

Pinus contorta Alters Microenvironmental Conditions and Reduces Plant Diversity in Patagonian Ecosystems
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
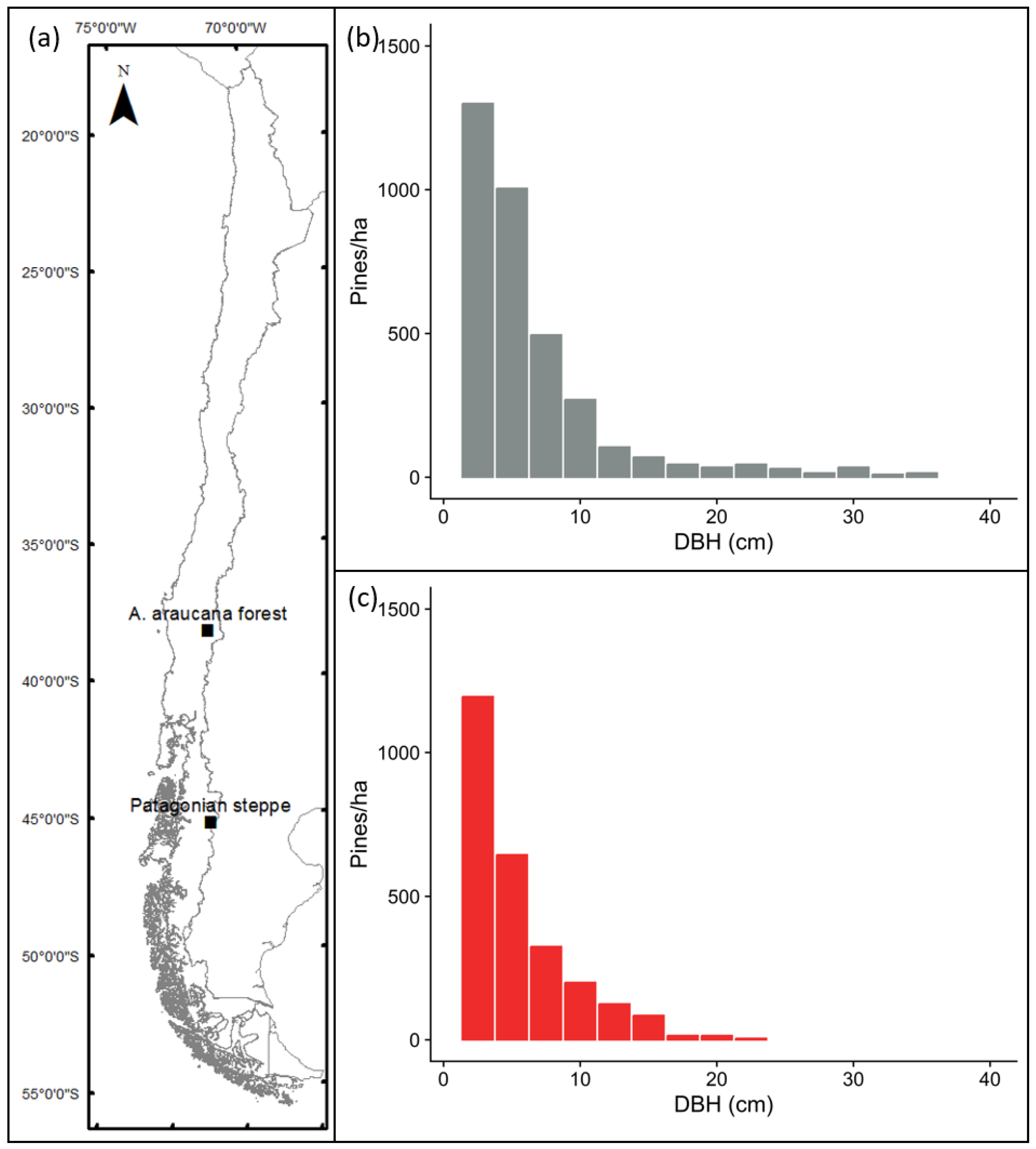
2.1. Study Area
2.2. Experimental Design
2.3. Data Analysis
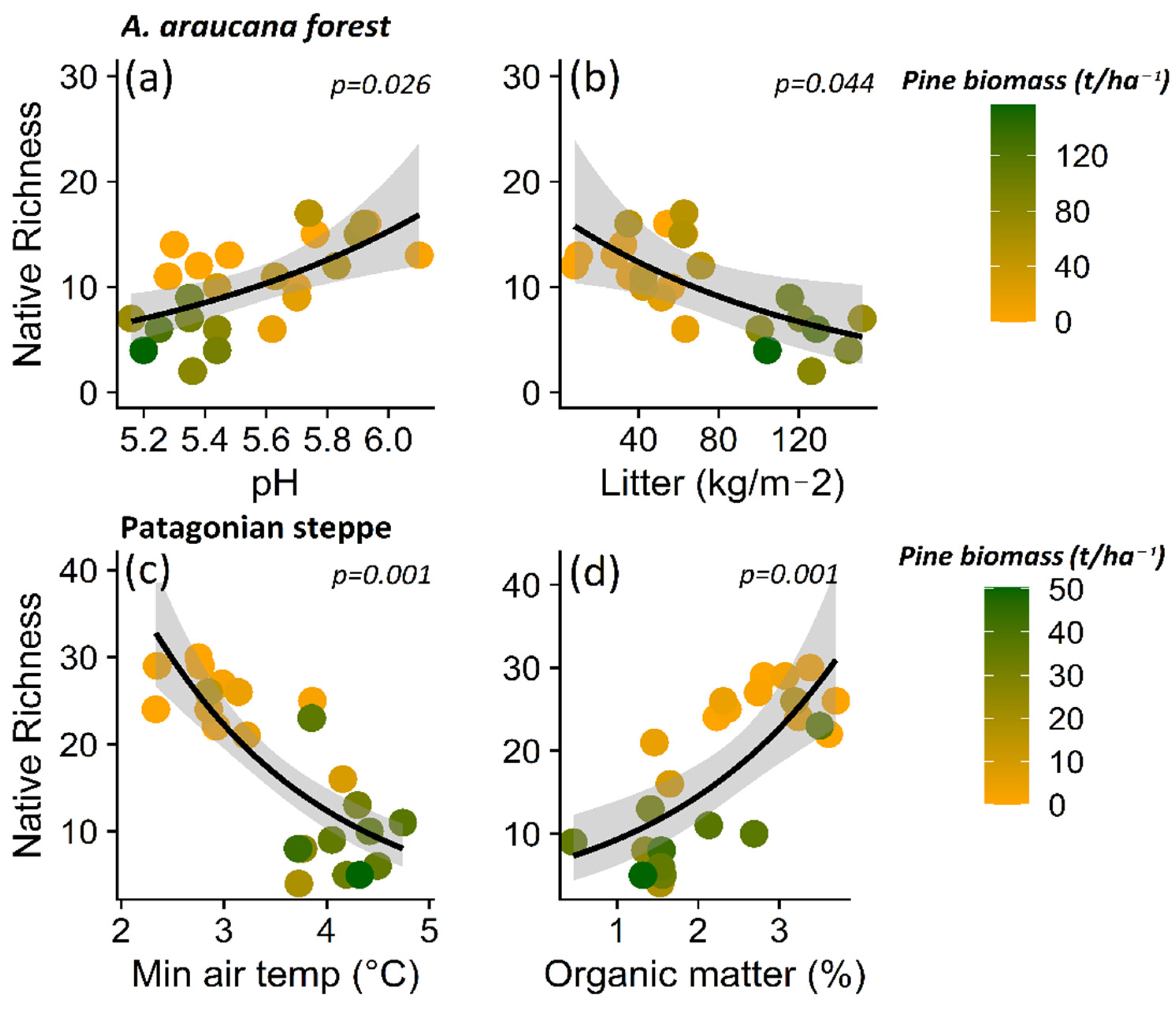
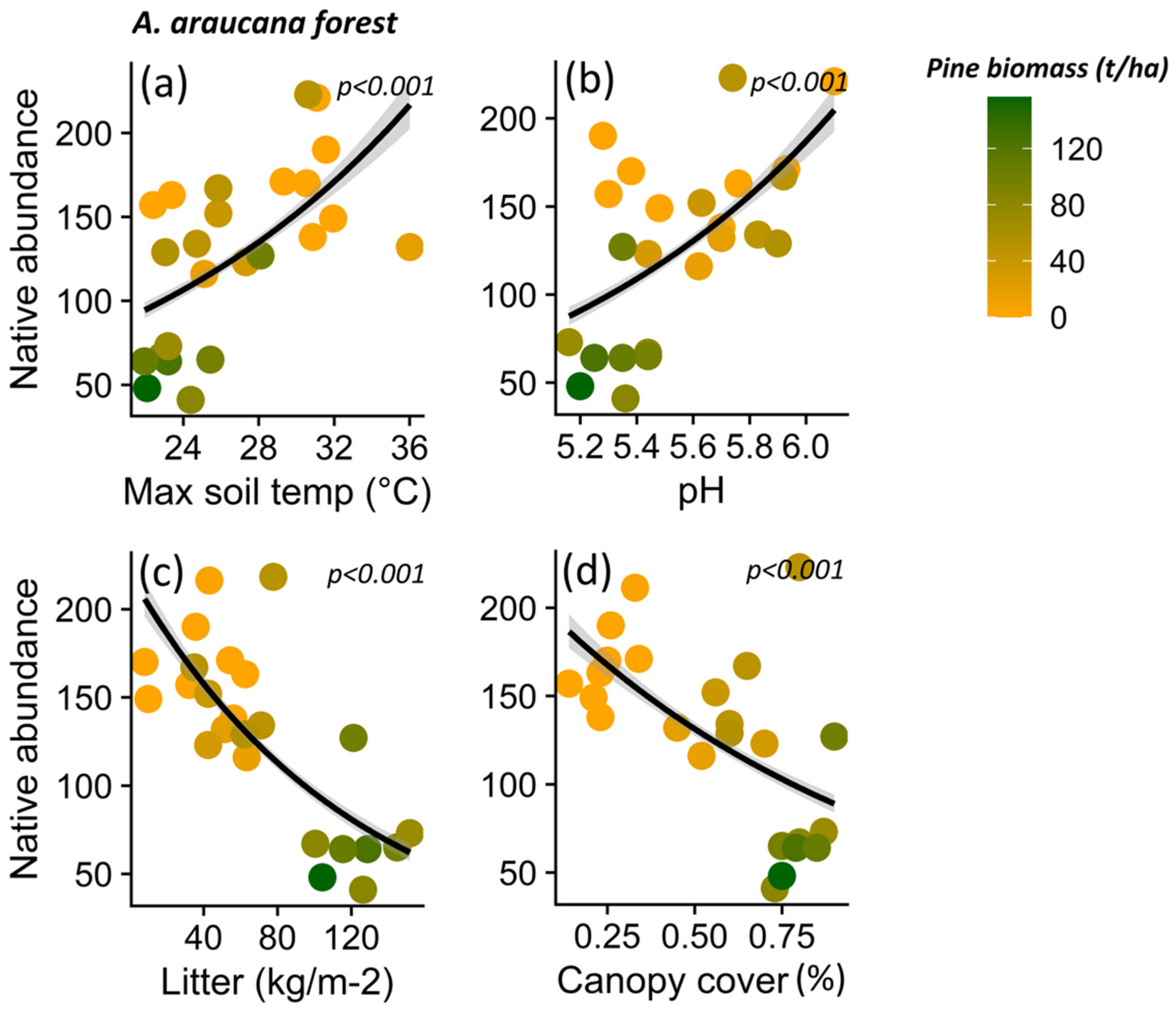
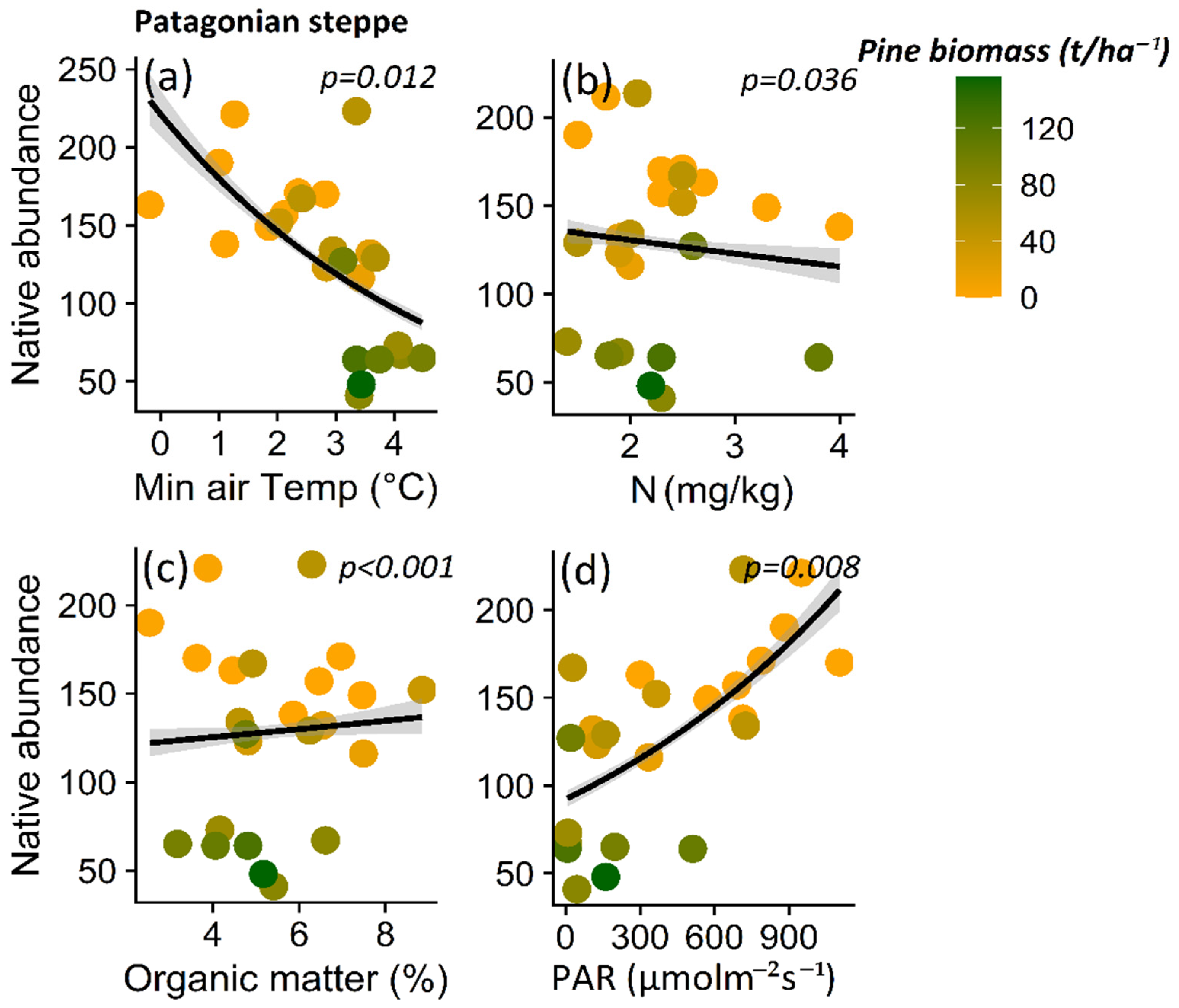
3. Results
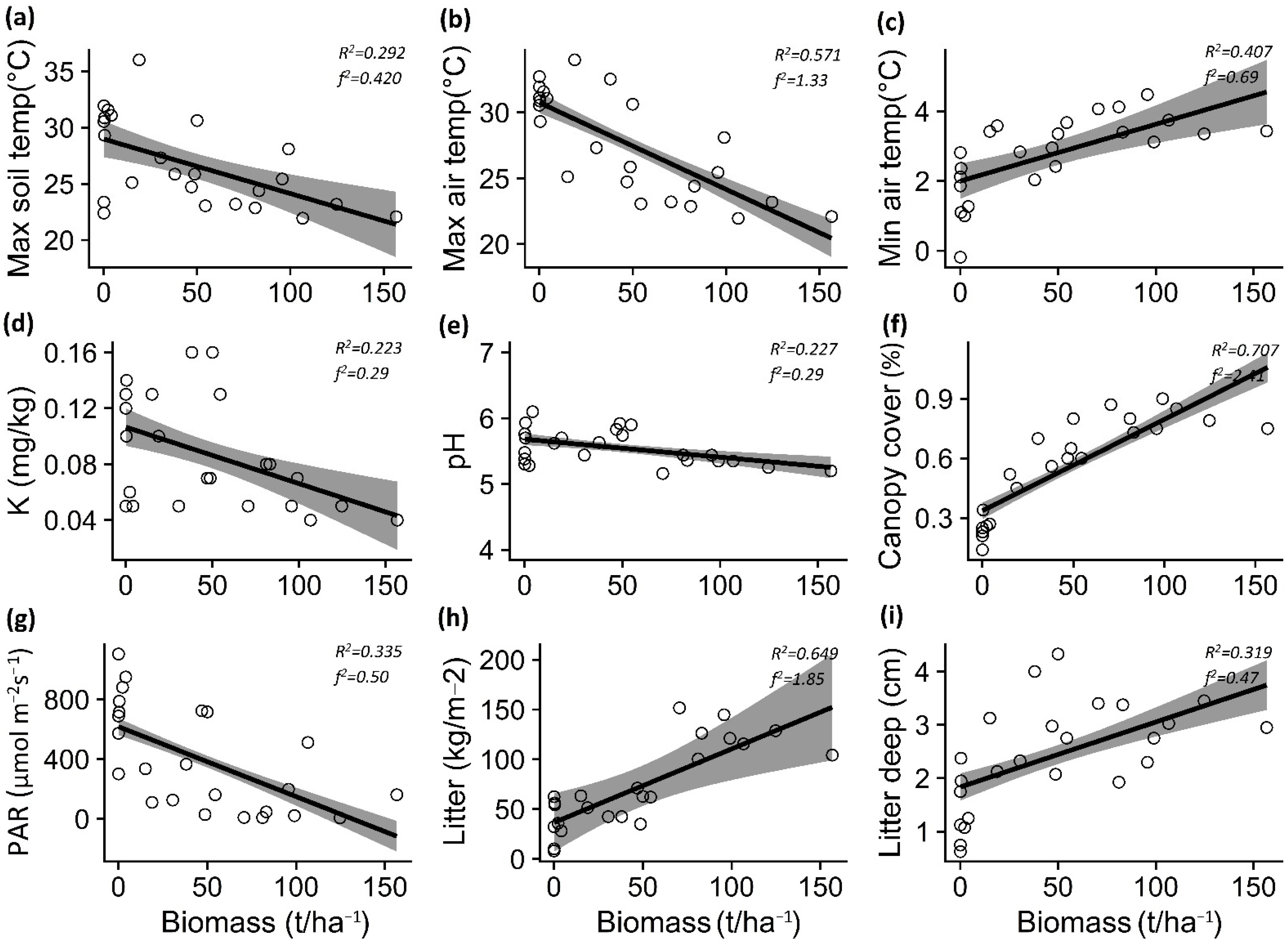
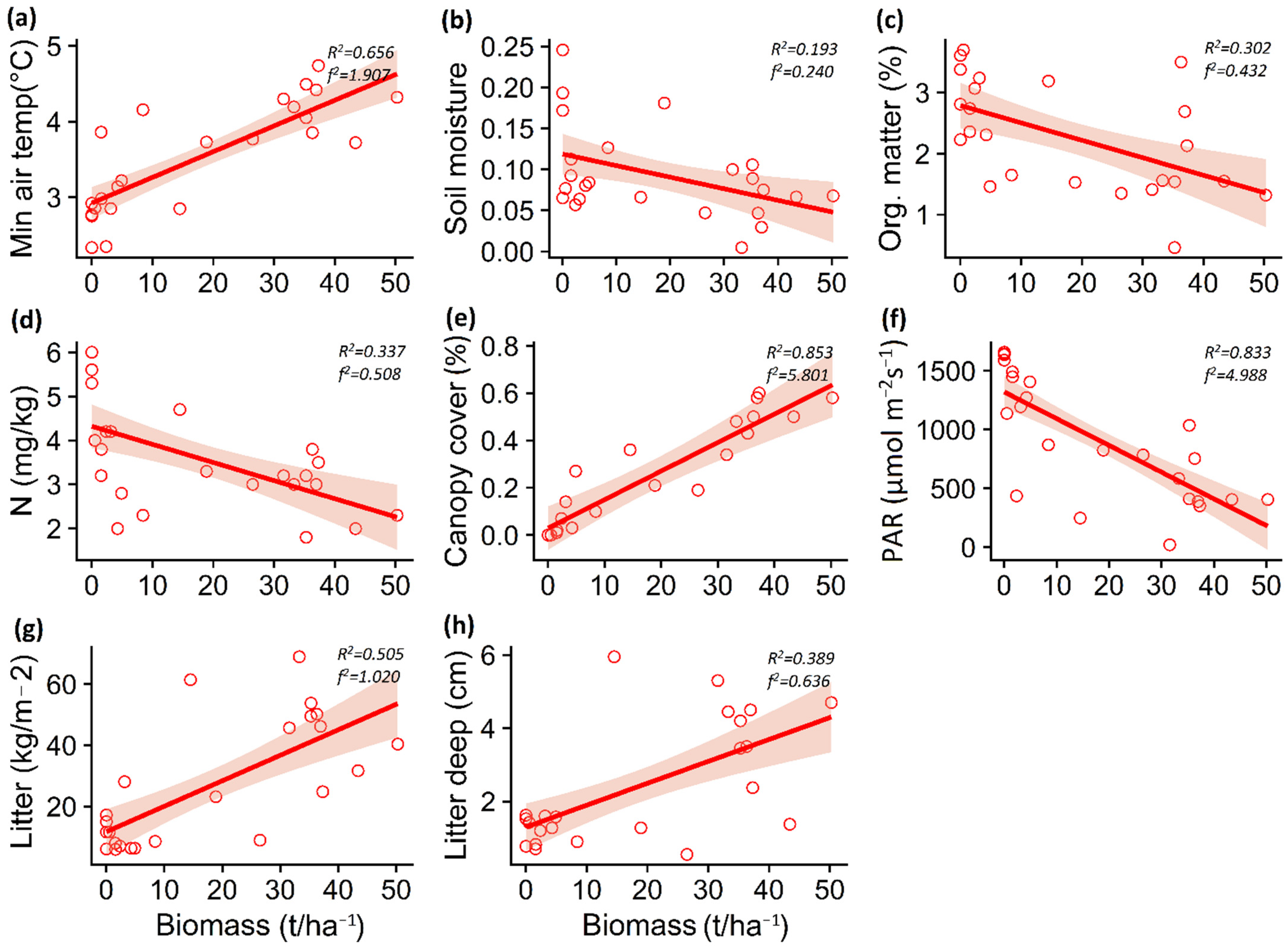
3.1. Impact of P. contorta Invasion on Environmental Conditions
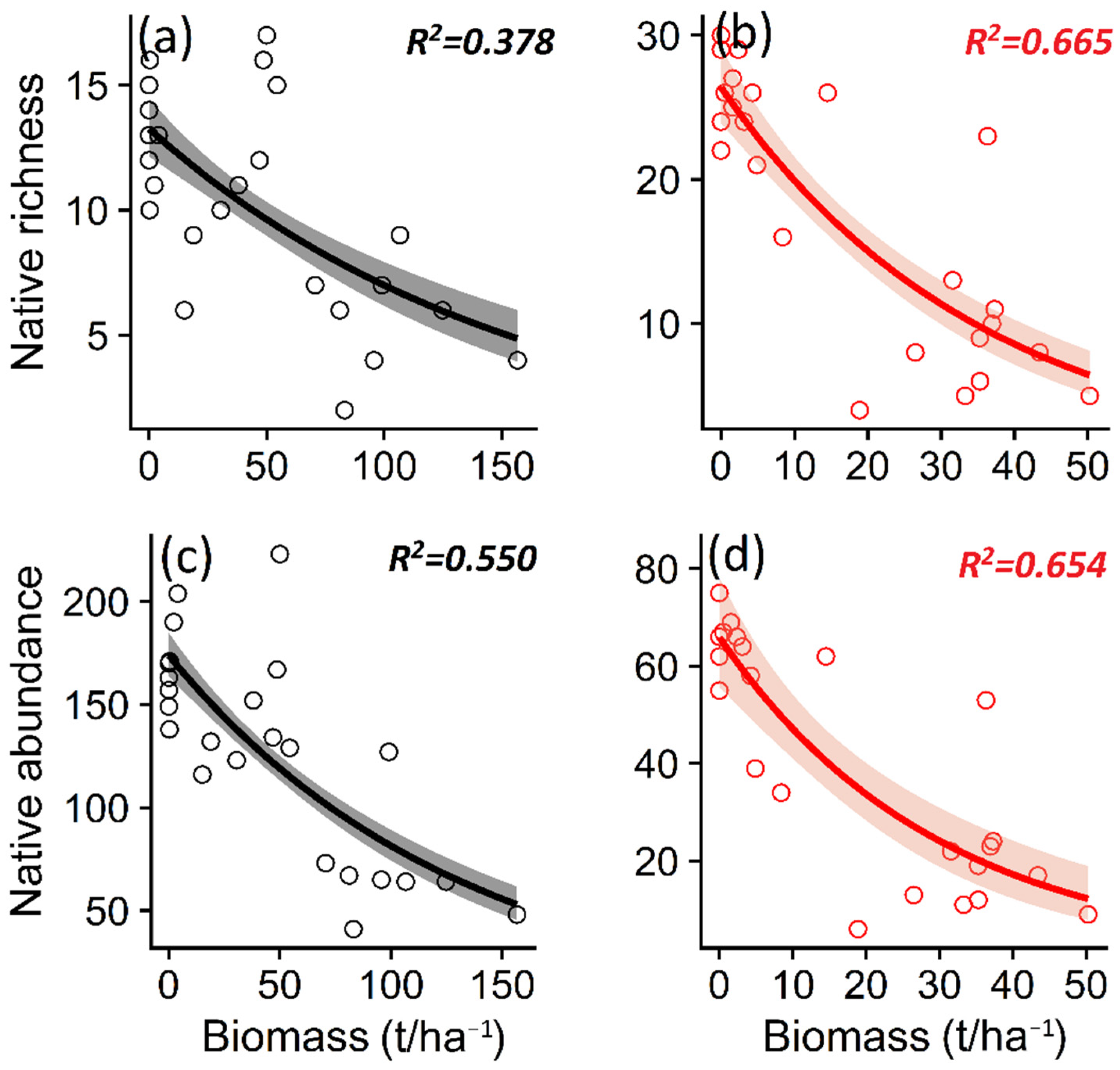
3.2. Effect of Microenvironmental Change on Native Plant Diversity in Ecosystems Invaded by P. contorta
4. Discussion
4.1. Impact of P. contorta Invasion on Microenvironmental Conditions
4.2. Variables Explaining the Reduction in Native Plant Richness in Invaded Ecosystems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Brundu, G.; Pauchard, A.; Pyšek, P.; Pergl, J.; Bindewald, A.M.; Brunori, A.; Canavan, S.; Campagnaro, T.; Celesti-Grapow, L.; de Sá Dechoum, M.; et al. Global Guidelines for the Sustainable Use of Non-Native Trees to Prevent Tree Invasions and Mitigate Their Negative Impacts. NeoBiota 2020, 61, 65–116. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Alonso, Á.; Saldaña-López, A.; Granda, E. Effects of Widespread Non-Native Trees on Regulating Ecosystem Services. Sci. Total Environ. 2021, 778, 146141. [Google Scholar] [CrossRef] [PubMed]
- Chikowore, G.; Martin, G.D.; Chidawanyika, F. An Assessment of the Invasive Alien Tree, Robinia pseudoacacia Canopy Traits and Its Effect on Grassland Microclimates and Subsequent Arthropod Assemblages. J. Insect Conserv. 2021, 25, 429–439. [Google Scholar] [CrossRef]
