
First Record of Osphya (Melandryidae: Osphyinae) from Chinese Mainland Based on Morphological Evidence and Mitochondrial Genome-Based Phylogeny of Tenebrionoidea †
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Photography and Terminology
2.3. DNA Extraction, PCR, and Sequencing
2.4. Phylogenetic Analyses
3. Results
3.1. Description of New Species
Osphya sinensis H. Liu et Y. Yang, sp. n.
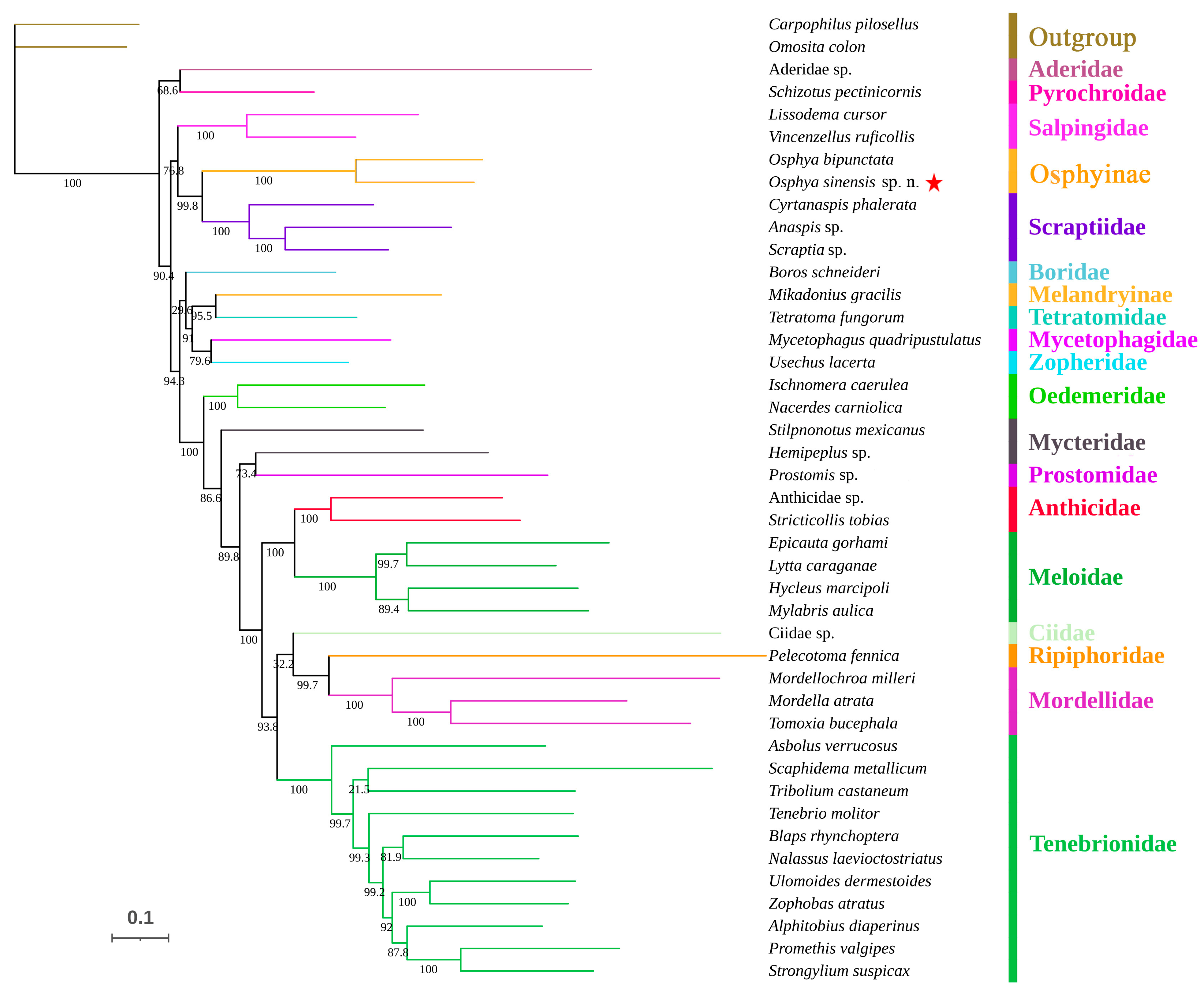
3.2. Phylogenetic Analyses
4. Discussion
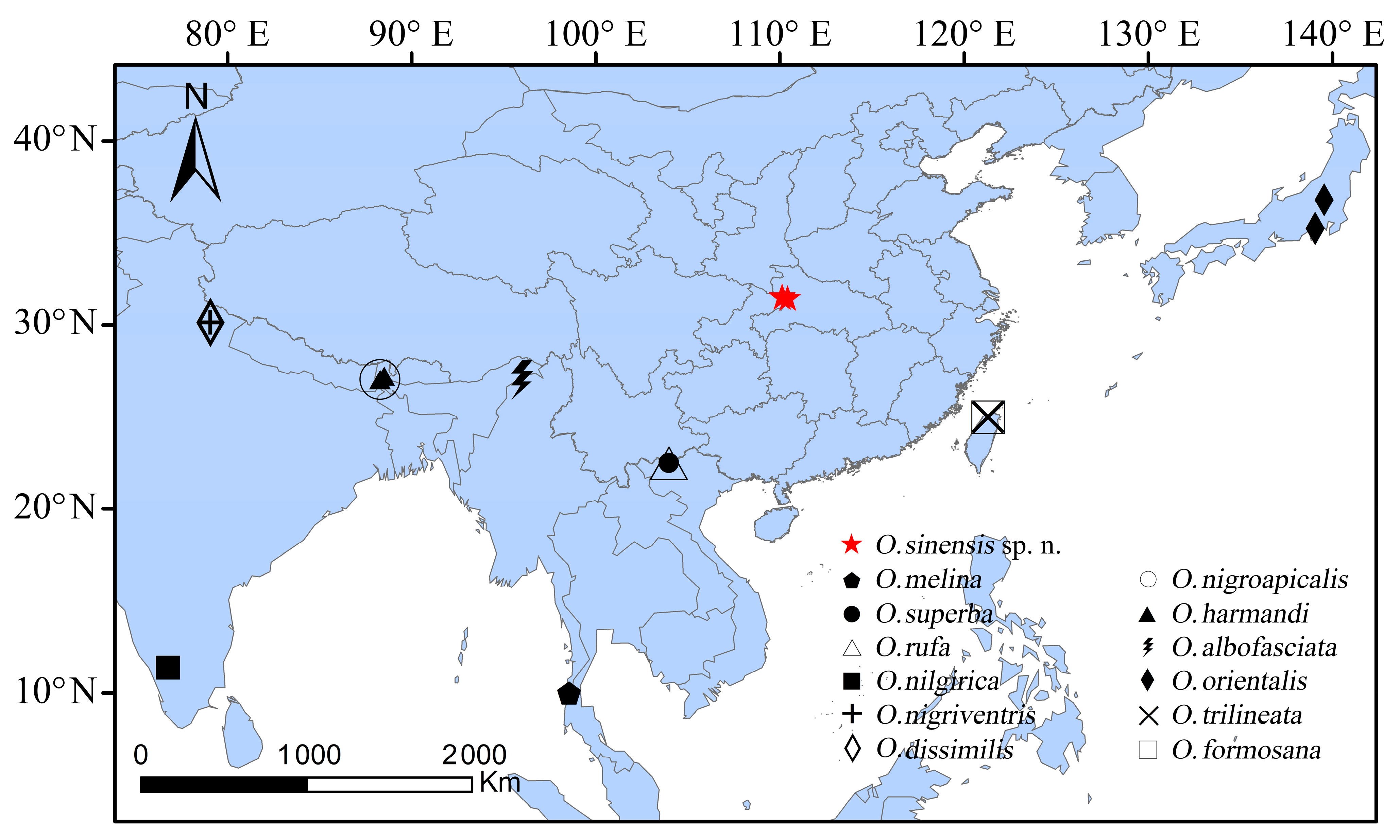
4.1. Separate Status of the New Species
| Key to the Osphya species in the Oriental and eastern and southern Palaearctic regions |
| 1. Body large-sized, more than 10 mm; elytra uniformly green or blue, with strongly metallic shine……………………………………………………………………………………..2 |
| - Body small or middle-sized, less than 8 mm; elytra black, yellow or reddish brown, or bicolored, at most with weakly metallic shine………………………………………………..3 |
| 2. Body 12 mm in length; body green, scutellum and legs uniformly orange……………………………………………………………………………………..O. superba |
| - Body 9.5–10 mm in length; body blue, scutellum dark blue, legs bicolored, mixed blue with orange..…………………………………………………………….………O. sinensis sp. n. |
| 3. Body covered with reddish-brown pubescences…………………………………………..4 |
| - Body covered with gray or black pubescences……………………………………………...5 |
| 4. Body 7 mm in length; elytra uniformly reddish brown……………………………. O. rufa |
| - Body 8 mm in length; elytra reddish brown, black at apices………………. O. nigroapicalis |
| 5. Elytra decorated with gray longitudinal or transverse bands……………………….……6 |
| - Elytra uniformly colored, without any longitudinal or transverse band……….………...8 |
| 6. Body 6–6.5 mm in length; each elytron with a longitudinal band along the suture and two transverse bands on the disc...……………………………………………...….O. orientalis |
| - Elytra with gray pubescent bands along sutures and on each side of the middle disc…7 |
| 7. Body slender, 6 mm in length; antennomeres IV and V relatively long……...O. trilineata |
| - Body stouter, 5 mm in length; antennomeres IV and V relatively short……..O. albofaciata |
| 8. Body mostly or uniformly yellow…………………………………………………………...9 |
| - Body black……………………………………………………………………………………...10 |
| 9. Body small and 4.25 mm in length, uniformly yellow……………………………O. melina |
| - Body middle-sized and 7.5 mm in length, mostly yellow, black at anterior and middle tibiae, apex of terminal maxillary palpomere, and antennomeres IV-XI……O. nigriventris |
| 10. Body 5.5 mm in length; antennomeres II, IV, and V subequal in length, III, VI, and VII slightly longer, VIII–XI stouter……………………………………………………...O. dissimilis |
