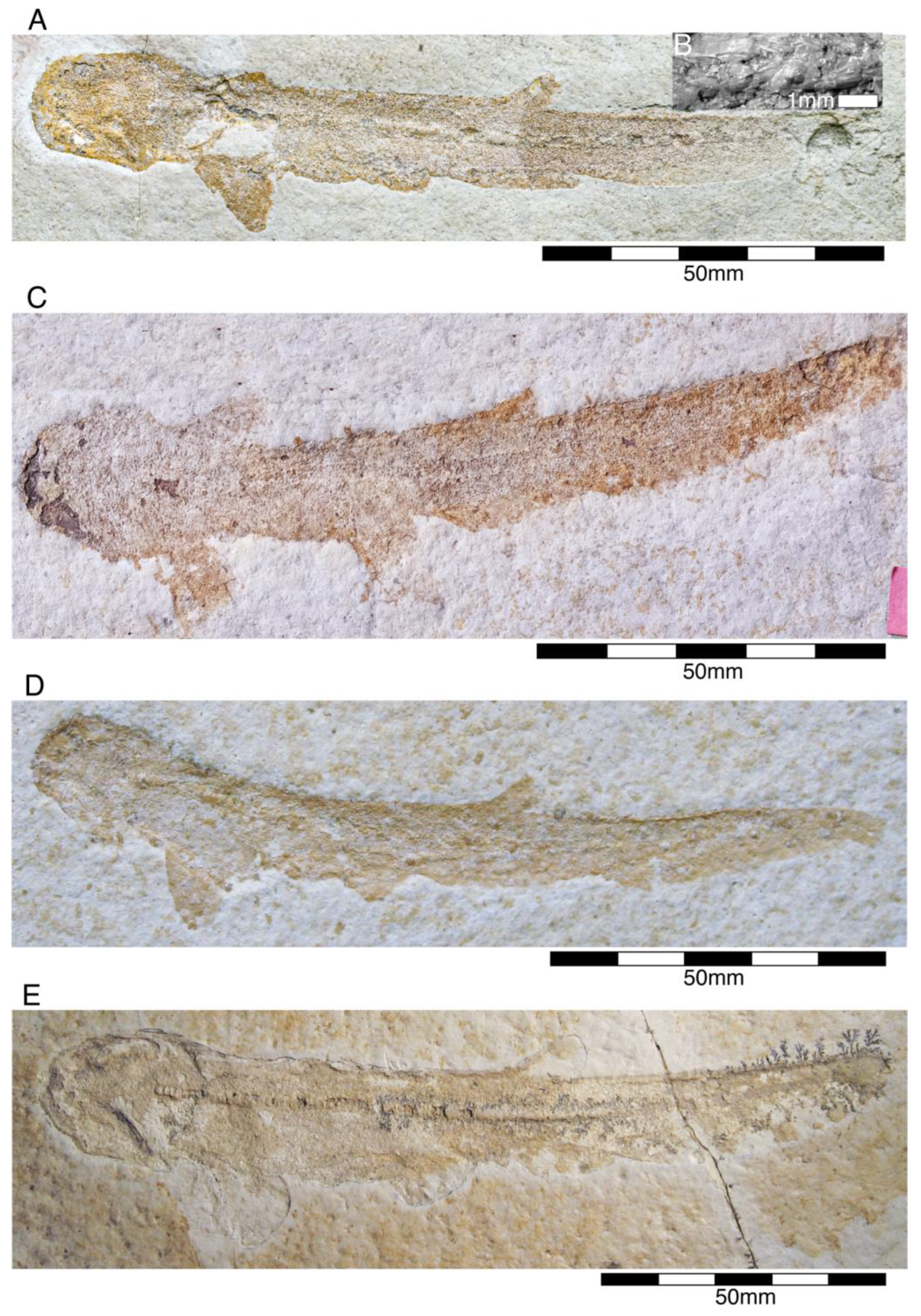
- Genus Chimaeropsis Zittel, 1887 [89]


- Family Rhinochimaeridae Garman, 1901 [99]
- Genus †Elasmodectes Newton, 1878 [124]
- †Elasmodectes avitus (von Meyer, 1862 [120])


Egg Capsules of Holocephalians
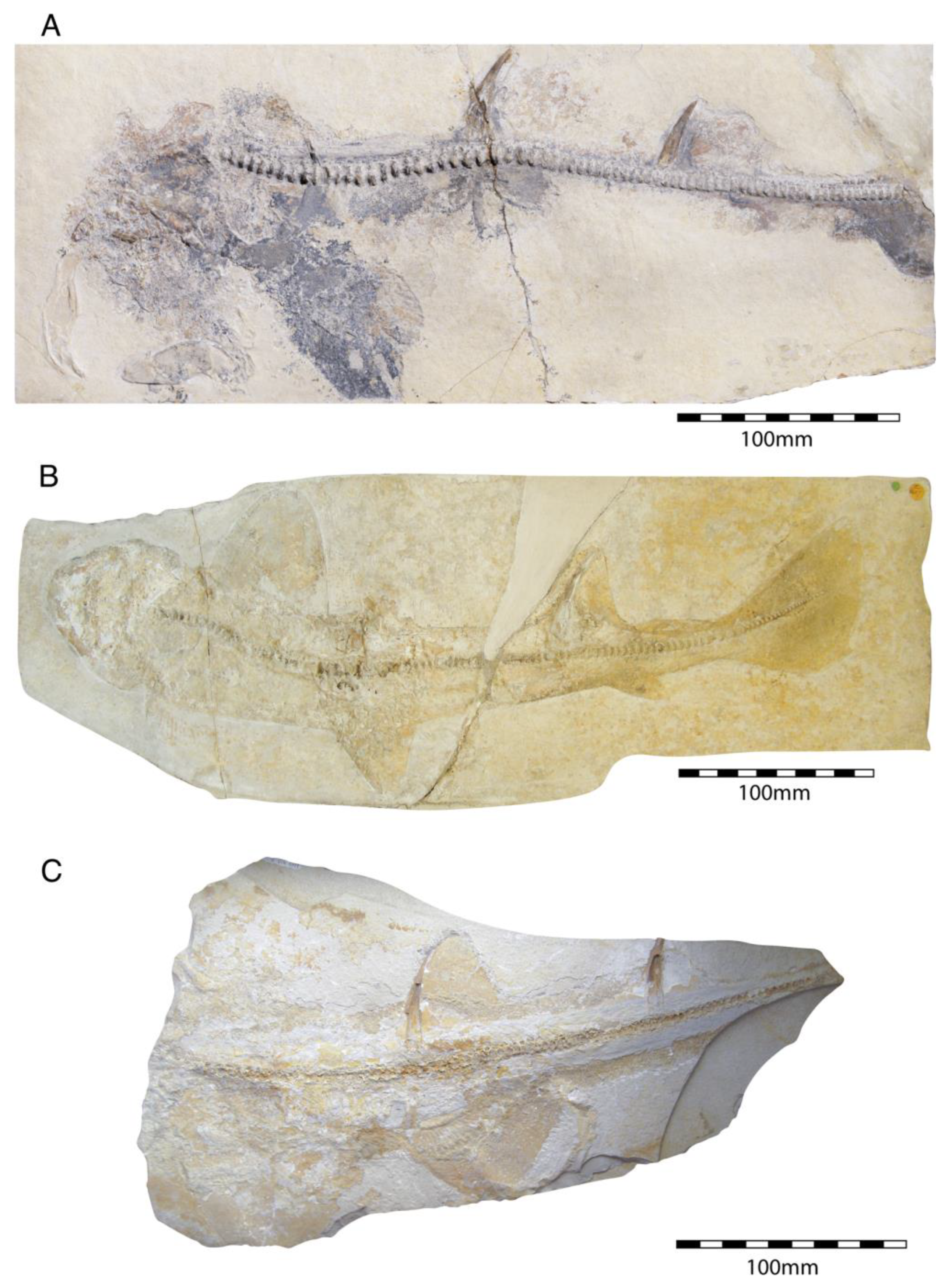
4.2. Euselachians
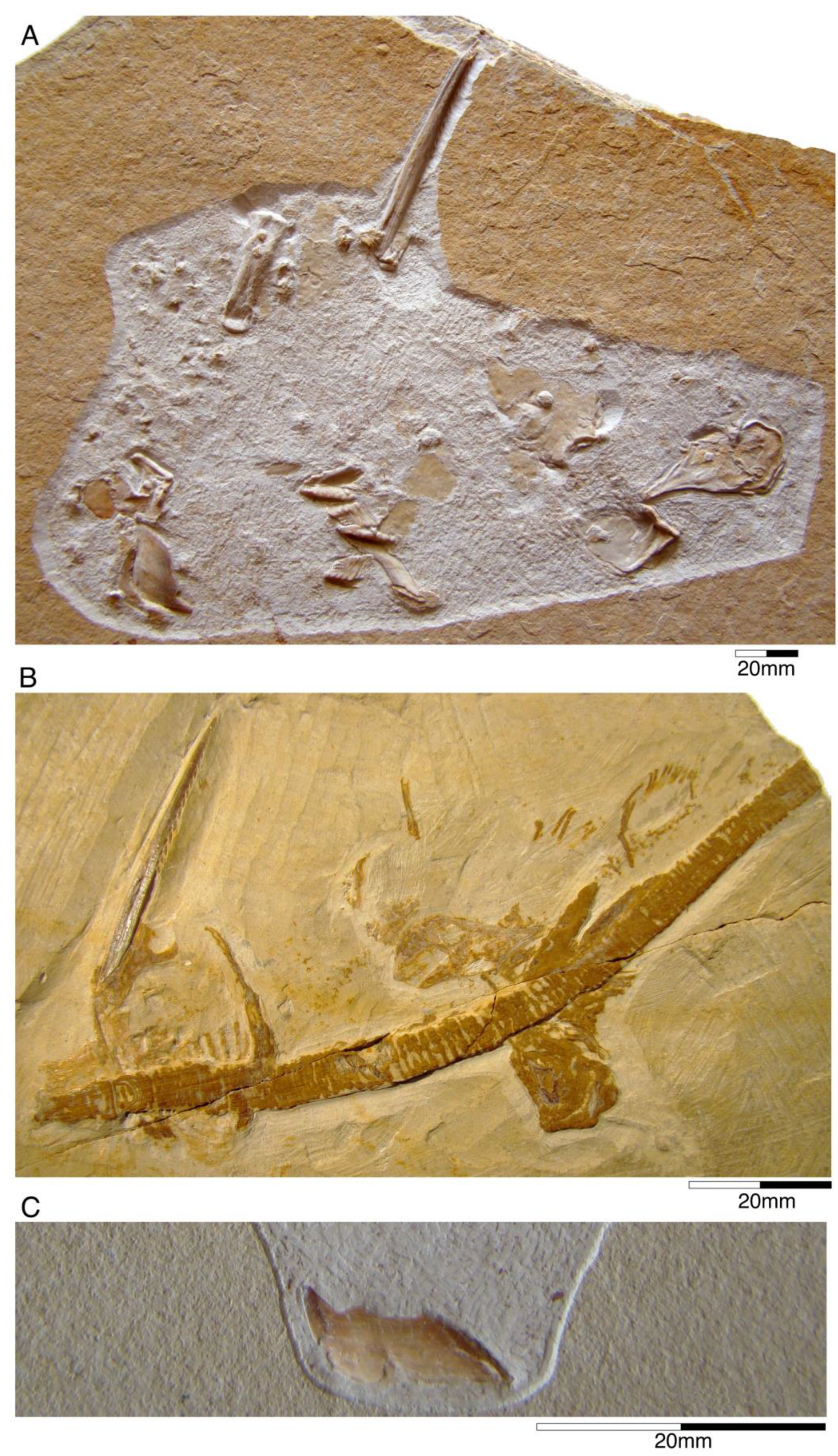
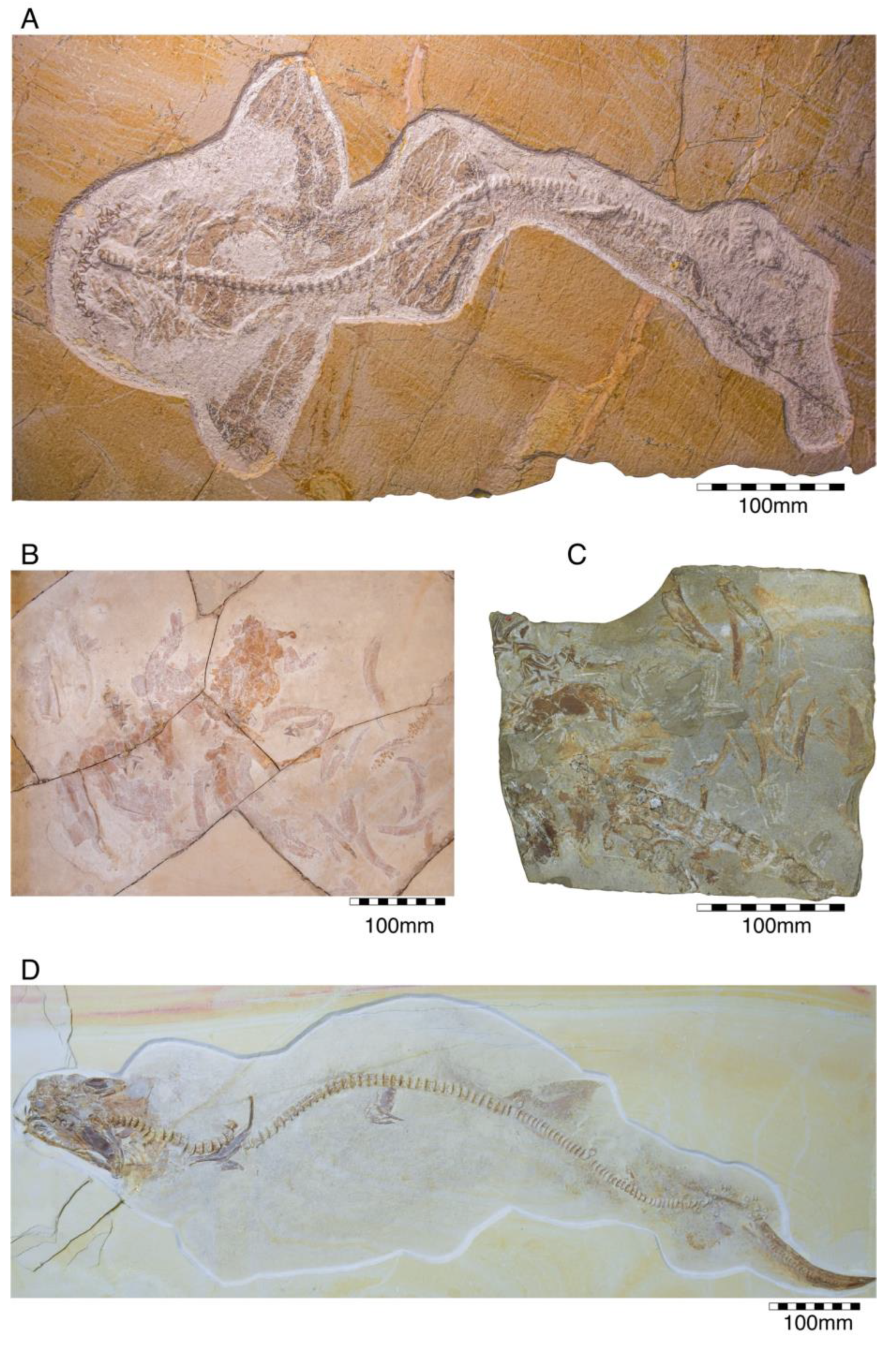
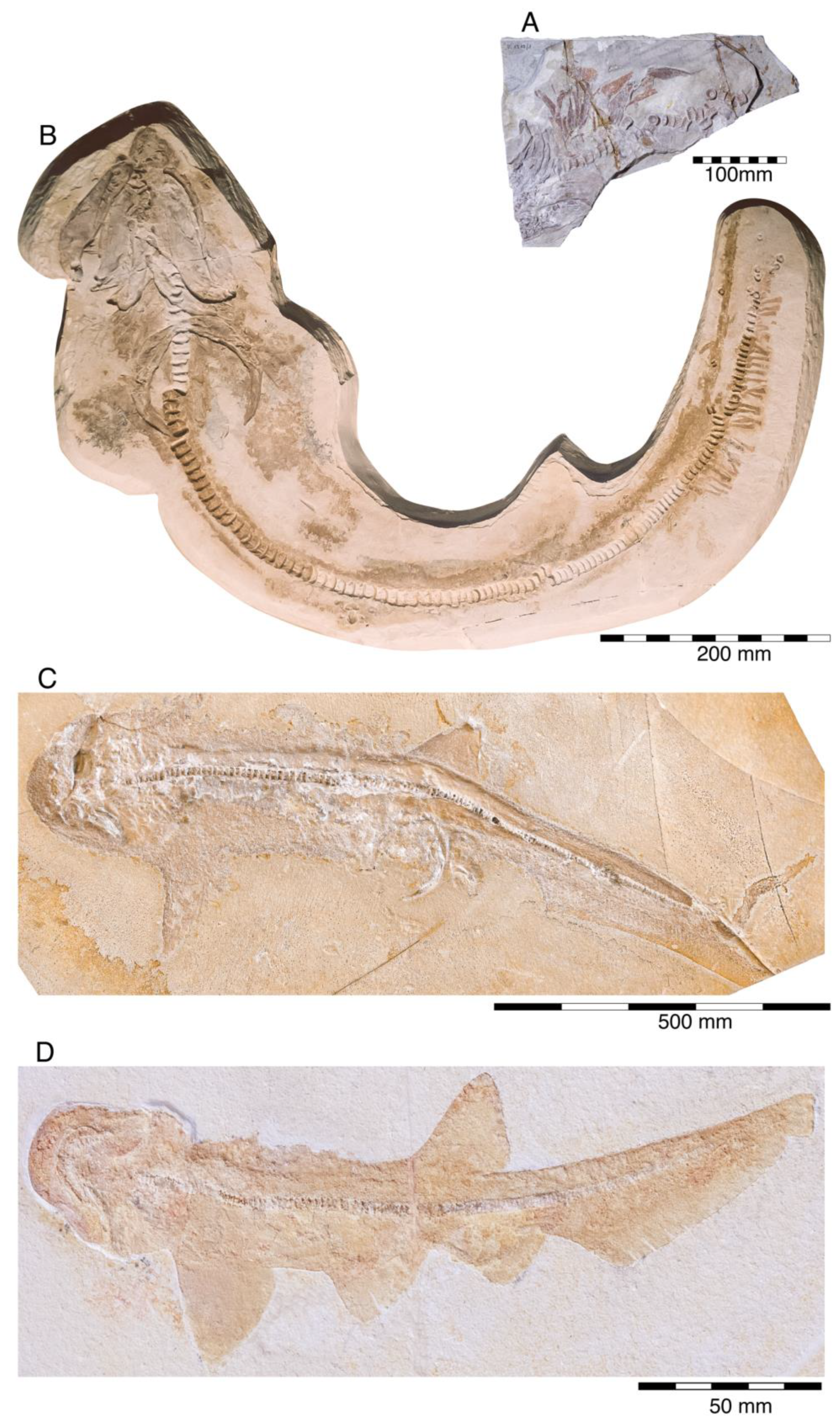
4.2.1. †Hybodontiformes
- Subclass Elasmobranchii Bonaparte, 1838 [141]
- Cohort Euselachii Hay, 1902 [139]
- Subcohort indet.