- Zeidler, M.; Duchoslav, M.; Banaš, M.; Lešková, M. Impacts of Introduced Dwarf Pine (Pinus mugo) on the Diversity and Composition of Alpine Vegetation. Community Ecol. 2012, 13, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Terwei, A.; Zerbe, S.; Mölder, I.; Annighöfer, P.; Kawaletz, H.; Ammer, C. Response of Floodplain Understorey Species to Environmental Gradients and Tree Invasion: A Functional Trait Perspective. Biol. Invasions 2016, 18, 2951–2973. [Google Scholar] [CrossRef]
- Franzese, J.; Urrutia, J.; García, R.A.; Taylor, K.; Pauchard, A. Pine Invasion Impacts on Plant Diversity in Patagonia: Invader Size and Invaded Habitat Matter. Biol. Invasions 2017, 19, 1015–1027. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. How Do Invasive Trees Impact Shrub Layer Diversity and Productivity in Temperate Forests? Ann. For. Sci. 2021, 78, 20. [Google Scholar] [CrossRef]
- Richardson, D.M.; Rejmánek, M. Conifers as Invasive Aliens: A Global Survey and Predictive Framework. Divers. Distrib. 2004, 10, 321–331. [Google Scholar] [CrossRef]
- Simberloff, D.; Nuñez, M.A.; Ledgard, N.J.; Pauchard, A.; Richardson, D.M.; Sarasola, M.; Van Wilgen, B.W.; Zalba, S.M.; Zenni, R.D.; Bustamante, R.; et al. Spread and Impact of Introduced Conifers in South America: Lessons from Other Southern Hemisphere Regions. Austral Ecol. 2010, 35, 489–504. [Google Scholar] [CrossRef]
- Sapsford, S.J.; Brandt, A.J.; Davis, K.T.; Peralta, G.; Dickie, I.A.; Gibson, R.D.; Green, J.L.; Hulme, P.E.; Nuñez, M.A.; Orwin, K.H.; et al. Towards a Framework for Understanding the Context Dependence of Impacts of Non-Native Tree Species. Funct. Ecol. 2020, 34, 944–955. [Google Scholar] [CrossRef]