| - Body longer than 6.0 mm; antennae unlike above…………………………………………11 |
| 11. Body 7.0 mm in length; elytra with weakly metallic shine…………………O. formosana |
| - Elytra without metallic shine………………………………………………………………...12 |
| 12. Body slender and 8.0 mm in length; head smaller……………………………O. harmandi |
| - Body stouter and 6.0 mm in length; head broader………………………………O. nilgirica |
4.2. Phylogenetic Position of the New Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikitsky, N.B.; Pollock, D.A. Melandryidae Leach, 1815. In Catalogue of Palaearctic Coleoptera, Volume 5: Tenebrionoidea; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2008; pp. 64–73. [Google Scholar]
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Reid, C.A.M.; Schmitt, M.; Slipinski, S.A.; et al. Family-group names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konvička, O. Osphya lehnertae sp. nov. from Greece (Coleopteta: Melandryidae). Klapalekiana. 2014, 50, 161–166. [Google Scholar]
- Konvička, O. Osphya brusteli sp. nov. from the Balkan Peninsula (Coleopteta: Melandryidae). Acta. Mus. Silesi. Sci. Natural. 2016, 65, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Lewis, G. On the Cistelidae and other heteromerous species of Japan. Ann. Mag. Nat. Hist. 1895, 15, 250–278, 422–448, pl. 8. [Google Scholar] [CrossRef]
- Champion, G.C. Melandryidae. In Biologia Centrali-Americana. Insecta, Coleoptera, Volume IV, part 2. [1889,1890,1891,1892,1893]; R. H. Porter: London, UK, 1889; pp. 75–103, 5 pls. [Google Scholar]
- Champion, G.C. Notes on Melandryidae (2–3). Entomol. Mon. Mag. 1916, 52, 99–108. [Google Scholar]
- Champion, G.C. Some Indian Coleoptera (2). Entomol. Mon. Mag. 1920, 56, 68–77. [Google Scholar]
- Champion, G.C. Some Indian Coleoptera (7). Entomol. Mon. Mag. 1922, 58, 68–76. [Google Scholar]
- Pic, M. Coleopteres exotiques nouveaux oupeuconnus (Suite). L’Echange. Rev. Linn. 1910, 26, 74–78. [Google Scholar]
- Pic, M. Notes diverses, descriptions et diagnoses (Suite). L’Echange Rev. Linn. 1921, 37, 9–10. [Google Scholar]
- Pic, M. Diagnoses de coleopteres exotiques (Suite). L’Echange Rev. Linn. 1921, 37, 13–15. [Google Scholar]
- Pic, M. Nouveaux coleopteres exotiques. Bull. Mus. Natl. Hist. Nat. Paris 1926, 32, 354–359. [Google Scholar] [CrossRef]
- Pic, M.H. Sauter’s Formosa-Ausbeute: Heteromera [ex parte] (Col.). Entomol. Mitt. 1927, 16, 48–49. [Google Scholar]
- Pic, M. Coléoptères de l’Indochine. Mél. Exot.-Entomol. 1927, 49, 1–36. [Google Scholar]
- Pic, M. Notes diverses, nouveautes (Suite). L’Echange Rev. Linn. 1937, 53, 1. [Google Scholar]