- Order †Hybodontiformes Maisey, 1975 [153]
- Superfamily †Hybodontoidea Owen, 1846 [170]
- Family †Hybodontidae Owen, 1846 [170]
- Subfamily †Hybodontinae Owen, 1846 [170]
- Genus †Hybodus Agassi, 1837 [171]
- †Hybodus fraasi Brown, 1900 [172]
- Subfamily †Acrodontinae Casier, 1959 [177]
- Genus †Asteracanthus Agassiz, 1837 [171]
- †Asteracanthus ornatissimus (Agassiz, 1837 [171])


- Genus †Strophodus Agassiz, 1838 [171]
- †Strophodus sp.

4.2.2. Subcohort Neoselachii Sensu Compagno, 1977 [21]
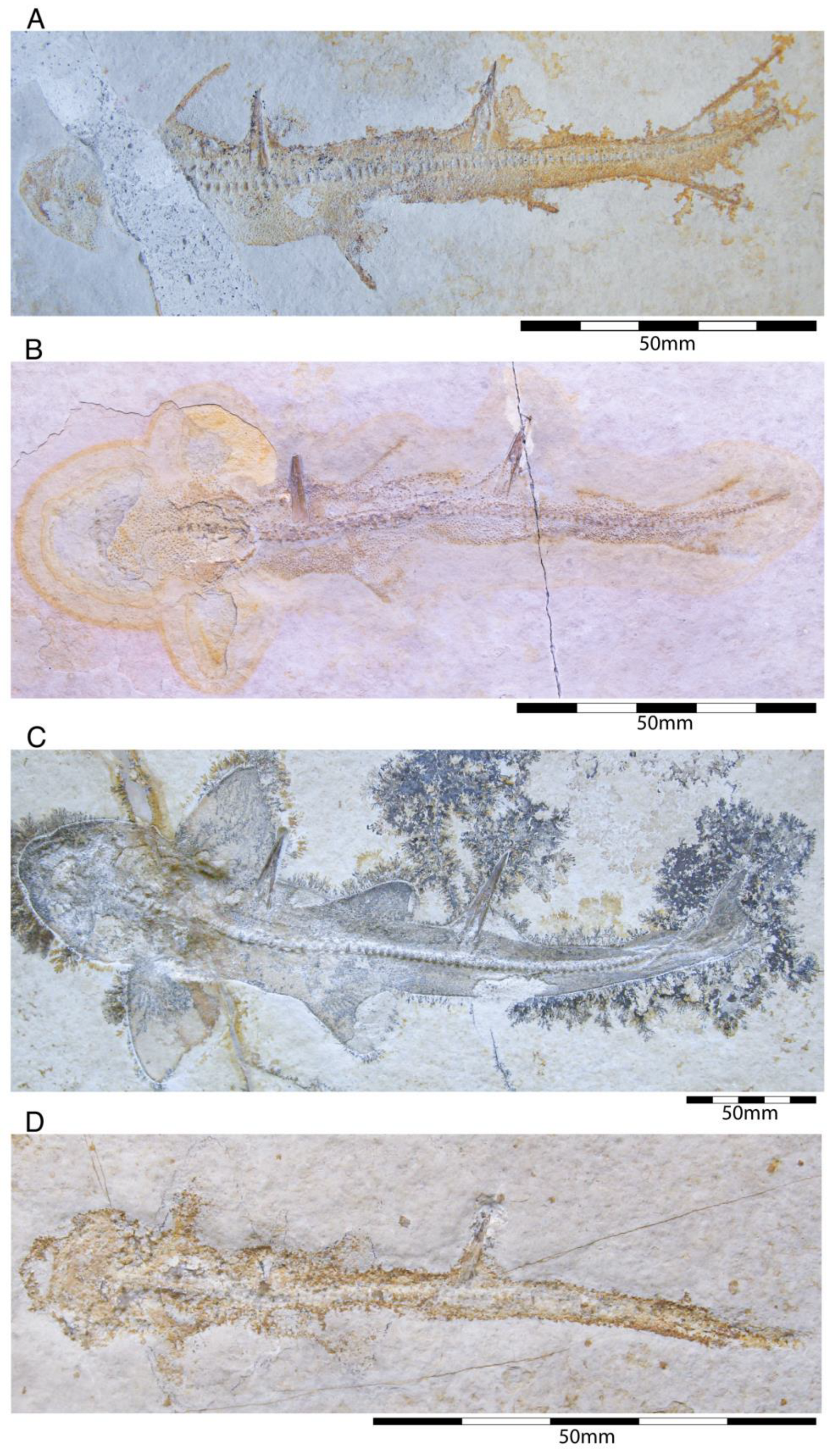
4.2.3. †Synechodontiformes
- Order †Synechodontiformes Duffin and Ward, 1993 [198]
- †Sphenodus macer (Quenstedt, 1851 [106])

- †Sphenodus nitidus Wagner, 1862 [105]
- Family †Palaeospinacidae Regan, 1906 [140]
- Genus †Synechodus Woodward, 1888 [178]
- †Synechodus ungeri Klug, 2009 [70]


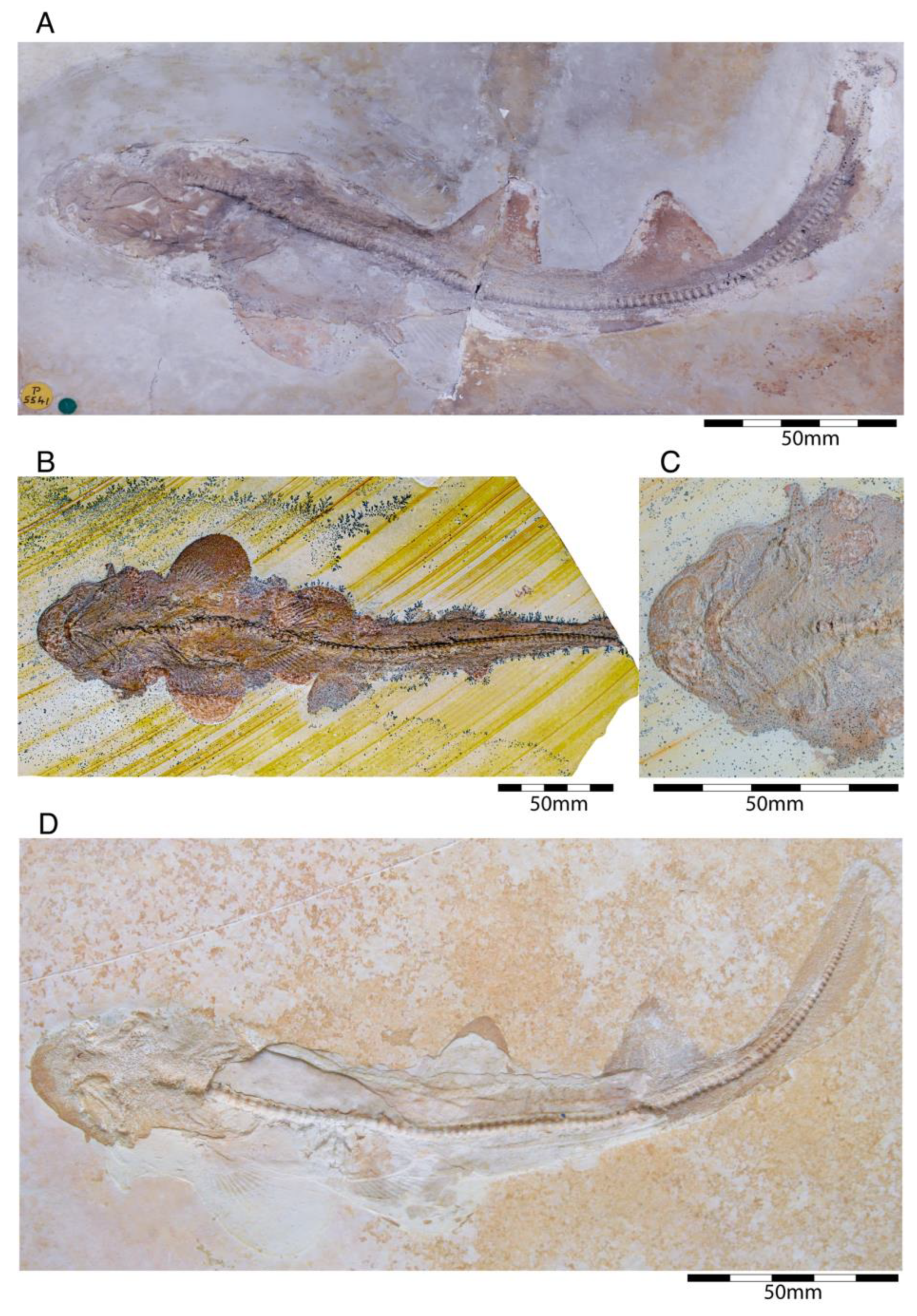
4.2.4. Heterodontiformes
- †Paracestracion falcifer Wagner, 1857 [100]
- †Paracestracion viohli Kriwet, 2008 [57]
- †Paracestracion danieli Slater, 2016 [221]

4.2.5. Orectolobiformes
- Order Orectolobiformes Applegate, 1974 [227]
- Incertae familiae
- Genus †Palaeorectolobus Kriwet, 2008 [58]
- †Palaeorectolobus agomphius Kriwet, 2008 [58]
4.2.6. Carcharhiniformes
- Order Carcharhiniformes Compagno, 1977 [21]
- Family Scyliorhinidae Gill, 1862 [239]
- Genus †Bavariscyllium Thies, 2005 [55]
- †Bavariscyllium tischlingeri Thies 2005 [55]
- Genus †Palaeoscyllium Wagner, 1857 [102]
Egg Capsules of Carcharhiniformes
4.2.7. Lamniformes
- Order Lamniformes Berg, 1958 [252]
- Incertae familiae
- Genus †Paleaocarcharias de Beaumont, 1960 [51]
- †Palaeocarcharias stromeri de Beaumont, 1960 [51]

4.2.8. Hexanchiformes
- Family †Crassodontidanidae Kriwet and Klug, 2011 [267]
- Genus †Crassodontidanus Kriwet and Klug, 2011 [267]
- †Crassodontidanus serranus Kriwet and Klug, 2011 [267]
4.2.9. Squatiniformes
- Order Squatiniformes de Buen, 1926 [262]

4.2.10. †Protospinax
- †Protospinax annectans Woodward, 1918 [232]

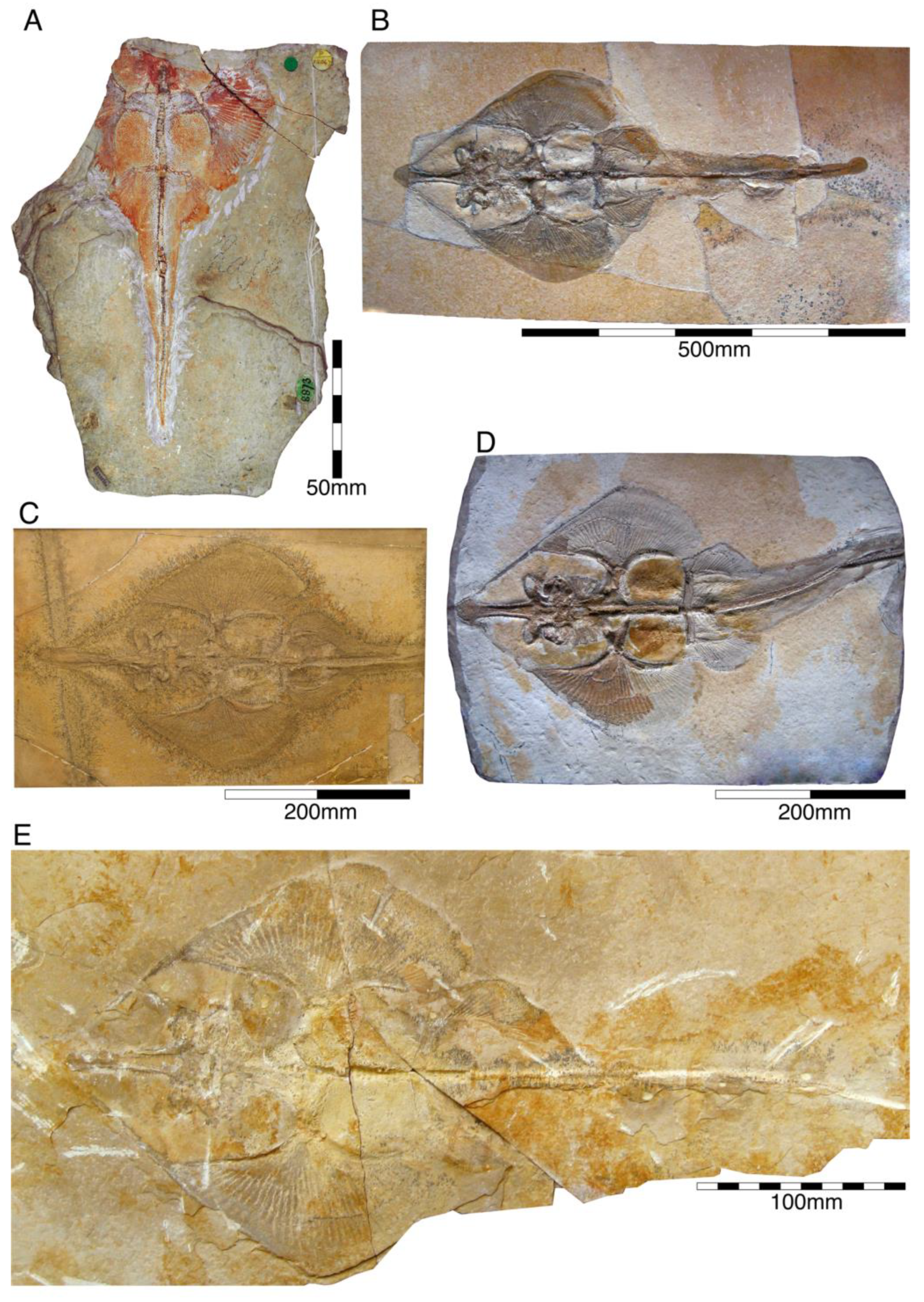
4.2.11. Batomorphii
- Superorder Batomorphii Cappetta, 1980 [302]
- Incerti ordinis
- Incertae familiae
- Genus †Asterodermus Agassiz, 1936 [171]
- †Asterodermus platypterus Agassiz, 1936 [171]

- Genus †Spathobatis Thiollière, 1852 [229]
- †Spathobatis bugesiacus Thiollière, 1852 [229]
Egg Capsules of Batomorphs
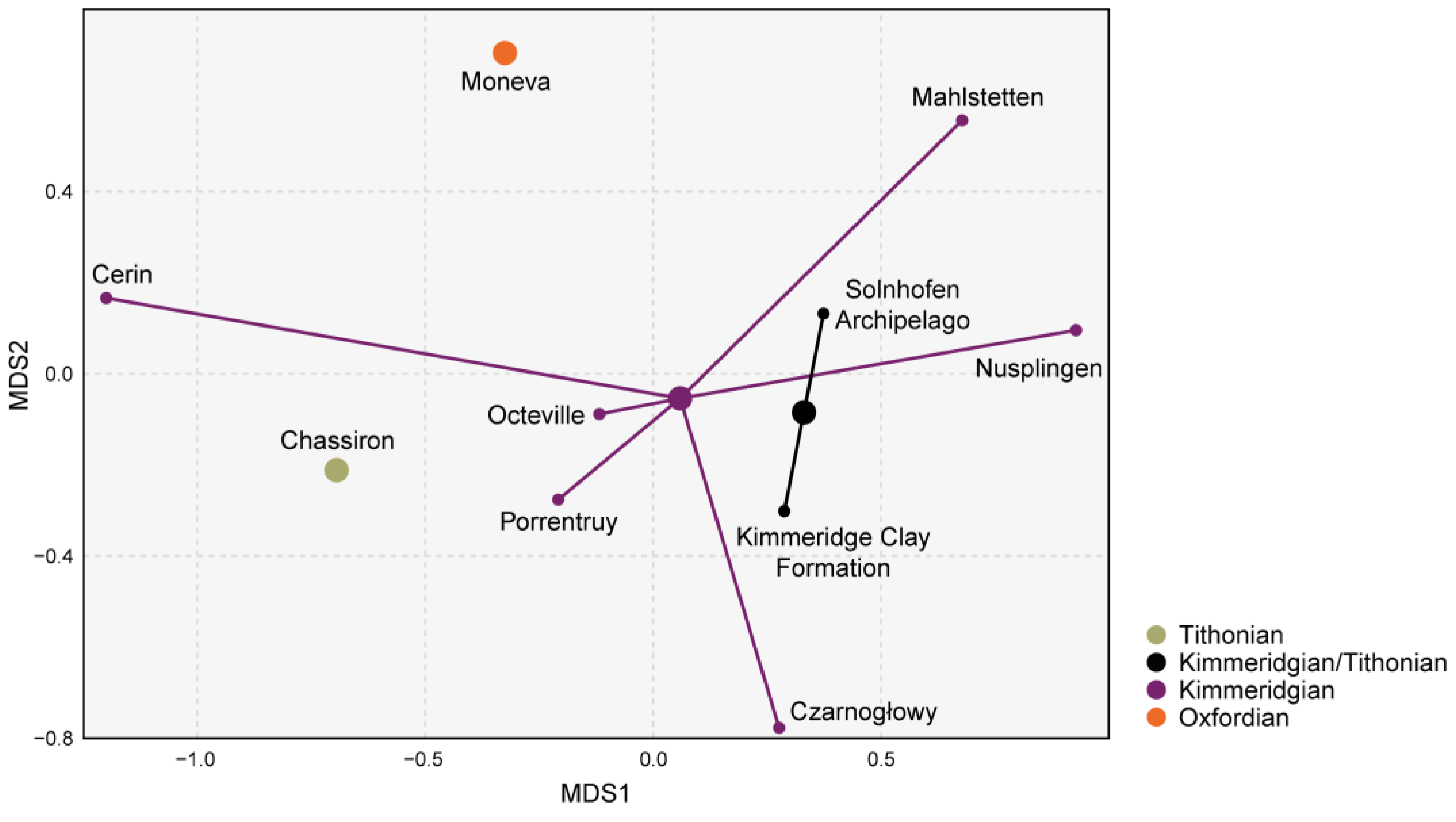
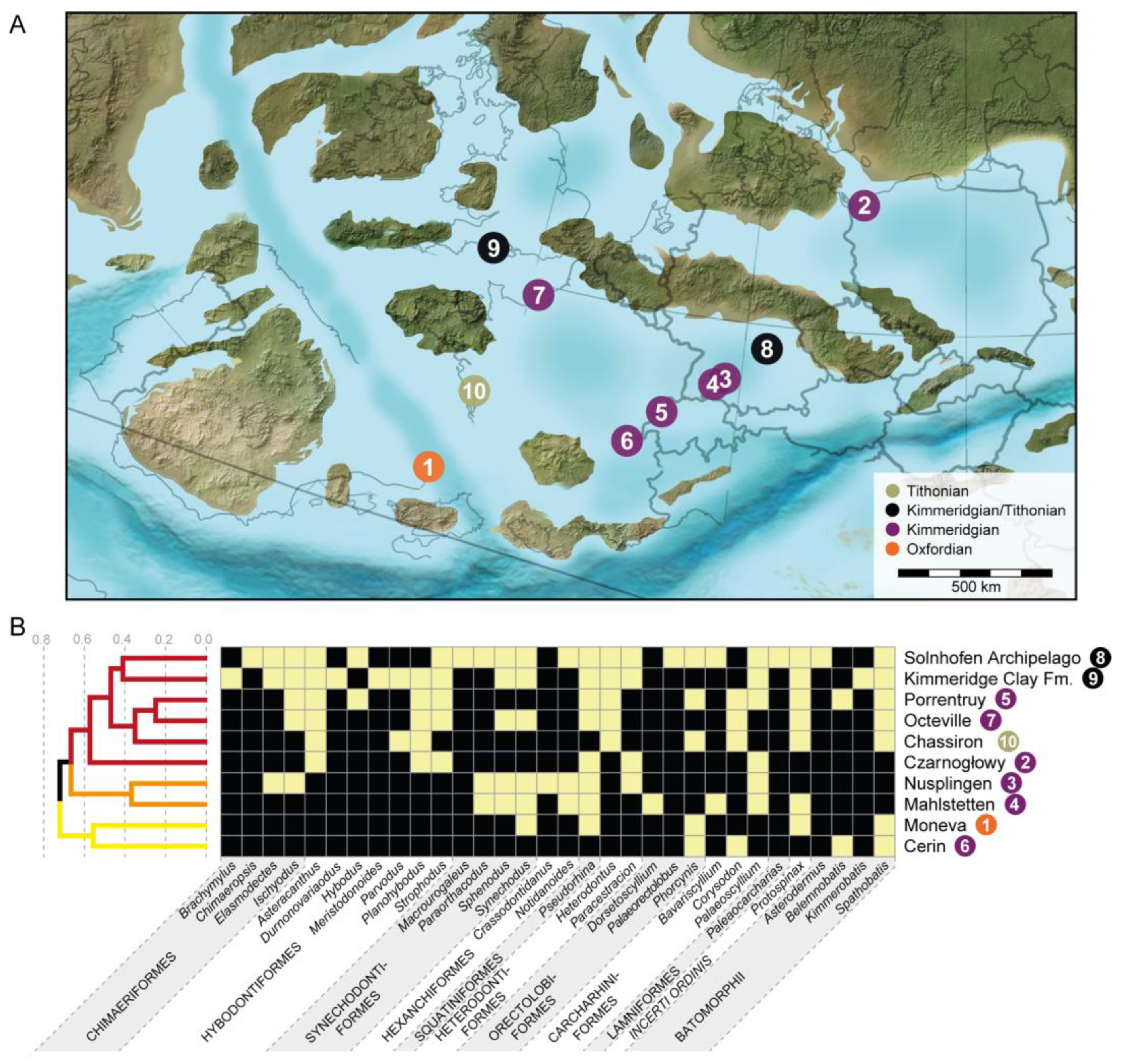
4.3. Faunal Relationships of the Solnhofen Archipelago Chondrichthyan Fauna


5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Keupp, H.; Koch, R.; Schweigert, G.; Viohl, G. Geological history of the Southern Franconian Alb–the area of the Solnhofen Lithographic Limestone. Neues Jahrb. Geol. Paläontol. Abh. 2007, 245, 3–21. [Google Scholar] [CrossRef]
- Viohl, G. Der geologische Rahmen: Die Südliche Frankenalb und ihre Entwicklung. In Solnhofen—Ein Fenster in Die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 56–62. ISBN 978-3-89937-195-6. [Google Scholar]
- Viohl, G. Die Grabung Schamhaupten. In Solnhofen—Ein Fenster in Die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 119–125. ISBN 978-3-89937-195-6. [Google Scholar]
- Peyer, K.; Charbonnier, S.; Allain, R.; Läng, É.; Vacant, R. A new look at the Late Jurassic Canjuers conservation Lagerstätte (Tithonian, Var, France). C. R. Palevol. 2014, 13, 403–420. [Google Scholar] [CrossRef]
- Bernier, P.; Barale, G.; Bourseau, J.P.; Buffetaut, E.; Gaillard, C.; Gall, J.C.; Wenz, S. The lithographic limestones of Cerin (southern Jura Mountains, France). A synthetic approach and environmental interpretation. C. R. Palevol. 2014, 13, 383–402. [Google Scholar] [CrossRef]
- Meyer, C.A.; Thüring, S.; Billon-Bruyat, J.P.; Marty, D.; Costeur, L.; Meyer, C.A.; Thüring, B. Late Jurassic marginal marine ecosystem of the Southern Jura Mountains. In Proceedings of the Abstracts and Field Guides, 5th International Symposium on Lithographic Limestone and Plattenkalk, Actes, Basel, Switzerland, 17–22 August 2009; pp. 130–141. [Google Scholar]
- Dietl, G.; Schweigert, G. Im Reich der Meerengel–Fossilien aus dem Nusplinger Plattenkalk; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2011; p. 144. ISBN 978-3-89937-139-0. [Google Scholar]
- Röper, M. Field trip C: Lithographic limestones and plattenkalk deposits of the Solnhofen and Mörnsheim formations near Eichstätt and Solnhofen. Zitteliana 2005, 26, 71–85. [Google Scholar]