- Peña, E.; Hidalgo, M.; Langdon, B.; Pauchard, A. Patterns of Spread of Pinus contorta Dougl. Ex Loud. Invasion in a Natural Reserve in Southern South America. For. Ecol. Manag. 2008, 256, 1049–1054. [Google Scholar] [CrossRef]
- Echeverria, C.; Coomes, D.; Salas, J.; Rey-Benayas, J.M.; Lara, A.; Newton, A. Rapid Deforestation and Fragmentation of Chilean Temperate Forests. Biol. Conserv. 2006, 130, 481–494. [Google Scholar] [CrossRef]
- Pauchard, A.; Bustamante, R.O. Forest Ecology and Management Habitat Suitability of Five Commonly Planted Non-Native Trees in Chile: Implications for an Invasion Process. For. Ecol. Manag. 2023, 529, 120726. [Google Scholar] [CrossRef]
- Langdon, B.; Pauchard, A.; Aguayo, M. Pinus contorta Invasion in the Chilean Patagonia: Local Patterns in a Global Context. Biol. Invasions 2010, 12, 3961–3971. [Google Scholar] [CrossRef]
- Langdon, B.; Cavieres, L.A.; Pauchard, A. At a Microsite Scale, Native Vegetation Determines Spatial Patterns and Survival of Pinus contorta Invasion in Patagonia. Forests 2019, 10, 654. [Google Scholar] [CrossRef] [Green Version]
- Davis, K.T.; Maxwell, B.D.; Caplat, P.; Pauchard, A.; Nuñez, M.A. Simulation Model Suggests That Fire Promotes Lodgepole Pine (Pinus contorta) Invasion in Patagonia. Biol. Invasions 2019, 1, 2287–2300. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.T.; Maxwell, B.D.; McWethy, D.B.; Pauchard, A.; Nuñez, M.A.; Whitlock, C. Pinus contorta Invasions Increase Wildfire Fuel Loads and May Create a Positive Feedback with Fire. Ecology 2017, 98, 678–687. [Google Scholar] [CrossRef] [Green Version]