- Recalde Irurzun, J.I.; Konvička, O.; Torres, J.L. On Osphya valdalitidae (Kraatz, 1868) and the Iberian Osphyinae (Coleopteta: Melandryidae). Heteropterus Rev. Entomol. 2017, 17, 41–52. [Google Scholar]
- Wang, W.; Feng, C.T.; Liu, F.Z.; Li, J.S. Biodiversity conservation in China: A review of recent studies and practices. Environ. Sci. Ecotechnol. 2020, 2, 100025. [Google Scholar] [CrossRef] [PubMed]
- Pollock, D.A. Melandryidae Leach, 1815. In American Beetles. Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea; Arnett, R.H., Jr., Thomas, M.C., Skelley, P.E., Frank, J.H., Eds.; CRC Press: Boca Raton, FL, USA, 2002; pp. 417–422. [Google Scholar]
- Nikitsky, N.B.; Pollock, D.A. Melandryidae Leach, 1815. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles. Morphology and Systematics; Volume 2 (Elateroidea, Bostrichiformia), Leschen, R.A.B., Beutel, R.G., Lawrence, J.F., Eds.; Walter de Gruyter: Berlin, Germany, 2010; pp. 520–533. [Google Scholar]
- Cameron, S.L.; Lambkin, C.L.; Barker, S.C.; Whiting, M.F. A mitochondrial genome of Diptera: Whole genome sequence data accurately resolve relationships over broad time scales with high precision. Syst. Entomol. 2007, 32, 40–59. [Google Scholar] [CrossRef]
- Cameron, S.L. Insect Mitochondrial Genomics: Implications for Evolution and Phylogeny. Ann. Rev. Entomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, F.; Jiang, P.; Wilson, J.J.; Cai, W.; Li, H. Compositional heterogeneity in true bug mitochondrial phylogenomics. Mol. Phylogenet. Evol. 2018, 118, 135–144. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Song, F.; Zhao, Y.; Wilson, J.J.; Cai, W. Higher-level phylogeny and evolutionary history of Pentatomorpha (Hemiptera: Heteroptera) inferred from mitochondrial genome sequences. Syst. Entomol. 2019, 44, 810–819. [Google Scholar] [CrossRef]
- Li, H.; Shao, R.; Song, N.; Song, F.; Jiang, P.; Li, Z.; Cai, W. Higher-level phylogeny of paraneopteran insects inferred from mitochondrial genome sequences. Sci. Rep. 2015, 5, 8527. [Google Scholar] [CrossRef] [PubMed]
- Curole, J.P.; Kocher, T.D. Mitogenomics: Digging deeper with complete mitochondrial genomes. Trends Ecol. Evol. 1999, 14, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Crampton-Platt, A.; Timmermans, M.J.T.N.; Gimmel, M.L.; Kutty, S.N.; Cockerill, T.D.; Khen, C.V.; Volger, A.P. Soup to Tree: The Phylogeny of Beetles Inferred by Mitochondrial Metagenomics of a Bornean Rainforest Sample. Mol. Biol. Evol. 2015, 32, 2302–2316. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, M.J.T.N.; Barton, C.; Haran, J.; Ahrens, D.; Culverwell, C.L.; Ollikainen, A.; Dodsworth, S.; Foster, P.G.; Bocak, L.; Vogler, A.P. Family-Level Sampling of Mitochondrial Genomes in Coleoptera: Compositional Heterogeneity and Phylogenetics. Genome Biol. 2016, 8, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linard, B.; Crampton-Platt, A.; Moriniere, J.; Timmermans, M.J.T.N.; Andújar, C.; Arribas, P.; Miller, K.E.; Lipecki, J.; Favreau, E.; Hunter, A.; et al. The contribution of mitochondrial metagenomics to large-scale data mining and phylogenetic analysis of Coleoptera. Mol. Phylogenet. Evol. 2018, 128, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, R.; Andújar, C.; Gómez-Rodríguez, C.; Bai, M.; Xue, H.; Tang, M.; Yang, C.; Tang, P.; Yang, X.; Vogler, A.P. The phylogeny of leaf beetles (Chrysomelidae) inferred from mitochondrial genomes. Syst. Entomol. 2019, 45, 188–204. [Google Scholar] [CrossRef]
- Nie, R.; Vogler, A.P.; Yang, X.K.; Lin, M. Higher-level phylogeny of longhorn beetles (Coleoptera: Chrysomeloidea) inferred from mitochondrial genomes. Syst. Entomol. 2020, 46, 56–70. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, Z.; Zheng, C.; Fang, J. Biodiversity in China’s mountains. Front. Ecol. Environ. 2006, 4, 347–352. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.; Liu, S.; Yang, Q.; Su, X.; Zhou, L.; Tang, M.; Fu, R.; Li, J.; Huang, Q. Ultra-deep sequencing enables high-fidelity recovery of biodiversity for bulk arthropod samples without PCR amplification. GigaScience 2013, 2, 2047-217X-2-4. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Gärtner, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, M.J.T.N.; Dodsworth, S.; Culverwell, C.L.; Bocak, L.; Ahrens., D.; Littlewood, D.T.J.; Pons, J.; Vogler, A.P. Why barcode? High-throughput multiplex sequencing of mitochondrial genomes for molecular systematics. Nucleic Acids Res. 2010, 38, e197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, C.; Zhang, L.F.; Lu, T.; Ma, J.N.; Zeng, C.J.; Yue, B.S.; Zhang, X.Y. Mitochondrial genomes of blister beetles (Coleoptera, Meloidae) and two large intergenic spacers in Hycleus genera. BMC Genom. 2017, 18, 698. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.L.; Sullivan, J.; Song, H.; Miller, K.B.; Whiting, M.F. A mitochondrial genome phylogeny of the Neuropterida (lace-wings, alderflies and snakeflies) and their relationship to the other holometabolous insect orders. Zool. Scr. 2009, 38, 575–590. [Google Scholar] [CrossRef]