- Röper, M. Field Trip B: East Bavarian Plattenkalk-different types of Upper Kimmeridgian to Lower Tithonian Plattenkalk deposits and facies. Zitteliana 2005, 26, 57–70. [Google Scholar]
- Barthel, K.W. On the deposition of the Solnhofen lithographic limestone (Lower Tithonian, Bavaria, Germany). Neues Jahrb. Geol. Paläontol. Abh. 1970, 135, 1–18. [Google Scholar]
- Arratia, G.; Schultze, H.-P.; Tischlinger, H.; Viohl, G. Solnhofen—Ein Fenster in Die Jurazeit 2; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 331–620. ISBN 978-3-89937-195-6. [Google Scholar]
- Fuchs, D.; von Boletzky, S.; Tischlinger, H. New evidence of functional suckers in belemnoid coleoids (Cephalopoda) weakens support for the ‘Neocoleoidea’ concept. J. Molluscan Stud. 2010, 76, 404–406. [Google Scholar] [CrossRef] [Green Version]
- Haug, J.T.; Haug, C.; Waloszek, D.; Schweigert, G. The importance of lithographic limestones for revealing ontogenies in fossil crustaceans. Swiss J. Geosci. 2011, 104, 85–98. [Google Scholar] [CrossRef]
- Klug, C.; Schweigert, G.; Hoffmann, R.; Weis, R.; De Baets, K. Fossilized leftover falls as sources of palaeoecological data: A ‘pabulite’ comprising a crustacean, a belemnite and a vertebrate from the Early Jurassic Posidonia Shale. Swiss J. Palaeontol. 2021, 140, 10. [Google Scholar] [CrossRef]
- Zell, P.; Beckmann, S.; Stinnesbeck, W. Liostrea roemeri (Ostreida, Bivalvia) attached to Upper Jurassic ammonites of northeastern Mexico. Paleobiodivers. Paleoenviron. 2014, 94, 439–451. [Google Scholar] [CrossRef]
- Leanza, H.; Zeiss, A. Upper Jurassic lithographic limestones from Argentina (Neuquén Basin): Stratigraphy and fossils. Facies 1990, 22, 169–185. [Google Scholar] [CrossRef]
- Tennant, J.P.; Mannion, P.D.; Upchurch, P.; Sutton, M.D.; Price, G.D. Biotic and environmental dynamics through the Late Jurassic–Early Cretaceous transition: Evidence for protracted faunal and ecological turnover. Biol. Rev. 2017, 92, 776–814. [Google Scholar] [CrossRef] [Green Version]
- Kriwet, J.; Klug, S. Late Jurassic selachians (Chondrichthyes, Elasmobranchii) from southern Germany: Re-evaluation on taxonomy and diversity. Zitteliana A 2004, 44, 67–95. [Google Scholar]
- Kriwet, J.; Klug, S. Knorpelfische (Chondrichthyes). In Solnhofen—Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 334–359. ISBN 978-3-89937-195-6. [Google Scholar]
- Compagno, L.J.V. Interrelationships of living elasmobranchs. Zool. J. Linn. Soc. 1973, 53, 15–61. [Google Scholar]
- Compagno, L.J.V. Phyletic relationships of living sharks and rays. Am. Zool. 1977, 17, 303–322. [Google Scholar] [CrossRef] [Green Version]
- Maisey, J.G. What is an ‘elasmobranch’? The impact of palaeontology in understanding elasmobranch phylogeny and evolution. J. Fish Biol. 2012, 80, 918–951. [Google Scholar] [CrossRef]
- Stahl, B.J. Chondrichthyes III—Holocephali. In Handbook of Paleoichthyology; Schultze, H.-P., Ed.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 1999; Volume 4, p. 129. [Google Scholar]
- Cappetta, H. Chondrichthyes: Mesozoic and Cenozoic Elasmobranchii. In Handbook of Paleoichthyology, Chondrichthyes; Schultze, H.-P., Ed.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2012; Volume 3E, p. 512. [Google Scholar]
- Thies, D.; Leidner, A. Sharks and guitarfishes (Elasmobranchii) from the Late Jurassic of Europe. Palaeodiversity 2011, 4, 63–184. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Malden, MA, USA, 2004; 248p. [Google Scholar]
- Goslee, S.C.; Urban, D.L. The ecodist package for dissimilarity-based analysis of ecological data. J. Stat. Softw. 2007, 22, 1–19. [Google Scholar] [CrossRef]
- Batool, F.; Hennig, C. Clustering with the average silhouette width. Comput. Stat. Data Anal. 2021, 158, 107190. [Google Scholar] [CrossRef]
- Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [Green Version]
- Minchin, P.R. An evaluation of the relative robustness of techniques for ecological ordination. Vegetation 1987, 69, 89–107. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Package ‘vegan’. Community Ecol. Package 2013, 2, 1–295. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Package ‘gplots’. In Various R Programming Tools for Plotting Data, R package version 3.1.3; 2022; Available online: https://CRAN.R-project.org/package=gplots (accessed on 3 February 2022).