- Cóbar-Carranza, A.J.; García, R.A.; Pauchard, A.; Peña, E. Effect of Pinus contorta Invasion on Forest Fuel Properties and Its Potential Implications on the Fire Regime of Araucaria Araucana and Nothofagus Antarctica Forests. Biol. Invasions 2014, 16, 2273–2291. [Google Scholar] [CrossRef]
- Bravo-Monasterio, P.; Pauchard, A.; Fajardo, A. Pinus contorta Invasion into Treeless Steppe Reduces Species Richness and Alters Species Traits of the Local Community. Biol. Invasions 2016, 18, 1883–1894. [Google Scholar] [CrossRef]
- Dickie, I.A.; St John, M.G.; Yeates, G.W.; Morse, C.W.; Bonner, K.I.; Orwin, K.; Peltzer, D.A. Belowground Legacies of Pinus contorta Invasion and Removal Result in Multiple Mechanisms of Invasional Meltdown. AoB Plants 2014, 6, plu056. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Wickham, H. Ggplot2: Elganta Graphics for Data Analysis; Springer: New York, NY, USA, 2008; ISBN 978-0-387-78170-9. [Google Scholar] [CrossRef]
- Taylor, K.T.; Maxwell, B.D.; Pauchard, A.; Nuñez, M.A.; Rew, L.J. Native versus Non-Native Invasions: Similarities and Differences in the Biodiversity Impacts of Pinus contorta in Introduced and Native Ranges. Divers. Distrib. 2016, 22, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Solfiyeni, S.; Syamsuardi, S.; Chairul, C.; Mukhtar, E. Impacts of Invasive Tree Species Bellucia pentamera on Plant Diversity, Microclimate and Soil of Secondary Tropical Forest in West Sumatra, Indonesia. Biodiversitas 2022, 23, 3135–3146. [Google Scholar] [CrossRef]