- Hong, K.J.; Ki, W.; Park, D.S.; Yang, B.K.; Lee, H.; Park, J.; Lee, W. The complete mitochondrial genome of Alphitobius diaperinus Panzer, 1797 (Coleoptera: Tenebrionidae). Mitochondrial DNA Part B 2020, 5, 2291–2293. [Google Scholar] [CrossRef]
- Rider, S.D., Jr. The complete mitochondrial genome of the desert darkling beetle Asbolus verrucosus (Coleoptera, Tenebrionidae). Mitochondrial DNA Part A 2015, 27, 2447. [Google Scholar] [CrossRef]
- Zhao, M.; He, Z.; Wang, C.; Sun, L.; Feng, Y. Mitochondrial genome of a medicinal beetle Blaps rhynchopetera (Coleoptera, Tenebrionidae) and phylogenetic analysis. Mitochondrial DNA Part B 2019, 4, 2583–2584. [Google Scholar] [CrossRef] [Green Version]
- Linard, B.; Arribas, P.; Andújar, C.; Crampton-Platt, A.; Volger, A.P. Lessons from genome skimming of arthropod-preserving ethanol. Mol. Ecol. Resour. 2016, 16, 1365–1377. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Chen, J.; Li, G.; Luo, J.; Wang, H.; Yang, Y.; Liang, S.; Ouyang, B. Complete mitochondrial genome of Promethis valgipes valgipes (Marseul) (Insecta: Coleoptera: Tenebrionidae). Mitochondrial DNA Part B 2021, 6, 538–539. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.N.; Wang, C.Y. Complete mitochondrial genome of yellow mealworm (Tenebrio molitor). Zool. Res. 2014, 35, 93–101. [Google Scholar]
- Friedrich, M.; Muqim, N. Sequence and phylogenetic analysis of the complete mitochondrial genome of the flour beetle Tribolium castanaeum. Mol. Phylogenet. Evol. 2003, 26, 502–512. [Google Scholar] [CrossRef]
- Wang, C.Y.; Feng, Y.; Chen, X.M. The complete mitochondrial genome of a medicinal insect Martianus dermestoides (Coleoptera, Tenebrionidae). Mitochondrial DNA Part A 2014, 27, 1627–1628. [Google Scholar]
- Bai, Y.; Wang, H.; Li, G.; Luo, J.; Liang, S.; Li, C. Complete mitochondrial genome of the super mealworm Zophobas atratus (Fab.) (Insecta: Coleoptera: Tenebrionidae). Mitochondrial DNA Part B 2019, 4, 1300–1301. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Lan, Y.; Xia, L.; Cui, M.; Sun, W.; Dong, Z.; Cao, Y. The first complete mitochondrial genomes of two sibling species from nitidulid beetles Pests. Insects 2019, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zhou, H.; Zhang, J.; Li, W.; Wang, G. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2019, 20, 348–355. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. Mafft Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usa-bility. Mol. Biol. Evol. 2013, 4, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. Partition Finder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; St. John, O.; Wild, R.; Hammond, P.M.; Ahrens, D.; Balke, M.; Caterino, M.S.; et al. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef] [PubMed]