- von Gümbel, C.W. Geognostische Beschreibung der Fränkischen Alb (Frankenjura mit dem Anstoßenden Fränkischen Keupergebiet); Theodor Fischer: Kassel, Germany, 1891. [Google Scholar]
- Fürsich, F.T.; Mäuser, M.; Schneider, S.; Werner, W. The Wattendorf Plattenkalk (Upper Kimmeridgian)—A new conservation lagerstätte from the northern Franconian Alb, southern Germany. Neues Jahrb. Geol. Paläontol. Abh. 2007, 245, 45–58. [Google Scholar] [CrossRef]
- Mäuser, M. Frankenland am Jurastrand—Versteinerte Schätze aus der Wattendorfer Lagune, 2nd ed.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2014; 60p, ISBN 978-3-89937-171-0. [Google Scholar]
- Mäuser, M. Oberjurassische Fossil-Lagerstätten außerhalb der Südlichen Frankenalb. Die laminierten Plattenkalke von Wattendorf in Oberfranken. In Solnhofen—Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 515–535. ISBN 978-3-89937-195-6. [Google Scholar]
- Niebuhr, B.; Pürner, T. Plattenkalk und Frankendolomit–Lithostratigraphie der Weißjura-Gruppe der Frankenalb (außeralpiner Oberjura, Bayern). SDGG 2014, 83, 5–72. [Google Scholar] [CrossRef]
- Joyce, W.G.; Mäuser, M. New material of named fossil turtles from the Late Jurassic (late Kimmeridgian) of Wattendorf. PLoS ONE 2019, 15, e0233483. [Google Scholar] [CrossRef] [PubMed]
- Sachs, S.; Young, M.T.; Abel, P.; Mallison, H. A new species of the metriorhynchid crocodylomorph Cricosaurus from the Upper Jurassic of southern Germany. Acta Palaeontol. Pol. 2019, 64, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Wellnhofer, P. Archaeopteryx—Der Urvogel von Solnhofen; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2008; p. 256. ISBN 978-3-89937-076-8. [Google Scholar]
- Foth, C.; Tischlinger, H.; Rauhut, O.W.M. New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature 2014, 511, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Rauhut, O.W.M.; Foth, C.; Tischlinger, H. The oldest Archaeopteryx (Theropoda: Avialiae): A new specimen from the Kimmeridgian/Tithonian boundary of Schamhaupten, Bavaria. PeerJ 2018, 6, e4191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweigert, G. Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany—First results and open questions. Neues Jahrb. Geol. Paläontol. Abh. 2007, 245, 117–125. [Google Scholar] [CrossRef]
- Schweigert, G. Biostratigraphie der Plattenkalke der Südlichen Frankenalb. In Solnhofen—Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 63–66. ISBN 978-3-89937-195-6. [Google Scholar]
- Viohl, G. The paleoenvironment of the Late Jurassic fishes from the southern Franconian Alb (Bavaria, Germany). In Mesozoic Fishes—Systematics and Paleoecology; Arratia, G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 1996; Volume 1, pp. 513–528. ISBN 3-923871-90-2. [Google Scholar]
- Neumeyer, O. Solnhofener Naturstein als Werkstoff: Geschichte, Abbau und Verwendung. In Solnhofen—Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 19–27. ISBN 978-3-89937-195-6. [Google Scholar]
- Viohl, G.; Zapp, M. Schamhaupten, an outstanding fossil-Lagerstätte in a silicified Plattenkalk around the Kimmeridgian-Tithonian boundary (Southern Franconian Alb, Bavaria). Neues Jahrb. Geol. Paläontol. Abh. 2007, 245, 127–142. [Google Scholar] [CrossRef]
- Heyng, A.M.; Leonhardt, U.; Krautworst, U.; Pöschl, R. Die Mörnsheimer Schichten am Schaudiberg. In Solnhofen—Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 137–152. ISBN 978-3-89937-195-6. [Google Scholar]
- Rauhut, O.W.M.; López-Arbarello, A.; Röper, M.; Rothgaenger, M. Vertebrate fossils from the Kimmeridgian of Brunn: The oldest fauna from the Solnhofen Archipelago (Late Jurassic, Bavaria, Germany). Zitteliana 2017, 89, 305–329. [Google Scholar]
- Münster, G. Beschreibung einiger neuen Fische aus der Jura-Formation von Bayern und Württemberg. Beiträge Zur Petrefactenkunde 1842, 5, 55–64. [Google Scholar]
- de Beaumont, G. Observations préliminaires sur trois Sélaciens nouveaux du Calcaire lithographique d’Eichstätt (Bavière). Eclogae Geol. Helv. 1960, 53, 315–328. [Google Scholar]
- Maisey, J.G. Anatomical revision of the fossil shark Hybodus fraasi (Chondrichthyes, Elasmobranchii). Am. Mus. Novit. 1986, 2857, 1–16. [Google Scholar]
- Duffin, C.J. The Upper Jurassic selachian Palaeocarcharias de Beaumont (1960). Zool. J. Linn. Soc. 1988, 94, 271–286. [Google Scholar] [CrossRef]
- Kriwet, J.; Klug, S. Dental patterns of the stem-group hexanchoid shark, Notidanoides muensteri (Elasmobranchii, Hexanchiformes). J. Vertebr. Paleontol. 2014, 34, 1292–1306. [Google Scholar] [CrossRef]
- Thies, D. A catshark (Neoselachii, Carcharhiniformes, Scyliorhinidae) from the Late Jurassic of Germany. Paläontol. Z. 2005, 79, 339–348. [Google Scholar] [CrossRef]
- Klug, S. The Late Jurassic neoselachian Macrourogaleus Fowler, 1947 is a palaeospinacid shark (Elasmobranchii; Synechodontiformes). Acta Geol. Pol. 2008, 58, 229–234. [Google Scholar]
- Kriwet, J. A new species of extinct bullhead sharks, Paracestracion viohli sp. nov. (Neoselachii, Heterodontiformes), from the Upper Jurassic of South Germany. Acta Geol. Pol. 2008, 58, 235–241. [Google Scholar]
- Kriwet, J. A Late Jurassic carpetshark (Neoselachii, Orectolobiformes) from southern Germany. In Mesozoic Fishes—Homology and Phylogeny; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2008; Volume 4, pp. 443–453. ISBN 978-3-89937-080-5. [Google Scholar]
- Slater, T.S.; Ashbrook, K.; Kriwet, J. Evolutionary relationships among bullhead sharks (Chondrichthyes, Heterodontiformes). Pap. Palaeontol. 2020, 6, 425–437. [Google Scholar] [CrossRef]
- Stumpf, S.; López-Romero, F.A.; Kindlimann, R.; Lacombat, F.; Pohl, B.; Kriwet, J. A unique hybodontiform skeleton provides novel insights into Mesozoic chondrichthyan life. Pap. Palaeontol. 2021, 7, 1479–1505. [Google Scholar] [CrossRef]
- Dietl, G.; Schweigert, G.; Franz, M.; Geyer, M. Profile des Nusplinger Plattenkalks (Oberjura, Schwäbische Alb). Stuttg. Beitr. Naturk. B 1998, 265, 1–37. [Google Scholar]
- Schweigert, G.; Roth, S. The Nusplingen Plattenkalk—A shark lagoon in the Late Jurassic of the Swabian Alb Geopark. Geoconserv. Res. 2021, 4, 347–356. [Google Scholar] [CrossRef]
- Dietl, G.; Schweigert, G. The Nusplingen Lithographic Limestone—A “fossil lagerstaette” of Late Kimmeridgian age from the Swabian Alb (Germany). Riv. Ital. Paleontol. S. 2004, 110, 303–309. [Google Scholar] [CrossRef]
- Fraas, O. Squatina acanthoderma, Der Meerengel von Nusplingen. Z. Dtsch. Geol. Ges. 1854, 6, 782–799. [Google Scholar]
- Fraas, O. Beiträge zum obersten weissen Jura in Schwaben. Jahresh. Ver. vaterl. Naturkd. Württemb. 1855, 11, 76–107. [Google Scholar]
- Schweizer, R. Die Elasmobranchier und Holocephalen aus den Nusplinger Plattenkalken. Palaeontogr. Abt. A 1964, 123, 58–110. [Google Scholar]
- Duffin, C.J. The palaeospinacid shark “Synechodus” jurensis Schweizer, 1964 from the Late Jurassic of Germany. Belg. Geol. Surv. Prof. Pap. 1993, 264, 157–174. [Google Scholar]
- Böttcher, R.; Duffin, C.J. The neoselachian shark Sphenodus from the Late Kimmeridgian (Late Jurassic) of Nusplingen and Egesheim (Baden-Württemberg, Germany). Stutt. Beitr. Naturkd. B (Geol. Und Paläontologie) 2000, 283, 1–31. [Google Scholar]
- de Carvalho, M.R.; Kriwet, J.; Thies, D. A systematic and anatomical revision of Late Jurassic angelsharks (Chondrichthyes: Squatinidae). In Mesozoic Fishes—Homology and Phylogeny; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2008; Volume 4, pp. 469–502. ISBN 978-3-89937-080-5. [Google Scholar]
- Klug, S. A new palaeospinacid shark (Chondrichthyes, Neoselachii) from the Upper Jurassic of southern Germany. J. Vertebr. Paleontol. 2009, 29, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Klug, S.; Kriwet, J.; Böttcher, R.; Schweigert, G.; Dietl, G. Skeletal anatomy of the extinct shark Paraorthacodus jurensis (Chondrichthyes; Palaeospinacidae), with comments on synechodontiform and palaeospinacid monophyly. Zool. J. Linn. Soc. 2009, 157, 107–134. [Google Scholar] [CrossRef] [Green Version]
- Ebert, M.; Kölbl-Ebert, M.; Lane, J.A. Fauna and Predator-Prey Relationships of Ettling, an Actinopterygian Fish-Dominated Konservat-Lagerstätte from the Late Jurassic of Southern Germany. PLoS ONE 2015, 10, e0116140. [Google Scholar] [CrossRef] [PubMed]
- Tischlinger, H.; Schweigert, G. Neuer Ammonitenfund ermöglicht Alterseinstufung der Fossillagerstätte Ettling. Archaeopteryx 2020, 36, 54–63. [Google Scholar]
- Darras, L.; Derycke, C.; Blieck, A.; Vachard, D. The oldest holocephalan (Chondrichthyes) from the Middle Devonian of the Boulonnais (Pas-de-Calais, France). C. R. Palevol 2008, 7, 297–304. [Google Scholar] [CrossRef]
- Didier, D.A.; Kemper, J.M.; Ebert, D.A. Phylogeny, Biology, and Classification of Extant Holocephalans. In Biology of Sharks and Their Relatives, 2nd ed.; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 97–124. ISBN 9780429106545. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016; 707p, ISBN 9781118342336. [Google Scholar]
- Pradel, A.; Langer, M.; Maisey, J.G.; Geffard-Kuriyama, D.; Cloetens, P.; Janvier, P.; Tafforeau, P. Skull and brain of a 300-million-year-old chimaeroid fish revealed by synchrotron holotomography. Proc. Natl. Acad. Sci. USA 2009, 106, 5224–5228. [Google Scholar] [CrossRef] [Green Version]
- Grogan, E.D.; Lund, R. The Origin and Relationships of Early Chondrichthyes. In Biology of Sharks and Their Relatives, 1st ed.; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 3–31. ISBN 0-8493-1514-X. [Google Scholar]
- Duffin, C. Plattenkalk Late Jurassic chimaeroids. In Extended Abstracts of the Fourth International Meeting on Mesozoic Fishes–Systematics, Homology, and Nomenclature. Miraflores de la Sierra, Madrid, Spain; Poyato-Ariza, F.J., Ed.; Ediciones UAM: Madrid, Spain, 2005; pp. 83–89. [Google Scholar]
- de Saint-Seine, P. Les poissons des calcaires lithographiques de Cerin (Ain). Nouv. Arch. Mus. Hist. Nat. Lyon 1949, 2, 1–357. [Google Scholar] [CrossRef]
- Wenz, S.; Bernier, P.; Barale, G.; Bourseau, J.-P.; Buffetaut, E.; Gaillard, C.; Gall, J.-C. L’Ichthyofaune des calcaires lithographiques du Kimméridgien supérieur de Cerin (Ain, France). Geobios 1994, 16, 61–70. [Google Scholar] [CrossRef]
- Leuzinger, L.; Cuny, G.; Popov, E.; Billon-Bruyat, J.P. A new chondrichthyan fauna from the Late Jurassic of the Swiss Jura (Kimmeridgian) dominated by hybodonts, chimaeroids and guitarfishes. Pap. Palaeontol. 2017, 3, 471–511. [Google Scholar] [CrossRef] [Green Version]
- Jerve, A.; Johanson, Z.; Ahlberg, P.; Boisvert, C. Embryonic development of fin spines in Callorhinchus milii (Holocephali); implications for chondrichthyan fin spine. Evol. Dev. 2014, 16, 339–353. [Google Scholar] [CrossRef]
- Huxley, T.H. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proc. Zool. Soc. Lond. 1880, 1880, 649–662. [Google Scholar]
- Zangerl, R. New chondrichthyes from the Mazon Creek Fauna (Pennsylvanian) of Illinois. In Mazon Creek Fossils; Nitecki, M.H., Ed.: Academic Press: New York, NY, USA, 1979; pp. 449–500. [Google Scholar]
- Bonaparte, C.L. Iconografia della fauna italica per le quattro classi degli animali vertebrati. Tomo III. Pesci. Roma 1832, 1, 1–6. [Google Scholar]
- Obruchev, D.V. Izuchenie edestid y raboty A. P. Karpinskogo. Trudy Paleontol. Inst. Akad. Nauk SSSR 1953, 45, 1–85. (In Russian) [Google Scholar]
- Patterson, C. The phylogeny of the chimaeroids. Philos. Trans. Proc. Royal. Soc. B 1965, 249, 101–219. [Google Scholar]
- von Zittel, K.A. Handbuch der Paläontologie. 1. Abtheilung Paläozoologie, 3: Vertebrata (Pisces, Amphibia, Reptilia, Aves); Oldenbourg: München, Leipzig, Germany, 1887; p. 900. [Google Scholar]
- Münster, G.G. Ueber eine neue Art Myriacanthus, Agassiz. Im oberen Jura-Kalk. Beiträge Zur Petrefactenkunde 1840, 3, 127–128. [Google Scholar]
- Braun, K.F.W. Verzeichniss der in der Kreis-Naturalien-Sammlung zu Bayreuth Befindlichen Petrefacten: Mit, I. Einer Geognostisch-Petrefactologischen Karte von Ober-Franken. II. Einer Geognostisch-Petrefactischen Uebersicht. III. Zweiundzwanzig Tafeln Abbildungen; Verlag von Leopold Voss: Leipzig, Germany, 1840; p. 118. [Google Scholar]
- Giebel, C.G. Fauna der Vorwelt mit Steter Berücksichtigung der Lebenden Thiere. Erster Band: Wirbelthiere. Dritte Abtheilung: Fische; Brockhaus: Leipzig, Germany, 1848; p. 467. [Google Scholar]
- Duffin, C.J. A new myriacanthid holocephalian from the Sinemurian (Lower Jurassic) of Belgium. Zool. J. Linn. Soc. 1984, 82, 55–71. [Google Scholar] [CrossRef]
- von Meyer, H. Die Knochen auf der Platte Tafel VIII. fig. 1. Beiträge zur Petrefacten-Kunde 1839, 1, 83–84. [Google Scholar]
- Riess, J. Ueber einige fossile Chimeriden-Reste im Münchener palaeontologischen Museum. Palaeontographica 1887, 34, 1–27. [Google Scholar]
- Woodward, A.S. Catalogue of the Fossil Fishes in the British Museum (Natural History); Part II. containing the Elasmobranchii (Acanthodii). Holocephali, Ichthyodorulites, Ostracodermi, Dipnoi, and Teleostomi (Crossopterygii and chondrostean Actinopterygii). 1981, p. 567. Available online: https://www.biodiversitylibrary.org/bibliography/161983 (accessed on 25 February 2023).