- Slabejová, D.; Bacigál, T.; Hegedüšová, K.; Májeková, J.; Medvecká, J.; Mikulová, K.; Šibíková, M.; Škodová, I.; Zaliberová, M.; Jarolímek, I. Comparison of the Understory Vegetation of Native Forests and Adjacent Robinia pseudoacacia Plantations in the Carpathian-Pannonian Region. For. Ecol. Manag. 2019, 439, 28–40. [Google Scholar] [CrossRef]
- Urrutia, J.; Pauchard, A.; García, R.A. Diferencias En La Composición Vegetal de Un Bosque de Araucaria araucana (Molina) K.Koch y Nothofagus antarctica (G. Forst.) Oerst. Asociadas a Un Gradiente de Invasión de Pinus contorta Douglas Ex Loudon. Gayana-Bot. 2013, 70, 92–100. [Google Scholar] [CrossRef]
- Pauchard, A.; Escudero, A.; García, R.A.; de la Cruz, M.; Langdon, B.; Cavieres, L.A.; Esquivel, J. Pine Invasions in Treeless Environments: Dispersal Overruns Microsite Heterogeneity. Ecol. Evol. 2016, 6, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzese, J.; Raffaele, E.; Chiuffo, M.C.; Blackhall, M. The Legacy of Pine Introduction Threatens the Fuel Traits of Patagonian Native Forests. Biol. Conserv. 2022, 267, 109472. [Google Scholar] [CrossRef]
- Erckie, L.; Adedoja, O.; Geerts, S.; van Wyk, E.; Boatwright, J.S. Impacts of an Invasive Alien Proteaceae on Native Plant Species Richness and Vegetation Structure. South Afr. J. Bot. 2022, 144, 332–338. [Google Scholar] [CrossRef]
- González-Muñoz, N.; Costa-Tenorio, M.; Espigares, T. Invasion of Alien Acacia Dealbata on Spanish Quercus Robur Forests: Impact on Soils and Vegetation. For. Ecol. Manag. 2012, 269, 214–221. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Rawai, A. Impacts of the Invasive Exotic Prosopis juliflora (Sw.) D.C. on the Native Flora and Soils of the UAE. Plant Ecol. 2007, 190, 23–35. [Google Scholar] [CrossRef]