- Crowson, R.A. Observations on the constitution and subfamilies of the family Melandryidae (Coleoptera). Eos 1966, 41, 507–513. [Google Scholar]
- Lawrence, J.F.; Newton, A.F., Jr. Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names). In Biology, Phylogeny, and Classification of Coleoptera: Papers Celebrating the 80th Birthday of Roy A. Crowson; Pakaluk, J., Slipinski, S.A., Eds.; Museum i Instytut Zoologii PAN: Warsaw, Poland, 1995; pp. 779–1006. [Google Scholar]
- Csiki, E. Serropalpidae. In Coleopterorum Catalogus auspiciis et auxilio, Volume 17, Part 77; Schenkling, S., Ed.; W. Junk: Berlin, Germany, 1924; pp. 1–62. [Google Scholar]
- Arnett, R.H. The Beetles of United States (A Manual for Identification); Catholic University of America Press: Washington, DC, USA, 1963; pp. 1–1112. [Google Scholar]
- Gunter, N.L.; Levkaničová, Z.; Weir, T.H.; Ślipiński, A.; Cameron, S.L.; Bocak, L. Toward a phylogeny of the Tenebrionoidea (Coleoptera). Mol. Phylogenet. Evol. 2014, 79, 305–312. [Google Scholar] [CrossRef]
- Lawrence, J.F. Order Coleoptera: Tetratomidae; Melandryidae; Mordellidae (Tenebrionoidea). In Immature Insects, Volume 2; Stehr, F.W., Ed.; Springer: Dubuque, IA, USA, 1991; pp. 502–508. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superfamily | Family | Species | GenBank Accession No. | References |
|---|---|---|---|---|
| Tenebrionoidea | Aderidae | Aderidae sp. | JX412763.1 | [28] |
| (ingroups) | Anthicidae | Anthicidae sp. | MH789722.1 | [27] |
| Stricticollis tobias (Marseul, 1879) | KX087350.1 | [29] | ||
| Boridae | Boros schneideri (Panzer, 1795) | HQ232823.1 | [39] | |
| Ciidae | Ciidae sp. | JX412846.1 | [28] | |
| Melandryidae | Mikadonius gracilis Lewis, 1895 | JX412823.1 | [28] | |
| Osphya bipunctata (Fabricius, 1775) | JX313675.1 | [28] | ||
| Osphya sinensis sp. n. | MW556727 | This study | ||
| Meloidae | Epicauta gorhami (Marseul, 1873) | NC_036042.1 | [40] | |
| Hycleus marcipoli Pan & Bologna, 2014 | NC_036044.1 | [40] | ||
| Lytta caraganae (Pallas, 1798) | NC_033339.1 | Unpublished | ||
| Mylabris aulica Menetries, 1832 | NC_036046.1 | [40] | ||
| Mordellidae | Mordella atrata Melsheimer, 1845 | NC_013254.1 | [41] | |
| Mordellochroa milleri (Emery, 1876) | KX087318.1 | [29] | ||
| Tomoxia bucephala (Costa, A., 1854) | KX087355.1 | [29] | ||
| Mycetophagidae | Mycetophagus quadripustulatus (Linnaeus, 1761) | HQ232824.1 | [39] | |
| Mycteridae | Hemipeplus sp. | JX412852.1 | [28] | |
| Stilpnonotus mexicanus (Thomson, 1860) | JX412811.1 | [28] | ||