- Lauer, B.; Popov, E.V.; Duffin, C.; Ward, D.J.; Lauer, R.H. A new holomorphic specimen of the rare holocephalian fish, Chimaeropsis paradoxa, from the Late Jurassic Plattenkalk of Germany. In Proceedings of the Meeting Program and Abstracts of the 78th Annual Meeting of the Society of Vertebrate Paleontology, Albuquerque, NM, USA, 17–20 October 2018; p. 165. [Google Scholar]
- Duffin, C.J.; Lauer, B.; Lauer, R. Chimaeroid egg cases from the Late Jurassic of the Solnhofen area (S Germany). Neues Jahrb. Geol. Paläontol. Abh. 2022, 306, 161–175. [Google Scholar] [CrossRef]
- Garman, S. Genera and families of the chimaeroids. Proc. N. Engl. ZoClub. 1901, 2, 75–77. [Google Scholar]
- Egerton, P. On some new species of fossil chimaeroid fishes, with remarks on their general affinities. Proc. Geol. Soc. Lond. 1843, 4, 197–199. [Google Scholar]
- Buckland, W. A Notice on the Fossil Beaks of four extinct Species of Fishes, referrible to the Genus Chimaera, that occur in the Oolitic and Cretaceous formations of England. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1936, 8, 4–7. [Google Scholar] [CrossRef]
- Wagner, J.A. Charakteristik neuer Arten von Knorpelfischen aus den lithographischen Schiefern der Umgegend von Solnhofen. Gel. Anz. Königl. Bayer. Akad. Wiss. 1857, 44, 288–293. [Google Scholar]
- Quenstedt, F.A. Der Jura; Laupp: Tübingen, Germany, 1858; p. 842. [Google Scholar]
- Philippi, E. Über Ischyodus suevicus n. sp. Ein Beitrag zur Kenntnis der fossilen Holocephalen. Palaeontographica 1897, 44, 1–10. [Google Scholar]
- Wagner, J.A. Monographie der fossilen Fische aus den lithographischen Schiefern Bayerns. I. Plakoiden und Pyknodonten. Abh. Math.-Physic. Cl. Königl. Bayer. Akad. Wiss. 1862, 9, 277–352. [Google Scholar]
- Quenstedt, F.A. Handbuch der Petrefaktenkunde; Tübingen, Mohr Siebeck Verlag, H. Laupp’sche Buchhandlung: Tübingen, Germany, 1851–1852; p. 1239. [Google Scholar]
- Wagner, J.A. Monographie der fossilen Fische aus den lithographischen Schiefern Bayerns. Abh. Math.-Physic. Cl. Königl. Bayer. Akad. Wiss. 1861, 9, 16–20. [Google Scholar]
- Kriwet, J.; Klug, S. An embryonic mandibular tooth plate and associated kiremains of a Late Jurassic chimaeroid (Holocephali, Chimaeriformes) from the Iberian Peninsula. J. Vertebr. Paleontol. 2011, 31, 954–961. [Google Scholar] [CrossRef]
- Schauinsland, H.H. Beiträge zur Entwicklungsgeschichte und Anatomie der Wirbelthiere, I; Sphenodon, Callorhynchus, Chamaeleo. Zoologica. 1903, Volume 39, pp. 1–98. Available online: https://www.biodiversitylibrary.org/bibliography/11952 (accessed on 25 February 2023).
- Didier, D.A. Phylogenetic systematics of extant chimaeroid fishes (Holocephali, Chimaeroidei). Am. Mus. Novit. 1995, 3119, 1–86. Available online: http://hdl.handle.net/2246/3652 (accessed on 14 April 2022).
- von Ammon, L. Ueber neue Stücke von Ischyodus. Ber. Des Nat. Ver. Regensbg. 1896, 5, 253–263. [Google Scholar]
- Heimberg, G. Neue Fischfunde aus dem Weißen Jura ζ von Württemberg. Palaeontogr. Abt. A 1949, 97, 75–98. [Google Scholar]
- Popov, E.V.; Ward, D.J.; Lepage, G. The diversity and nomenclatural revision of the Holocephalian fishes (Chimaeriformes) from the Kimmeridgian (Late Jurassic) of Western Europe. J. Vertebr. Paleontol. 2009, 29, 166A. [Google Scholar]
- Popov, E.V.; Shapovalov, K.M. A New Genus of Elephant Fish (Holocephali: Callorhinchidae) from the Late Jurassic of Central Russia. Paleontol. J. 2021, 55, 410–420. [Google Scholar] [CrossRef]
- Mäuser, M. Frankenland am Jurastrand—Versteinerte Schätze aus der Wattendorfer Lagune (Führer zur gleichnamigen Ausstellug des Naturkunde-Museums Bamberg); Verlag Dr. Friedrich Pfeil: Munich, Germany, 2008; 60p, ISBN 9783899370973. [Google Scholar]
- Werdelin, L. A new chimaeroid fish from the Cretaceous of Lebanon. Geobios 1986, 19, 393–397. [Google Scholar] [CrossRef]
- Herman, J.; Hovestadt-Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the study of the comparative morphology of teeth and other relevant ichthyodorulites in living supraspecific taxa of Chondrichthyan fishes. Part C: Holocephali 1: Order Chimaeriformes—Suborder Chimaeroidei—Family Callorhynchidae—Subfamiliy Callorhynchinae—Genus: Callorhinchus, Family Chimaeridae—Genera: Chimaera and Hydrolagus, Family Rhinochimaeridae—Genera: Harriotta, Neoharriotta, and Rhinochimaera. Bull. Inst. Royal Sci. Nat. Belg. Biol. 2001, 71, 5–35. [Google Scholar]
- Smith, M.M.; Underwood, C.; Goral, T.; Healy, C.; Johanson, Z. Growth and mineralogy in dental plates of the holocephalian Harriotta raleighana (Chondrichthyes): Novel dentine and conserved patterning combine to create a unique chondrichthyan dentition. Zool. Lett. 2019, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.; Manzanares, E.; Underwood, C.; Healy, C.; Clark, B.; Johanson, Z. 2020. Holocephalian (Chondrichthyes) dental plates with hypermineralized dentine as a substitute for missing teeth through developmental plasticity. J. Fish Biol. 2020, 97, 16–27. [Google Scholar] [CrossRef]
- von Meyer, H. Chimaera (Ganodus) avita aus dem lithographischen Schiefer von Eichstätt. Palaeontogr. Abt. A 1862, 10, 87–95. [Google Scholar]
- Schweigert, G.; Dietl, G.; Dietl, O.; Kapitzke, M.; Rieter, M.; Ilg, A.; Russ, B. Der Nusplinger Plattenkalk (Weisser Jura ζ)—Grabungskampagne 2011. Jahresh. Ver. Vaterl. Naturkd. Württemb 2012, 168, 53–73. [Google Scholar]
- Duffin, C.J. A callorhynchid chimaeroid (Pisces, Holocephali) from the Nusplingen Plattenkalk (Late Jurassic, SW Germany). Neues Jahrb. Geol. Paläontol. Abh. 2018, 289, 161–175. [Google Scholar] [CrossRef]
- Duffin, C.J.; Lars, J. A fin spine of Recurvacanthus (Myriacanthidae, Holocephali) from the Posidonienschiefer (Early Jurassic) of SW Germany. Neues Jahrb. Geol. Paläontol. Abh. 2020, 296, 317–326. [Google Scholar] [CrossRef]
- Newton, E.T. The chimaeroid fishes of the British Cretaceous rocks. Mem. Geol. Surv. UK Monogr. 1878, 4, 1–64. [Google Scholar]
- von Meyer, H. No title. Neues Jahrb. Min. Geog. Geol. Pet. 1860, 210–212. [Google Scholar]
- Lauer, B.H.; Lauer, R.H.; Bernard, E.; Duffin, C.J.; Popov, E.V.; Ward, D.J. Observations on the Mesozoic chimeroid Elasmodectes Newton 1878. In Proceedings of the Abstracts of Papers of the 79th Annual Meeting of the Society of Vertebrate Paleontology, Brisbane, QLD, Australia, 9–12 October 2019; Volume 138. [Google Scholar]
- von Ammon, L. Ein schönes Exemplar von Ischyodus avitus. Geogn. Jahresh. 1899, 12, 158–160. [Google Scholar]
- Fischer, J.; Licht, M.; Kriwet, J.; Schneider, J.W.; Buchwitz, M.; Bartsch, P. Egg capsule morphology provides new information about the interrelationships of chondrichthyan fishes. J. Syst. Palaeontol. 2014, 12, 389–399. [Google Scholar] [CrossRef]
- Fischer, J. On the fossil record of chimaeroid (Holocephali) egg cases. In GeoBonn Abstract Book; 2018; p. 249. [Google Scholar]
- Brown, R.W. Fossil egg capsules of chimaeroid fishes. J. Paleontol. 1946, 20, 261–266. [Google Scholar]
- Reichenbach-Klinke, H.H.; Frickhinger, K.A. Neuer Fund einer Holocephalen-Eikapsel im Ob. Jura des Solnhofener Schiefers. Fossilien 1986, 6, 256–257. [Google Scholar]
- Duffin, C.J.; Lauer, B.; Lauer, R.; Popov, E.; Ward, D.J. Chimeroid egg capsules from the Late Jurassic lithographic limestones of southern Germany. In Proceedings of the The Society of Vertebrate Paleontology 80th Annual Meeting, Virtual Conference, 12–16 October 2020; pp. 101–102. [Google Scholar]
- Frickhinger, K.A. The Fossils of Solnhofen; Goldschneck-Verlag: Korb, Germany, 1994; p. 333. ISBN 3-926129-15-8. [Google Scholar]
- Resch, U.; Lauer, B. Chimaeropsis paradoxa—Eine wenig bekannte Chimäre aus den Solnhofener Plattenkalken. Der Steinkern 2016, 27, 52–61. [Google Scholar]
- Harrison, G.W.M.; Kirkland, J.I.; Fischer, J.; San Miguel, G.; Wood, J.R.; Santucci, V.L. Two chimaeroid egg case remains from the Late Cretaceous, Mesa Verde National Park, Colorado, USA. Bull. New Mexico Mus. Nat. Hist. Sci. 2021, 82, 113–120. [Google Scholar]
- Maisey, J.G.; Naylor, G.J.P.; Ward, D.J. Mesozoic elasmobranchs, neoselachian phylogeny and the rise of modern elasmobranch diversity. In Mesozoic Fishes—Systematics, Paleoenvironments and Biodiversity; Arratia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; Volume 3, pp. 17–56. ISBN 978-3-89937-053-9. [Google Scholar]
- Maisey, J.G.; Janvier, P.; Pradel, A.; Denton, J.S.S.; Bronson, A.; Miller, R.; Burrow, C.J. Doliodus and pucapampellids: Contrasting perspectives on stem chondrichthyan morphology. In Evolution and Development of Fishes; Johanson, Z., Underwood, C., Richter, M., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 87–109. [Google Scholar] [CrossRef]
- Villalobos-Segura, E.; Marramà, G.; Carnevale, G.; Claeson, K.M.; Underwood, C.J.; Naylor, G.J.P.; Kriwet, J. The Phylogeny of Rays and Skates (Chondrichthyes: Elasmobranchii) Based on Morphological Characters Revisited. Diversity 2022, 14, 456. [Google Scholar] [CrossRef]
- Hay, O.P. Bibliography and catalogue of the fossil Vertebrata of North America. Bull. U.S. Geol. Surv. 1902, 179, 1–868. [Google Scholar] [CrossRef]
- Regan, C.T. A classification of the selachian fishes. Proc. Zool. Soc. Lond. 1906, 1906, 722–758. [Google Scholar]
- Bonaparte, C.L. Selachorum tabula analytica. N. Ann. Sci. Nat. Bologna 1838, 2, 195–214. [Google Scholar]
- Pradel, A.; Tafforeau, P.; Maisey, J.G.; Janvier, P. A new Paleozoic Symmoriiformes (Chondrichthyes) from the Late Carboniferous of Kansas (USA) and cladistic analysis of early chondrichthyans. PLoS ONE 2011, 6, e24938. [Google Scholar] [CrossRef]
- Coates, M.I.; Gess, R.W.; Finarelli, J.A.; Criswell, K.E.; Tietjen, K. A symmoriiform chondrichthyan braincase and the origin of chimaeroid fishes. Nature 2017, 541, 208–211. [Google Scholar] [CrossRef]
- Coates, M.I.; Finarelli, J.A.; Sansom, I.J.; Andreev, P.S.; Criswell, K.E.; Tietjen, K.; Rivers, M.L.; La Riviere, P.J. An early chondrichthyan and the evolutionary assembly of a shark body plan. Proc. Royal Soc. B. 2018, 285, 20172418. [Google Scholar] [CrossRef]
- Frey, L.; Coates, M.I.; Tietjen, K.; Rücklin, M.; Klug, C. A symmoriiform from the Late Devonian of Morocco demonstrates a derived jaw function in ancient chondrichthyans. Commun. Biol. 2020, 3, 681. [Google Scholar] [CrossRef]
- Frey, L.; Coates, M.; Ginter, M.; Hairapetian, V.; Rücklin, M.; Jerjen, I.; Klug, C. The early elasmobranch Phoebodus: Phylogenetic relationships, ecomorphology and a new time-scale for shark evolution. Proc. Royal Soc. B. 2019, 286, 20191336. [Google Scholar] [CrossRef]
- Maisey, J.G. Remarks on the inner ear of elasmobranchs and its interpretation from skeletal labyrinth morphology. J. Morphol. 2001, 250, 236–264. [Google Scholar] [CrossRef]
- Maisey, J.G.; Lane, J.A. Labyrinth morphology and the evolution of low-frequency phonoreception in elasmobranchs. C. R. Palevol. 2010, 9, 289–309. [Google Scholar] [CrossRef]
- Maisey, J.G. Chondrichthyan phylogeny: A look at the evidence. J. Vertebr. Paleontol. 1984, 4, 359–371. [Google Scholar] [CrossRef]
- Pradel, A.; Maisey, J.G.; Tafforeau, P.; Mapes, R.H.; Mallatt, J. A Palaeozoic shark with osteichthyan-like branchial arches. Nature 2014, 509, 608–611. [Google Scholar] [CrossRef]
- Feichtinger, I.; Engelbrecht, A.; Lukeneder, A.; Kriwet, J. New chondrichthyans characterised by cladodont-like tooth morphologies from the Early Cretaceous of Austria, with remarks on the microstructural diversity of enameloid. Hist. Biol. 2020, 32, 823–836. [Google Scholar] [CrossRef]
- Maisey, J.G.; Denton, J.S.S. Dermal denticle patterning in the Cretaceous hybodont shark Tribodus limae (Euselachii, Hybodontiformes), and its implications for the evolution of patterning in the chondrichthyan dermal skeleton. J. Vertebr. Paleontol. 2016, 36, e1179200. [Google Scholar] [CrossRef] [Green Version]
- Maisey, J.G. The interrelationships of phalacanthous selachians. Neues Jahrb. Geol. Paläontol. 1975, 9, 553–567. [Google Scholar]
- Stumpf, S.; Etches, S.; Underwood, C.J.; Kriwet, J. Durnonovariaodus maiseyi gen. et sp. nov., a new hybodontiform shark-like chondrichthyan from the Upper Jurassic Kimmeridge Clay Formation of England. PeerJ 2021, 9, e11362. [Google Scholar] [CrossRef]