- Vilà, M.; Tessier, M.; Suehs, C.M.; Brundu, G.; Carta, L.; Galanidis, A.; Lambdon, P.; Manca, M.; Médail, F.; Moragues, E.; et al. Local and Regional Assessments of the Impacts of Plant Invaders on Vegetation Structure and Soil Properties of Mediterranean Islands. J. Biogeogr. 2006, 33, 853–861. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate Change Threats to Plant Diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.I.; McLaughlin, Ó.B.; Marteinsdóttir, B.; O’Gorman, E.J. Soil Temperature Effects on the Structure and Diversity of Plant and Invertebrate Communities in a Natural Warming Experiment. J. Anim. Ecol. 2018, 87, 634–646. [Google Scholar] [CrossRef] [Green Version]
- De Sassi, C.; Lewis, O.T.; Tylianakis, J.M. Plant-Mediated and Nonadditive Effects of Two Global Change Drivers on an Insect Herbivore Community. Ecology 2012, 93, 1892–1901. [Google Scholar] [CrossRef] [PubMed]
- Forrest, J.R.K. Plant-Pollinator Interactions and Phenological Change: What Can We Learn about Climate Impacts from Experiments and Observations? Oikos 2015, 124, 4–13. [Google Scholar] [CrossRef]
- Giejsztowt, J.; Classen, A.T.; Deslippe, J.R. Climate Change and Invasion May Synergistically Affect Native Plant Reproduction. Ecology 2020, 101, e02913. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D.; Von Holle, B. Positive Interactions of Nonindigenous Species. Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Sagarin, R.; Pauchard, A. Observation and Ecology: Broadening the Scope of Science to Understand a Complex World; Island Press: Washington, DC, USA, 2013; pp. 1–213. [Google Scholar]