| Oedemeridae | Ischnomera caerulea (Linnaeus, 1758) | JX412790.1 | [28] | |
| Nacerdes carniolica (Gistl, 1832) | KX087319.1 | [29] | ||
| Prostomidae | Prostomis sp. | JX412787.1 | [28] | |
| Pyrochroidae | Schizotus pectinicornis (Linnaeus, 1758) | KX087342.1 | [29] | |
| Ripiphoridae | Pelecotoma fennica (Paykull, 1799) | NC_036277.1 | [29] | |
| Salpingidae | Lissodema cursor (Gyllenhal, 1813) | KX087307.1 | [29] | |
| Vincenzellus ruficollis (Panzer, 1794) | NC_036274.1 | [29] | ||
| Scraptiidae | Anaspis sp. | JX412856.1 | [28] | |
| Cyrtanaspis phalerata (Germar, 1831) | KX087279.1 | [29] | ||
| Scraptia sp. | JX412825.2 | [28] | ||
| Tenebrionidae | Alphitobius diaperinus (Panzer, 1797) | NC_049092.1 | [42] | |
| Asbolus verrucosus LeConte, 1851 | NC_027256.1 | [43] | ||
| Blaps rhynchoptera Fairmaire, 1886 | NC_047449.1 | [44] | ||
| Nalassus laevioctostriatus (Goeze, 1777) | KT876905.1 | [45] | ||
| Promethis valgipes (Marseul, 1876) | MW201671.1 | [46] | ||
| Scaphidema metallicum (Fabricius, 1792) | KX087341.1 | [29] | ||
| Strongylium suspicax (Kolbe, 1894) | JX412780.1 | [28] | ||
| Tenebrio molitor Linnaeus, 1758 | NC_024633.1 | [47] | ||
| Tribolium castaneum (Herbst, 1797) | NC_003081.2 | [48] | ||
| Ulomoides dermestoides (Fairmaire, 1893) | NC_025332.1 | [49] | ||
| Zophobas atratus (Fabricius, 1775) | NC_041101.1 | [50] | ||
| Tetratomidae | Tetratoma fungorum Fabricius, 1790 | NC_036276.1 | [29] | |
| Zopheridae | Usechus lacerta Motschulsky, 1854 | NC_036269.1 | [29] | |
| Cucujoidea | Nitidulidae | Carpophilus pilosellus Motschulsky, 1858 | NC_046035.1 | [51] |
| (outgroups) | Omosita colon (Linnaeus, 1758) | NC_050852.1 | Unpublished |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Yuan, L.; Wang, P.; Pan, Z.; Tong, J.; Wu, G.; Yang, Y. First Record of Osphya (Melandryidae: Osphyinae) from Chinese Mainland Based on Morphological Evidence and Mitochondrial Genome-Based Phylogeny of Tenebrionoidea. Diversity 2023, 15, 282. https://doi.org/10.3390/d15020282
Liu H, Yuan L, Wang P, Pan Z, Tong J, Wu G, Yang Y. First Record of Osphya (Melandryidae: Osphyinae) from Chinese Mainland Based on Morphological Evidence and Mitochondrial Genome-Based Phylogeny of Tenebrionoidea. Diversity. 2023; 15(2):282. https://doi.org/10.3390/d15020282
Chicago/Turabian StyleLiu, Haoyu, Lilan Yuan, Ping Wang, Zhao Pan, Junbo Tong, Gang Wu, and Yuxia Yang. 2023. "First Record of Osphya (Melandryidae: Osphyinae) from Chinese Mainland Based on Morphological Evidence and Mitochondrial Genome-Based Phylogeny of Tenebrionoidea" Diversity 15, no. 2: 282. https://doi.org/10.3390/d15020282