- Coates, M.I.; Gess, R.W. A new reconstruction of Onychoselache traquairi, comments on early chondrichthyan pectoral girdles and hybodontiform phylogeny. Palaeontology 2007, 50, 1421–1446. [Google Scholar] [CrossRef]
- Rees, J. Interrelationships of Mesozoic hybodont sharks as indicated by dental morphology-preliminary results. Acta Geol. Pol. 2008, 58, 217–221. [Google Scholar]
- Maisey, J.G. Growth and form of finspines in hybodont sharks. Palaeontology 1978, 21, 657–666. [Google Scholar]
- Maisey, J.G. The anatomy and interrelationships of Mesozoic hybodont sharks. Am. Mus. Novit. 1982, 2724, 1–48. [Google Scholar]
- Rees, J.; Underwood, C.J. The status of the shark genus Lissodus, and the position of nominal Lissodus species within the Hybodontoidea. J. Vertebr. Paleontol. 2002, 22, 471–479. [Google Scholar] [CrossRef]
- Klug, S.; Tütken, T.; Wings, O.; Pfretzschner, H.-U.; Martin, T. A Late Jurassic freshwater shark assemblage (Chondrichthyes, Hybodontiformes) from the southern Junggar Basin, Xinjiang, Northwest China. Palaeobiodivers. Palaeoenviron. 2010, 90, 241–257. [Google Scholar] [CrossRef]
- Fischer, J.; Voigt, S.; Schneider, J.W.; Buchwitz, M.; Voigt, S. A selachian freshwater fauna from the Triassic of Kyrgyzstan and its implication for Mesozoic shark nurseries. J. Vertebr. Paleontol. 2011, 31, 937–953. [Google Scholar] [CrossRef]
- Wen, W.; Kriwet, J.; Zhang, Q.; Benton, M.J.; Duffin, C.J.; Huang, J.; Zhou, C.; Hu, S.; Ma, Z. Hybodontiform shark remains (Chondrichthyes, Elasmobranchii) from the Lower Triassic of Yunnan Province, China, with comments on hybodontiform diversity across the PTB. J. Vertebr. Paleontol. 2022, 42, e2108712. [Google Scholar] [CrossRef]
- Cuny, G.; Ouaja, M.; Srarfi, D.; Schmitz, L.; Buffetaut, E.; Benton, M.J. Fossil sharks from the Early Cretaceous of Tunisia. Rev. De Paléobiologie 2004, 9, 127–142. [Google Scholar]
- Cuny, G.; Suteethorn, V.; Kamha, S.; Buffetaut, E.; Philippe, M. A new hybodont shark assemblage from the Lower Cretaceous of Thailand. Hist. Biol. 2006, 18, 21–31. [Google Scholar] [CrossRef]
- Cuny, G.; Cavin, L.; Suteethorn, V. A new hybodont with a cutting dentition from the Lower Cretaceous of Thailand. Cretac. Res. 2009, 30, 515–520. [Google Scholar] [CrossRef]
- Cuny, G.; Laojumpon, C.; Cheychiw, O.; Lauprasert, K. Fossil vertebrate remains from Kut Island (Gulf of Thailand, Early Cretaceous). Cretac. Res. 2010, 31, 415–423. [Google Scholar] [CrossRef]
- Stumpf, S.; Meng, S.; Kriwet, J. Diversity Patterns of Late Jurassic Chondrichthyans: New Insights from a Historically Collected Hybodontiform Tooth Assemblage from Poland. Diversity 2022, 14, 85. [Google Scholar] [CrossRef]
- Late Jurassic selachians (Chondrichthyes: Hybodontiformes, Neoselachii) from Central Portugal. Neues Jahrb. Geol. Paläontol. Mh. 2004, 4, 233–256. [CrossRef]
- Vullo, R.; Abit, D.; Ballèvre, M.; Billon-Bruyat, J.-P.; Bourgeais, R.; Buffetaut, E.; Daviero-Gomez, V.; Garcia, G.; Gomez, B.; Mazin, J.-M.; et al. Palaeontology of the Purbeck-type (Tithonian, Late Jurassic) bonebeds of Chassiron (Oléron Island, western France). C. R. Palevol 2014, 13, 421–441. [Google Scholar] [CrossRef]
- Owen, R. Lectures on the Comparative Anatomy and Physiology of the Vertebrate Animals, Delivered at the Royal College of Surgeons of England in 1844 and 1846; Part 1. Fishes; Longman: London, UK, 1846. [Google Scholar]
- Agassiz, L.J.R. Recherches sur les Poissons Fossils; Petitpierre: Neuchâtel, Switzerland, 1833–1844; Volumes 1–5. [Google Scholar]
- Brown, C. Ueber das Genus Hybodus und seine systematische Stellung. Palaeontographica 1900, 46, 149–174. [Google Scholar]
- Maisey, J.G. Cranial anatomy of the Lower Jurassic shark Hybodus reticulatus (Chondrichthyes: Elasmobranchii), with comments on hybodontid systematics. Am. Mus. Novit. 1987, 2878, 1–39. [Google Scholar]
- Egerton, P. Description of the Mouth of a Hybodus found by Mr. Boscawen Ibbetson in the Isle of Wight. Quart. J. Geol. Soc. 1844, 1, 414–416. [Google Scholar] [CrossRef] [Green Version]
- Rees, J.; Underwood, C.J. Hybodont sharks of the English Bathonian and Callovian (Middle Jurassic). Palaeontology 2008, 51, 117–147. [Google Scholar] [CrossRef] [Green Version]
- Frickinger, K.A. The Fossils of Solnhofen 2; Goldschneck-Verlag: Korb, Germany, 1999; p. 190. ISBN 3-926129-26-3. [Google Scholar]
- Casier, E. Contributions à l’étude des poissons fossiles de la Belgique. XII—Sélaciens et Holocéphales sinémuriens de la Province de Luxembourg. Bull. Inst. R. Sci. Nat. Belg. 1959, 38, 1–35. [Google Scholar]
- Woodward, A.S. On some Remains of the Extinct Selachian Asteracanthus from the Oxford Clay of Peterborough, preserved in the Collection of Alfred, N. Leeds, Esq., of Eyebury. Ann. Mag. Nat. Hist. 1888, 2, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Woodward, A.S. Catalogue of the Fossil Fishes in the British Museum (Natural History); Part I.; British Museum: London, UK, 1889; p. 474. [Google Scholar]
- Martill, D.M. Fossils of the Oxford Clay. In Field Guide to Fossils 4; Martill, D.M., Hudson, J.D., Eds.; Palaeontological Association: London, UK, 1991; pp. 197–225. [Google Scholar]
- Szabó, M.; Főzy, I. Asteracanthus (Hybodontiformes: Acrodontidae) remains from the Jurassic of Hungary, with the description of a new species and with remarks on the taxonomy and paleobiology of the genus. Neues Jahrb. Geol. Paläontol. Abh. 2020, 297, 295–309. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, S. A small assemblage of marine hybodont sharks from the Bathonian of the Jaisalmer Basin, India. Neues Jahrb. Geol. Paläontol. Abh. 2021, 301, 317–333. [Google Scholar] [CrossRef]
- Carrillo-Briceño, J.D.; Cadena, E.A. A new hybodontiform shark (Strophodus Agassiz 1838) from the Lower Cretaceous (Valanginian-Hauterivian) of Colombia. PeerJ 2022, 10, e13496. [Google Scholar] [CrossRef]
- Pfeil, F.H. Ein neues Asteracanthus-Gebiss aus den Kieselplattenkalken (Oberjura, Tithonium, Malm Zeta 3, Mörnsheim-Formation) des Besuchersteinbruchs in Mühlheim. Fr. Bayer. Staatssamml. Paläontol. Hist. Geol. München 2011, 39, 36–60. [Google Scholar]
- Peyer, B. Die schweizerischen Funde von Asteracanthus (Strophodus). Schweiz. Palaeontol. Abh. 1946, 64, 1–101. [Google Scholar]
- Ebert, D.A.; Dando, M.; Fowler, S. Sharks of the World: A Complete Guide; Princeton University Press: Princeton, NJ, USA, 2021; p. 624. [Google Scholar]
- Anderson, T.F.; Popp, B.N.; Williams, A.C.; Ho, L.Z.; Hudson, J.D. The stable isotopic records of fossils from the Peterborough Member, Oxford Clay Formation (Jurassic), UK: Palaeoenvironmental implications. J. Geol. Soc. 1994, 151, 125–138. [Google Scholar] [CrossRef]
- Dromart, G.; Garcia, J.P.; Picard, S.; Atrops, F.; Lécuyer, C.; Sheppard, S.M.F. Ice age at the Middle–Late Jurassic transition? Earth Planet Sci. Lett. 2003, 213, 205–220. [Google Scholar] [CrossRef]
- Lécuyer, C.; Picard, S.; Garcia, J.P.; Sheppard, S.M.F.; Grandjean, P.; Dromart, G. Thermal evolution of Tethyan surface waters during the Middle-Late Jurassic: Evidence from D18O values of marine fish teeth. Paleoceanogr. Paleoclimatol. 2003, 18, 1–16. [Google Scholar] [CrossRef]
- Maisey, J.G. Finspine morphogenesis in squalid and heterodontid sharks. Zool. J. Linn. Soc. 1979, 66, 161–183. [Google Scholar] [CrossRef]
- da Silva, J.P.C.; de Carvalho, M.R. Morphology and phylogenetic significance of the pectoral articular region in elasmobranchs (Chondrichthyes). Zool. J. Linn. Soc. 2015, 175, 525–568. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, B. The xenacanth shark neurocranium, with comments on elasmobranch monophyly. Bull. Am. Mus. 1981, 169, 3–66. [Google Scholar]
- Naylor, G.J.P.; Caira, J.N.; Jensen, K.; Rosana, K.A.M.; Straube, N.; Lakner, C. Elasmobranch phylogeny: A mitochondrial estimate based on 595 species. In Biology of Sharks and Their Relatives, 2nd ed.; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 31–56. ISBN 9780429106545. [Google Scholar]
- Li, C.; Matthes-Rosana, K.A.; Garcia, M.; Naylor, G.J.P. Phylogenetics of Chondrichthyes and the problem of rooting phylogenies with distant outgroups. Mol. Phylogenetics Evol. 2012, 63, 365–373. [Google Scholar] [CrossRef]
- Li, J.; Sun, Z.; Cuny, G.; Ji, C.; Jiang, D.; Zhou, M. An unusual shark assemblage from the Ladinian–Carnian interval of South China. Pap. Palaeotol. 2021, 8, e1404. [Google Scholar] [CrossRef]
- Ivanov, A.O.; Duffin, C.J.; Naugolnykh, S.V. A new euselachian shark from the early Permian of the Middle Urals, Russia. Acta Palaeontol. Pol. 2017, 62, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Guinot, G.; Underwood, C.J.; Cappetta, H.; Ward, D.J. Sharks (Elasmobranchii: Euselachii) from the Late Cretaceous of France and the UK. J. Syst. Palaeontol. 2013, 11, 589–671. [Google Scholar] [CrossRef]
- Duffin, C.J.; Ward, D.J. The Early Jurassic palaeospinacid sharks of Lyme Regis, southern England. Belg. Geol. Surv. Prof. Pap. 1993, 264, 53–102. [Google Scholar]
- Glikman, L.S. On the relationships between the families Lamnidae and Odontaspidae and on new lamnid genera from the Late Cretaceous. Tr. Geol. Muz. AP Karpinskogo Akad. Nauk. SSSR 1957, 1, 110–117. (In Russian) [Google Scholar]
- Thies, D.; Vespermann, J.; Solcher, J. Two new neoselachian sharks (Elasmobranchii, Neoselachii, Synechodontiformes) from the Rhaetian (Late Triassic) of Europe. Palaeontogr. A 2014, 303, 137–167. [Google Scholar] [CrossRef]
- Guinot, G.; Cappetta, H.; Adnet, S. A rare elasmobranch assemblage from the Valanginian (Lower Cretaceous) of southern France. Cretac. Res. 2014, 48, 54–84. [Google Scholar] [CrossRef]
- Stumpf, S.; Kriwet, J. A new Pliensbachian elasmobranch (Vertebrata, Chondrichthyes) assemblage from Europe, and its contribution to the understanding of late Early Jurassic elasmobranch diversity and distributional patterns. PalZ 2019, 93, 637–658. [Google Scholar] [CrossRef] [Green Version]
- Underwood, C.J. Sharks and rays. In Fossils of the Kimmeridge Clay Formation; Martill, D.M., Etches, S., Eds.; Palaeontological Association: London, UK, 2020; Volume 2, pp. 14–32. ISBN 978-0-901702-75-3. [Google Scholar]
- Klug, S. Monophyly, phylogeny and systematic position of the †Synechodontiformes (Chondrichthyes, Neoselachii). Zool. Scr. 2010, 39, 37–49. [Google Scholar] [CrossRef]
- Maisey, J.G. Cranial Morphology of the Fossil Elasmobranch Synechodus dubrisiensis. Am. Mus. Novit. 1985, 2804, 1–28. [Google Scholar]
- Cuny, G.; Rieppel, O.; Sander, P.M. The shark fauna from the Middle Triassic (Anisian) of North-Western Nevada. Zool. J. Linn. Soc. 2001, 133, 285–301. [Google Scholar] [CrossRef]
- Cuny, G.; Tabouelle, J. First mention of the family Pseudonotidanidae (Chondrichthyes: Neoselachii) in the Jurassic of Normandy. Bull. Sci. Géol. Normandes 2014, 7, 21–28. [Google Scholar]
- Musper, F. Der Brenztaloolith, sein Fossilgehalt und seine Deutung. Württemb. Jahrb. 1920, 76, 1–35. [Google Scholar]
- Batchelor, T.J.; Duffin, C.J. First description of sharks’ teeth from the Ferruginous Sands Formation (Aptian, Early Cretaceous) of the Isle of Wight. Proc. Geol. Assoc. 2020, 131, 353–359. [Google Scholar] [CrossRef]
- Klug, S.; Kriwet, J. A new basal galeomorph shark (Synechodontiformes, Neoselachii) from the Early Jurassic of Europe. Naturwissenschaften 2008, 95, 443–448. [Google Scholar] [CrossRef]
- Duffin, C.J. New records of Late Jurassic Sharks Teeth from Southern Germany. Stutt. Beitr. Naturkd. B (Geol. Paläontologie) 1993, 193, 1–13. [Google Scholar]
- Hasse, C. Das Natürliche System der Elasmobranchier auf Grundlage des Baues und der Entwicklung ihrer Wirbelsäule; Besonderer Theil, Gustav Fischer: Jena, Germany, 1882; pp. 1–285. [Google Scholar]
- Rafinesque, C.S. Indice d’ittiologia siciliana ossia catalogo metodico dei nomi latini, italiani, e siciliani, e siciliani dei pesci, che si rinvengono in Sicilia disposti secondo un metodo naturale eseguito da un appendice che contiene la descrizione di alcuni nuovi pesci siciliani. In Ill Opuscolo del Signore C.S. Rafinesque Schmaltz. Giovanni del Nobolo, Messina. 1810, pp. 1–70. Available online: https://www.biodiversitylibrary.org/bibliography/58965 (accessed on 25 February 2023).