- Dickie, I.A.; Bennett, B.M.; Burrows, L.E.; Nuñez, M.A.; Peltzer, D.A.; Porté, A.; Richardson, D.M.; Rejmánek, M.; Rundel, P.W.; van Wilgen, B.W. Conflicting Values: Ecosystem Services and Invasive Tree Management. Biol. Invasions 2014, 16, 705–719. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. araucana Forest | Patagonia Steppe | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Estimated | SE | t-Value | p-Value | f2 * | Estimated | SE | t-Value | p-Value | f2 * | |
| Temp. max. soil | −0.048 | 0.014 | −3.244 | 0.003 | 0.42 | −0.092 | 0.048 | −1.923 | 0.067 | 0.14 |
| Temp. max. air | −0.065 | 0.011 | −5.624 | <0.001 | 1.33 | 0.020 | 0.026 | 0.762 | 0.454 | 0.02 |
| Temp. min. soil | −0.001 | 0.011 | −0.167 | 0.869 | 0.01 | −0.006 | 0.012 | −0.479 | 0.636 | 0.01 |
| Temp. min. air | 0.016 | 0.003 | 4.104 | 0.004 | 0.69 | 0.033 | 0.005 | 6.478 | <0.001 | 1.90 |
| Soil moisture | −0.007 | 0.001 | −0.394 | 0.697 | 0.01 | −0.001 | 0.006 | −2.298 | 0.031 | 0.24 |
| Organic matter | −0.006 | 0.006 | −0.921 | 0.367 | 0.04 | −0.028 | 0.009 | −3.088 | 0.005 | 0.43 |
| Nitrate (NO3-) | −0.006 | 0.003 | −0.203 | 0.841 | 0.01 | −0.041 | 0.012 | −3.347 | 0.002 | 0.50 |
| Phosphorus (P) | 0.010 | 0.021 | 0.505 | 0.618 | 0.29 | −0.017 | 0.029 | −0.606 | 0.551 | 0.05 |
| Potassium (K) | −0.004 | 0.001 | −2.517 | 0.019 | 0.29 | −0.001 | 0.001 | −1.165 | 0.257 | 0.01 |
| pH | −0.002 | 0.001 | −2.547 | 0.018 | 0.29 | <0.006 | 0.001 | 0.05 | 0.960 | 0.01 |
| Canopy cover | 0.004 | 0.006 | 7.518 | <0.001 | 2.41 | 0.012 | 0.009 | 12.486 | <0.001 | 5.80 |
| PAR | −4.685 | 1.267 | −3.697 | 0.001 | 0.50 | −22.716 | 4.144 | −5.482 | <0.001 | 4.98 |