- Fowler, H.W. New Taxonomic names of fish-like vertebrates. Not. Nat. Acad. Nat. Sci. Philadelphia 1947, 187, 1–16. [Google Scholar]
- Berg, L.S. A classification of fish-like vertebrates. Proc. USSR Acad. Sci. 1937, 4, 1277–1280. [Google Scholar]
- de Blainville, H.M.D. Prodrome d’une nouvelle distribution systématique du règne animal. Bull. Sci. Soc. Philom. Paris 1816, 8, 105–124. [Google Scholar]
- Thies, D. Jurazeitliche Neoselachier aus Deutschland und S-England. Cour. Forsch. Inst. Senckenberg 1982, 58, 1–116. [Google Scholar]
- Plieninger, T. The remains of vertebrate animals in the coral limestone of Schnaitheim. Jahresh. Ver. vaterl. Naturkd. Württemb. 1847, 3, 226–227. [Google Scholar]
- Eastman, C.R. New elasmobranchs from Solenhofen in the Carnegie Museum. Am. J. Sci. 1911, 31, 399–404. [Google Scholar] [CrossRef]
- Underwood, C.J.; Ward, D.J. Neoselachian sharks and rays from the British Bathonian (Middle Jurassic). Palaeontology 2004, 47, 447–501. [Google Scholar] [CrossRef]
- Slater, T.S. Sharks with question marks: Impacts of a new fossil on interrelationships of early bullhead sharks. Pal. Ass. Newsl. 2016, 93, 68–72. [Google Scholar]
- Gray, J.E. List of the Specimens of Fish in the Collection of the British Museum; Part, I. Chondropterygii; British Museum (Natural History): London, UK, 1851; p. 160. [Google Scholar]
- Koken, E. Pisces. 3–142. In Grundzüge der Paläontologie; Zittel, K.A., Ed.; Oldenbourg: München/Berlin, Germany, 1911; Volume 2, p. 142. [Google Scholar]
- Maia, A.M.; Wilga, C.A.; Lauder, G.V. Biomechanics of locomotion in sharks, rays, and chimaeras. In Biology of Sharks and Their Relatives, 2nd ed.; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 125–148. ISBN 9780429106545. [Google Scholar]
- Maisey, J.G. Fossil hornshark finspines (Elasmobranchii; Heterodontidae) with notes on a new species (Heterodontus tuberculatus). Neues Jahrb. Geol. Paläontol. Abh. 1982, 164, 393–413. [Google Scholar] [CrossRef]
- Hovestadt, D.C. Reassessment and Revision of the fossil Heterodontidae (Chondrichthyes: Neoselachii) based on tooth morphology of extant taxa. Palaeontos 2018, 30, 3–120. [Google Scholar]
- Applegate, S.P. A revision of the higher taxa of Orectoloboids. J. Mar. Biol. Assoc. India 1974, 14, 743–751. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Genera/Species by Family/Subfamily. 2022. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 26 April 2022).
- Thiollière, V. Troisième notice sur les gisements à poissons fossiles situés dans le Jura du département de l’Ain. Annls Sci. Phys. Nat. Lyon 1852, 4, 353–446. [Google Scholar]
- Cappetta, H. Mesozoic and Cenozoic Elasmobranchii. In Handbook of Palaeoichthyology, Chondrichthyes II.; Schultze, H.-P., Ed.; Verlag Dr. Fischer: Stuttgart, Germany, New York, NY, USA, 1987; Volume 3B, p. 193. [Google Scholar]
- White, E.G. A classification and phylogeny of the elasmobranch fishes. Am. Mus. Novit. 1936, 837, 1–16. [Google Scholar]
- Woodward, A.S. On two new elasmobranch fishes (Crossorhinus jurassicus sp. nov. and Protospinax annectans gen. et sp. nov.) from the Upper Jurassic lithographic stone of Bavaria. In Proceedings of the Zoological Society of London; Blackwell Publishing Ltd.: Oxford, UK, 1918; pp. 231–235. [Google Scholar]
- Pollerspöck, J.; Straube, N. Bibliography Database of Living/Fossil Sharks, Rays and Chimaeras (Chondrichthyes: Elasmobranchii, Holocephali)—List of Valid Extant Species—List of Described Extant Species—Statistic. World Wide Web Electronic Publication, 2021, Version 03/2021, ISSN: 2195-6499. Available online: www.shark-references.com (accessed on 12 September 2022).
- Underwood, C.J.; Ward, D.J. A review of the Mesozoic Record of the Carcharhiniformes. In Mesozoic Fishes—Homology and Phylogeny; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2008; Volume 4, pp. 433–442. ISBN 978-3-89937-080-5. [Google Scholar]
- Brée, B.; Condamine, F.L.; Guinot, G. Combining palaeontological and neontological data shows a delayed diversification burst of carcharhiniform sharks likely mediated by environmental change. Sci. Rep. 2022, 12, 21906. [Google Scholar] [CrossRef]
- Marck, W.V.D. Fossile Fische, Krebse und Pflanzen aus dem Plattenkalk der jüngsten Kreide in Westphalen. Palaeontographica (1846–1933) 1863, 1–40. [Google Scholar]
- Vélez-Zuazo, X.; Agnarsson, I. Shark Tales: A molecular species-level phylogeny of sharks (Selachimorpha, Chondrichthyes). Mol. Phylogenet. Evol. 2011, 58, 207–217. [Google Scholar] [CrossRef]
- Soares, K.D.; de Carvalho, M.R. Phylogenetic relationship of catshark species of the genus Scyliorhinus (Chondrichthyes, Carcharhiniformes, Scyliorhinidae) based on comparative morphology. Zoosyst. Evol. 2020, 96, 345–395. [Google Scholar] [CrossRef]
- Gill, T. Analytical synopsis of the Order of Squali and revision of the nomenclature of the genera. Ann. Lyceum Nat. Hist. New York 1862, 7, 367–408. [Google Scholar] [CrossRef]
- Thies, D.; Mudroch, A.; Turner, S. The potential of vertebrate microfossils for marine to non-marine correlation in the Late Jurassic. Prog. Nat. Sci. 2007, 17, 655–663. [Google Scholar] [CrossRef]
- Underwood, C.J.; Mitchell, S. Albian and Cenomanian (Cretaceous) Selachian Faunas from North East England. Spec. Pap. Palaeontol. 1999, 60, 9–59. [Google Scholar]
- Thies, D.; Candoni, L. Corysodon Saint-Seine 1949—A valid genus of Mesozoic neoselachian sharks. Geologica et Palaeontologica 1998, 32, 221–233. [Google Scholar]
- Leidner, A.; Thies, D. Placoid scales and oral teeth of Late Jurassic elasmobranchs from Europe. In Mesozoic Fishes—Systematics and Fossil Record; Arratia, G., Schultze, H.-P., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 1999; Volume 2, pp. 29–40. ISBN 3-931516-48-2. [Google Scholar]
- Candoni, L. Découverte de Parasymbolus octevillensis gen. et sp. nov. (Scyliorhinidae–Elasmobranchii) dans le Kimméridgien de Normandie, France. Belg. Geol. Surv. Prof. Pap. 1993, 264, 147–156. [Google Scholar]
- Candoni, L. Parasymbolus octevillensis Candoni 1993 (Scyliorhinidae-Elasmobranchii) du Kimmeridgien havrais. Bull. Soc. Géol. 1994, 81, 47–53. [Google Scholar]
- Underwood, C.J. Sharks, rays and a chimaeroid from the Kimmeridgian (Late Jurassic) of Ringstead, southern England. Palaeontology 2002, 45, 297–325. [Google Scholar] [CrossRef] [Green Version]
- Kriwet, J. Late Jurassic Elasmobranch and Actinopterygian fishes from Portugal and Spain. J. Iber. Geol. 1998, 24, 241–260. [Google Scholar]
- Klug, S.; Kriwet, J. An offshore fish assemblage (Elasmobranchii, Actinopterygii) from the Late Jurassic of NE Spain. Paläontol. Z. 2013, 87, 235–257. [Google Scholar] [CrossRef]
- Rees, J. Neoselachian shark and ray teeth from the Valanginian, Lower Cretaceous, of Wąwał, Central Poland. Palaeontology 2005, 48, 209–221. [Google Scholar] [CrossRef]
- Sweetman, S.C.; Underwood, C.J. A neoselachian shark from the non-marine Wessex Formation (Wealden Group: Early Cretaceous, Barremian) of the Isle of Wight, southern England. Palaeontology 2006, 49, 457–465. [Google Scholar] [CrossRef]
- Kiel, S.; Peckmann, J.; Simon, K. Catshark egg capsules from a Late Eocene deep-water methane-seep deposit in western Washington State, USA. Acta Palaeontol. Pol. 2013, 58, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Berg, L.S. System der Rezenten und Fossilen Fischartigen und Fische; VEB Verlag der Wissenschaften: Berlin, Germany, 1958; p. 310. [Google Scholar]
- Kriwet, J.; Klug, S.; Canudo, J.I.; Cuenca-Bescos, G. A new Early Cretaceous lamniform shark (Chondrichthyes, Neoselachii). Zool. J. Linn. Soc. 2008, 154, 278–290. [Google Scholar] [CrossRef]
- Carrillo-Briceño, J.D.; Parra, J.D.; Luque, J. A new lamniform shark Protolamna ricaurtei sp. nov. from the Lower Cretaceous of Colombia. Cretac. Res. 2019, 95, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Jambura, P.L.; Türtscher, J.; Kindlimann, R.; Metscher, B.; Pfaff, C.; Stumpf, S.; Weber, G.W.; Kriwet, J. Evolutionary trajectories of tooth histology patterns in modern sharks (Chondrichthyes, Elasmobranchii). J. Anat. 2020, 236, 753–771. [Google Scholar] [CrossRef]
- Applegate, S.P. The origin of the Lamniform Sharks—A Study in Morphology and Paleontology of Recent and fossil genera. In Proceedings of the American Elasmobranch Society, 17th Annual Meeting, State College, PA, USA, 6–10 July 2001. [Google Scholar]
- Landemaine, O.; Thies, D.; Waschkewitz, J. The Late Jurassic shark Palaeocarcharias (Elasmobranchii, Selachimorpha)—Functional Morphology of Teeth, Dermal Cephalic Lobes and Phylogenetic Position. Palaeontogr. Abt. A 2018, 312, 103–165. [Google Scholar] [CrossRef]
- Wills, S.; Bernard, E.L.; Brewer, P.; Underwood, C.J.; Ward, D.J. Palaeontology, stratigraphy and sedimentology of Woodeaton Quarry (Oxfordshire) and a new microvertebrate site from the White Limestone Formation (Bathonian, Jurassic). Proc. Geol. Assoc. 2019, 130, 170–186. [Google Scholar] [CrossRef] [Green Version]
- Underwood, C.J.; Ward, D.J. Environmental distribution of Bathonian (Middle Jurassic) neoselachians in Southern England. In Mesozoic fishes—Systematics, Paleoenvironments and Biodiversity; Arratia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; Volume 3, pp. 111–122. ISBN 978-3-89937-053-9. [Google Scholar]
- Košťák, M.; Vaňková, L.; Mazuch, M.; Bubík, M.; Reháková, D. Cephalopods, small vertebrate fauna and stable isotope (δ13C, δ18O) record from the Jurassic-Cretaceous transition (uppermost Crassicollaria through Calpionella Zones) of the Outer Western Carpathians, Kurovice Quarry (Czechia). Cretac. Res. 2018, 92, 43–65. [Google Scholar] [CrossRef]
- Jambura, P.L.; Kindlimann, R.; López-Romero, F.; Marramà, G.; Pfaff, C.; Stumpf, S.; Türtscher, J.; Underwood, C.J.; Ward, D.J.; Kriwet, J. Micro-computed tomography imaging reveals the development of a unique tooth mineralization pattern in mackerel sharks (Chondrichthyes; Lamniformes) in deep time. Sci. Rep. 2019, 9, 9652. [Google Scholar] [CrossRef] [Green Version]
- de Buen, F. Catálogo ictiológico del Mediterráneo español y de Marruecos: Recopilando lo publicado sobre peces de las costas mediterránea y próximas del Atlántico (Mar de España). Resultados de las ampafias Realizadas por Acuerdos Internacionales. Inst. Español De Oceanogr. 1926, 2, 1–221. [Google Scholar]
- Maisey, J.G. The postorbital palatoquadrate articulation in elasmobranchs. J. Morphol. 2008, 269, 1022–1040. [Google Scholar] [CrossRef]
- Paillard, A.; Shimada, K.; Pimiento, C. The fossil record of extant elasmobranchs. J. Fish Biol. 2021, 98, 445–455. [Google Scholar] [CrossRef]
- Rafinesque, C.S. Caratteri di Alcuni Nuovi Generi e Nuove Specie di Animali e Pinate della Sicilia, con Varie Osservazioni Sopra i Medisimi, lère Partie. 1810, 106p. Available online: https://www.biodiversitylibrary.org/bibliography/104418 (accessed on 25 February 2023).
- Ayres, W.O. Descriptions of new species of Californian fishes. Proc. Calif. Acad. Sci. (Series 1) 1855, 1, 23–77. [Google Scholar]
- Kriwet, J.; Klug, S. A new Jurassic cow shark (Chondrichthyes, Hexanchiformes) with comments on Jurassic hexanchiform systematics. Swiss J. Geosci. 2011, 104, 107–114. [Google Scholar] [CrossRef]
- Maisey, J.G. The Upper Jurassic hexanchoid elasmobranch Notidanoides n. g. Neues Jahrb. Geol. Paläontol. Abh. 1986, 172, 83–106. [Google Scholar] [CrossRef]
- Cappetta, H. Hexanchiforme nouveau (Neoselachii) du Crétacé inférieur du Sud de la France. Palaeovertebrata 1990, 20, 33–54. [Google Scholar]
- Beyrich, H.E.; Freiherr von Buch, C.L.; von Carnall, C.L.; Ewald, J.; Guggenheim, M.; von Strombeck, A.; Zerrenner, A. Reise nach Kelheim, Ingolstadt, Eichstätt, Solnhofen und Pappenheim. ZDGG 1849, 1, 423–447. [Google Scholar]
- Pfeil, F.H. Tooth morphological studies on extant and fossil sharks of the order Chlamydoselachiformes and Echinorhiniformes. Palaeo Ichthyologica 1983, 1, 1–315. (In German) [Google Scholar]
- Münster, G.G. Description of some new fish from the Jurassic Formation of Bavaria and Württemberg. Contrib. Petrefactol. 1843, 6, 53–56. [Google Scholar]
- Ward, D.J.; Thies, D. Hexanchid shark teeth (Neoselachii, Vertebrata) from the Lower Cretaceous of Germany and England. Mes. Res. 1987, 1, 89–106. [Google Scholar]
- Maisey, J.G.; Wolfram, K.E. Notidanus. In Living Fossils; Eldredge, N., Stanley, S.M., Eds.; Springer: New York, NY, USA, 1984; pp. 170–180. [Google Scholar]
- Cuvier, G. Le Règne Animal Distribué D’après son Organisation Pour Servir de Base à L’histoire Naturelle des Animaux et D’introduction à L’anatomie Comparée. Les Reptiles, les Poissons, les Mollusques et les Annélides; Deterville: Paris, France, 1817; p. 532. [Google Scholar]
- Jaekel, O. Über Hybodus Ag. Sber. Ges. Naturf. Freunde Berlin 1989, 89, 135–146. [Google Scholar]
- von Meyer, H. Letter on Thaumas and Acrosaurus. In New Yearbook of Mineralogy, Geognosy, Geology and Petrefacts. 1856, pp. 418–420. Available online: https://shark-references.com/species/view/Squatina-speciosa (accessed on 25 February 2023).
- Duméril, C. Zoologie Analytique, ou Méthode Naturelle de Classification des Animaux, Rendue Plus Facile à l’aide de Tableaux Synoptiques; Librairie Allais: Paris, France, 1806; pp. 331–344. [Google Scholar]
- Cappetta, H. Elasmobranchii Post-Triadici. In Fossilium Catalogus, I: Animalia; Riegraf, W., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2006; p. 472. [Google Scholar]
- Maisey, J.G.; Ehret, D.J.; Denton, J.S. A new genus of Late Cretaceous angel shark (Elasmobranchii; Squatinidae), with comments on squatinid phylogeny. Am. Mus. Novit. 2020, 2020, 1–29. [Google Scholar] [CrossRef]
- Mudroch, A. Fischzähne aus dem Oberjura Nordwesteuropas: Systematik, Biogeochemie und Palökologie. Ph.D. Thesis, University of Hannover, Hannover, Germany, 2001. [Google Scholar]
- Delsate, D. Une nouvelle faune de poissons et requins toarciens du sud du Luxembourg (Dudelange) et de l’Allemagne (Schömberg). Bull. Acad. Lorraine Sci. 2003, 42, 13–49. [Google Scholar]
- Kriwet, J. Neoselachian remains [Chondrichthyes, Elasmobranchii] from the Middle Jurassic of SW Germany and NW Poland. Acta Palaeontol. Pol. 2003, 48, 583–594. [Google Scholar]
- Rees, J. Neoselachian sharks from the Callovian–Oxfordian (Jurassic) of Ogrodzieniec, Zawiercie Region, southern Poland. Palaeontology 2010, 53, 887–902. [Google Scholar] [CrossRef]
- Rees, J. Palaeoecological Implications of Neoselachian Shark Teeth from the Bathonian (Middle Jurassic) Ore-Bearing Clays at Gnaszyn, Kraków-Silesia Homocline, Poland. Acta Geol. Pol. 2012, 62, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Thies, D. Some Problematical Sharks Teeth (Chondrichthyes, Neoselachii) from the Early and Middle Jurassic of Germany. PalZ. 1989, 63, 103–117. [Google Scholar] [CrossRef]
- Müller, A.; Diedrich, C.G. Selachier (Pisces, Chondrichthyes) aus dem Cenomanium von Ascheloh am Teutoburger Wald (Nordrhein-Westfalen, NW-Deutschland). Geol. Und Palaeontol. Westfal. 1991, 20, 3–105. [Google Scholar]
- Underwood, C.J. Diversification of the Neoselachii (Chondrichthyes) during the Jurassic and Cretaceous. Paleobiology 2006, 32, 215. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.J. Sharks and Rays. In Field Guide to Fossils: Number 12—Fossils of the Gault Clay; Young, J.R., Gale, A.S., Knight, R.I., Smith, A.B., Eds.; Palaeontological Association: London, UK, 2010; pp. 275–299. ISBN 978-1-4443-3542-2. [Google Scholar]
- Underwood, C.J.; Mitchell, S.F.; Veltcamp, K.J. Shark and ray teeth from the Hauterivian (Lower Cretaceous) of north-east England. Palaeontology 1999, 42, 287–302. [Google Scholar] [CrossRef] [Green Version]
- Maisey, J.G. The Jurassic Selachian Fish Protospinax Woodward. Palaeontology 1976, 19, 733–747. [Google Scholar]
- De Carvalho, M.R.; Maisey, J.G. Phylogenetic relationships of the Late Jurassic shark Protospinax Woodward 1919 (Chondrichthyes: Elasmobranchii). In Mesozoic Fishes—Systematics and Paleoecology; Arratia, G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 1996; Volume 1, pp. 9–46. ISBN 978-3-923871-90-2. [Google Scholar]
- White, E.G. Interrelationships of the Elasmobranchs with a Key to the Order Galea. Bull. Am. Mus. Nat. Hist. 1937, 74, 25–138. Available online: https://hdl.handle.net/2246/369 (accessed on 25 February 2023).
- Schaeffer, B. Comments on Elasmobranch Evolution. In Sharks, skates and rays; Gilbert, P.W., Mathewson, R.F., Rall, D.P., Eds.; John Hopkins Press: Baltimore, MD, USA, 1967; pp. 3–35. ISBN 978-0-8018-0217-1. [Google Scholar]
- Shirai, S. Phylogenetic relationships of the angel sharks, with comments on elasmobranch phylogeny (Chondrichthyes, Squatinidae). Copeia 1992, 1992, 505–518. [Google Scholar] [CrossRef]
- Douady, C.J.; Dosay, M.; Shivji, M.S.; Stanhope, M.J. Molecular phylogenetic evidence refuting the hypothesis of Batoidea (rays and skates) as derived sharks. Mol. Phylogenet. Evol. 2003, 26, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Kousteni, V.; Mazzoleni, S.; Vasileiadou, K.; Rovatsos, M. Complete mitochondrial DNA genome of nine species of sharks and rays and their phylogenetic placement among modern elasmobranchs. Genes 2021, 12, 324. [Google Scholar] [CrossRef] [PubMed]
- Flammensbeck, C.K.; Pollerspöck, J.; Schedel, F.D.B.; Matzke, N.J.; Straube, N. Of teeth and trees: A fossil tip-dating approach to infer divergence times of extinct and extant squaliform sharks. Zool. Scr. 2018, 47, 539–557. [Google Scholar] [CrossRef]
- Maisey, J.G. An evaluation of jaw suspension in sharks. Am. Mus. Novit. 1980, 2706, 1–17. [Google Scholar]
- Jambura, P.L.; Villalobos-Segura, E.; Türtscher, J.; Begat, A.; Staggl, M.A.; Stumpf, S.; Kindlimann, R.; Klug, S.; Lacombat, F.; Pohl, B.; et al. Systematics and Phylogenetic Interrelationships of the Enigmatic Late Jurassic Shark Protospinax annectans Woodward, 1918 with Comments on the Shark–Ray Sister Group Relationship. Diversity 2023, 15, 311. [Google Scholar] [CrossRef]
- Stein, R.W.; Mull, C.G.; Kuhn, T.S.; Aschliman, N.C.; Davidson, L.N.K.; Joy, J.B.; Smith, G.J.; Dulvy, N.K.; Mooers, A.O. Global priorities for conserving the evolutionary history of sharks, rays and chimaeras. Nat. Ecol. Evol. 2018, 2, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Cappetta, H. Les Sélaciens du Crétacé supérieur du Liban. II. Batoïdes. Palaeontogr. Abt. A 1980, 168, 149–229. [Google Scholar]
- de Carvalho, M.R. Higher-level elasmobranch phylogeny, basal squaleans, and paraphyly. In Interrelationships of Fishes; Stiassny, M.L.J., Parenti, L.R., Johnson, G.D., Eds.; Atlantic Press: New York, NY, USA, 1996; pp. 593–660. [Google Scholar]
- Dunn, K.A.; Morrissey, J.F. Molecular phylogeny of elasmobranchs. Copeia 1995, 1995, 526–531. [Google Scholar] [CrossRef]
- Schwartz, F.J.; Maddock, M.B. Cytogenetics of the elasmobranchs: Genome evolution and phylogenetic implications. Mar. Freshw. Res. 2002, 53, 491–502. [Google Scholar] [CrossRef]
- Winchell, C.J.; Martin, A.P.; Mallatt, J. Phylogeny of elasmobranchs based on LSU and SSU ribosomal RNA genes. Mol. Phylogenet. Evol. 2004, 31, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Aschliman, N.C.; Claeson, K.M.; McEachran, J.D. Phylogeny of Batoidea. In Biology of Sharks and Their Relatives, 2nd ed.; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 57–95. ISBN 9780429106545. [Google Scholar]
- Last, P.R.; White, W.; de Carvalho, M.R.; Seret, B.; Stehmann, M.; Naylor, G.J.P. Rays of the World; CSIRO Publishing: Clayton, Australia, 2016; p. 790. ISBN 978-1-5017-0532-8. [Google Scholar]
- Guinot, G.; Adnet, S.; Cappetta, H. An analytical approach for estimating fossil record and diversification events in sharks, skates and rays. PLoS ONE 2012, 7, e44632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvage, H.E. Notice sur un spathobate du terrain portlandien de Boulogne-sur-Mer. Bull. Soc. Acad. Boulogne-Sur-Mer. 1873, 2, 94–103. [Google Scholar]
- Cavin, L.; Cappetta, H.; Séret, B. Révision de Belemnobatis morinicus (Sauvage, 1873) du Portlandien du Boulonnais (Pas-de-Calais, France). Comparaison avec quelques Rhinobatidae Jurassiques. Geol. Et Palaeontol. 1995, 29, 245–267. [Google Scholar]
- Underwood, C.J.; Claeson, K.M. The late Jurassic ray Kimmerobatis etchesi gen. et sp. nov. and the Jurassic radiation of the Batoidea. Proc. Geol. Assoc. 2017, 130, 345–354. [Google Scholar] [CrossRef]
- Cione, A.L. First report of a Jurassic ray outside of Europe. In Systematics and Fossil Record; Arratia, G., Schultze, H.-P., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 1999; Volume 2, pp. 21–28. [Google Scholar]
- Underwood, C.J.; Rees, J. Selachian faunas from the earliest Cretaceous Purbeck Group of Dorset, southern England. Spec. Pap. Palaeontol. 2002, 68, 83–102. [Google Scholar]
- Biddle, J.P. Les Elasmobranches de l’Albien inférieur et moyen (Crétacé inférieur) de la Marne et de la Haute-Marne (France). Belg. Geol. Surv. Prof. Pap. 1993, 264, 191–240. [Google Scholar]
- Kriwet, J. Neoselachier (Pisces, Elasmobranchii) aus der Unterkreide (unteres Barremium) von Galve und Alcaine (Spanien, Provinz Teruel). Palaeo Ichthyologica 1999, 9, 113–142. [Google Scholar]
- Kriwet, J.; Nunn, E.V.; Klug, S. Neoselachians (Chondrichthyes, Elasmobranchii) from the Lower and lower Upper Cretaceous of north-eastern Spain. Zool. J. Linn. Soc. 2009, 155, 316–347. [Google Scholar] [CrossRef] [Green Version]
- Delsate, D.; Candoni, L. Les premières raies (Chondrichthyes-Elasmobranchii) au Jurassique Inférieur en Lorraine (Luxembourg, Belgique, France) et dans le Bassin de Paris. Bull. Acad. Lorraine Sci. 2002, 41, 93–103. [Google Scholar]
- Delsate, D.; Candoni, L. Description de nouveaux morphotypes dentaires de Batomorphii toarciens (Jurassique inférieur) du Bassin de Paris: Archaeobatidae nov. fam. Bull. Soc. Nat. Luxemb. 2001, 102, 131–144. [Google Scholar]
- Biddle, J.P.; Landemaine, O. Contribution à l’étude des Sélaciens du Crétacé du Bassin de Paris. Découverte de quelques nouvelles espèces associées à une faune de type wealdien dans le Barrémien supérieur (Crétacé inférieur) des environs de Troyes (Aube). Musée de Saint–Dizier Saint–Dizier Cahiers 1988, 2, 1–22. [Google Scholar]
- Fischer, J. On the fossil record of skate (Batoidea) egg cases. In Proceedings of the 88. Jahrestagung der Paläontologischen Gesellschaft, Münster, Germany, 26–30 March 2017; Volume 109, p. 39. [Google Scholar]
- Steininger, F. Zur Kenntnis fossiler Euselachier-Eikapseln aus dem Ober-Oligozän von Mitteleuropa. Mitt. Bayer. Staatssamml. Paläontol. Hist. Geol. 1966, 6, 37–49. [Google Scholar]
- Tittensor, D.P.; Mora, C.; Jetz, W.; Lotze, H.K.; Ricard, D.; Berghe, E.V.; Worm, B. Global patterns and predictors of marine biodiversity across taxa. Nature 2010, 466, 1098–1101. [Google Scholar] [CrossRef]
- Rocha, L.A.; Rocha, C.R.; Robertson, D.R.; Bowen, B.W. Comparative phylogeography of Atlantic reef fishes indicates both origin and accumulation of diversity in the Caribbean. BMC Evol. Biol. 2008, 8, 157. [Google Scholar] [CrossRef] [Green Version]
- Schultze, H.-P.; Arratia, G. Knochenfische im weiteren Sinne (Osteichthyes oder Osteognathostomata). In Solnhofen—Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2015; pp. 360–409. ISBN 978-3-89937-195-6. [Google Scholar]
- Roden, V.J.; Hausmann, I.M.; Nützel, A.; Seuss, B.; Reich, M.; Urlichs, M.; Hagdorn, H.; Kiessling, W. Fossil liberation: A model to explain high biodiversity in the Triassic Cassian Formation. Palaeontology 2020, 63, 85–102. [Google Scholar] [CrossRef] [Green Version]
- Kriwet, J.; Klug, S. Diversity and biogeography patterns of Late Jurassic neoselachians (Chondrichthyes: Elasmobranchii). Geol. Soc. Spec. Publ. 2008, 295, 55–70. [Google Scholar] [CrossRef]
- Candoni, L. Deux faunes inédites de sélaciens dans le Jurassique terminal Français-Premiers résultats stratigraphiques. SGNAMH 1995, 82, 29–49. [Google Scholar]
- Röper, M.; Reich, M. Overview of the distribution of echinoderms in the Upper Jurassic limestone (Kimmeridgium–tithonium) of the Solnhofen archipelago (Bavaria, Germany). Zitteliana 2018, 92, 37–40. [Google Scholar]
- Benson, R.B.J.; Butler, R.J. Uncovering the diversification history of marine tetrapods: Ecology influences the effect of geological sampling biases. Geol. Soc. Spec. Publ. 2011, 358, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Flannery Sutherland, J.T.; Moon, B.C.; Stubbs, T.L.; Benton, M.J. Does exceptional preservation distort our view of disparity in the fossil record? Proc. Royal Soc. B 2019, 286, 20190091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wignall, P.B. Benthic palaeoecology of the late Jurassic Kimmeridge Clay of England. Spec. Pap Palaeontol. 1990, 43, 1–74. [Google Scholar]
- Batchelor, T.J.; Ward, D.J. Fish remains from a temporary exposure of Hythe Beds (Aptian-Lower Cretaceous) near Godstone, Surrey. Mesozoic Res. 1990, 2, 181–203. [Google Scholar]
- Underwood, C.J. Environmental controls on the distribution of neoselachian sharks and rays within the British Bathonian (Middle Jurassic). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 203, 107–126. [Google Scholar] [CrossRef] [Green Version]

















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalobos-Segura, E.; Stumpf, S.; Türtscher, J.; Jambura, P.L.; Begat, A.; López-Romero, F.A.; Fischer, J.; Kriwet, J. A Synoptic Review of the Cartilaginous Fishes (Chondrichthyes: Holocephali, Elasmobranchii) from the Upper Jurassic Konservat-Lagerstätten of Southern Germany: Taxonomy, Diversity, and Faunal Relationships. Diversity 2023, 15, 386. https://doi.org/10.3390/d15030386
Villalobos-Segura E, Stumpf S, Türtscher J, Jambura PL, Begat A, López-Romero FA, Fischer J, Kriwet J. A Synoptic Review of the Cartilaginous Fishes (Chondrichthyes: Holocephali, Elasmobranchii) from the Upper Jurassic Konservat-Lagerstätten of Southern Germany: Taxonomy, Diversity, and Faunal Relationships. Diversity. 2023; 15(3):386. https://doi.org/10.3390/d15030386
Chicago/Turabian StyleVillalobos-Segura, Eduardo, Sebastian Stumpf, Julia Türtscher, Patrick L. Jambura, Arnaud Begat, Faviel A. López-Romero, Jan Fischer, and Jürgen Kriwet. 2023. "A Synoptic Review of the Cartilaginous Fishes (Chondrichthyes: Holocephali, Elasmobranchii) from the Upper Jurassic Konservat-Lagerstätten of Southern Germany: Taxonomy, Diversity, and Faunal Relationships" Diversity 15, no. 3: 386. https://doi.org/10.3390/d15030386