| Total Litter | 0.738 | 0.0005 | 6.390 | <0.001 | 1.85 | 0.833 | 0.175 | 4.739 | <0.001 | 1.02 |
| Litter depth | 0.012 | 0.003 | 3.215 | 0.003 | 0.47 | 0.059 | 0.015 | 3.750 | 0.001 | 0.63 |
| Non-nat. richness | −0.010 | 0.003 | −2.942 | 0.007 | 0.39 | −0.040 | 0.009 | −4.052 | 0.005 | 0.42 |
| Non-nat. abundance | −0.204 | 0.041 | −4.973 | <0.001 | 1.12 | −0.165 | 0.027 | −5.996 | <0.001 | 0.62 |
| Nat. richness | −0.058 | 0.015 | −3.846 | 0.008 | 0.67 | −0.435 | 0.063 | −6.842 | <0.001 | 0.68 |
| Nat. abundance | −0.855 | 0.158 | −5.400 | <0.001 | 1.33 | −1.185 | 0.177 | −6.681 | <0.001 | 0.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García, R.A.; Fuentes-Lillo, E.; Cavieres, L.; Cóbar-Carranza, A.J.; Davis, K.T.; Naour, M.; Núñez, M.A.; Maxwell, B.D.; Lembrechts, J.J.; Pauchard, A. Pinus contorta Alters Microenvironmental Conditions and Reduces Plant Diversity in Patagonian Ecosystems. Diversity 2023, 15, 320. https://doi.org/10.3390/d15030320
García RA, Fuentes-Lillo E, Cavieres L, Cóbar-Carranza AJ, Davis KT, Naour M, Núñez MA, Maxwell BD, Lembrechts JJ, Pauchard A. Pinus contorta Alters Microenvironmental Conditions and Reduces Plant Diversity in Patagonian Ecosystems. Diversity. 2023; 15(3):320. https://doi.org/10.3390/d15030320
Chicago/Turabian StyleGarcía, Rafael A., Eduardo Fuentes-Lillo, Lohengrin Cavieres, Ana J. Cóbar-Carranza, Kimberley T. Davis, Matías Naour, Martín A. Núñez, Bruce D. Maxwell, Jonas J. Lembrechts, and Aníbal Pauchard. 2023. "Pinus contorta Alters Microenvironmental Conditions and Reduces Plant Diversity in Patagonian Ecosystems" Diversity 15, no. 3: 320. https://doi.org/10.3390/d15030320