
Can Gap-Cutting Help to Preserve Forest Spider Communities?
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
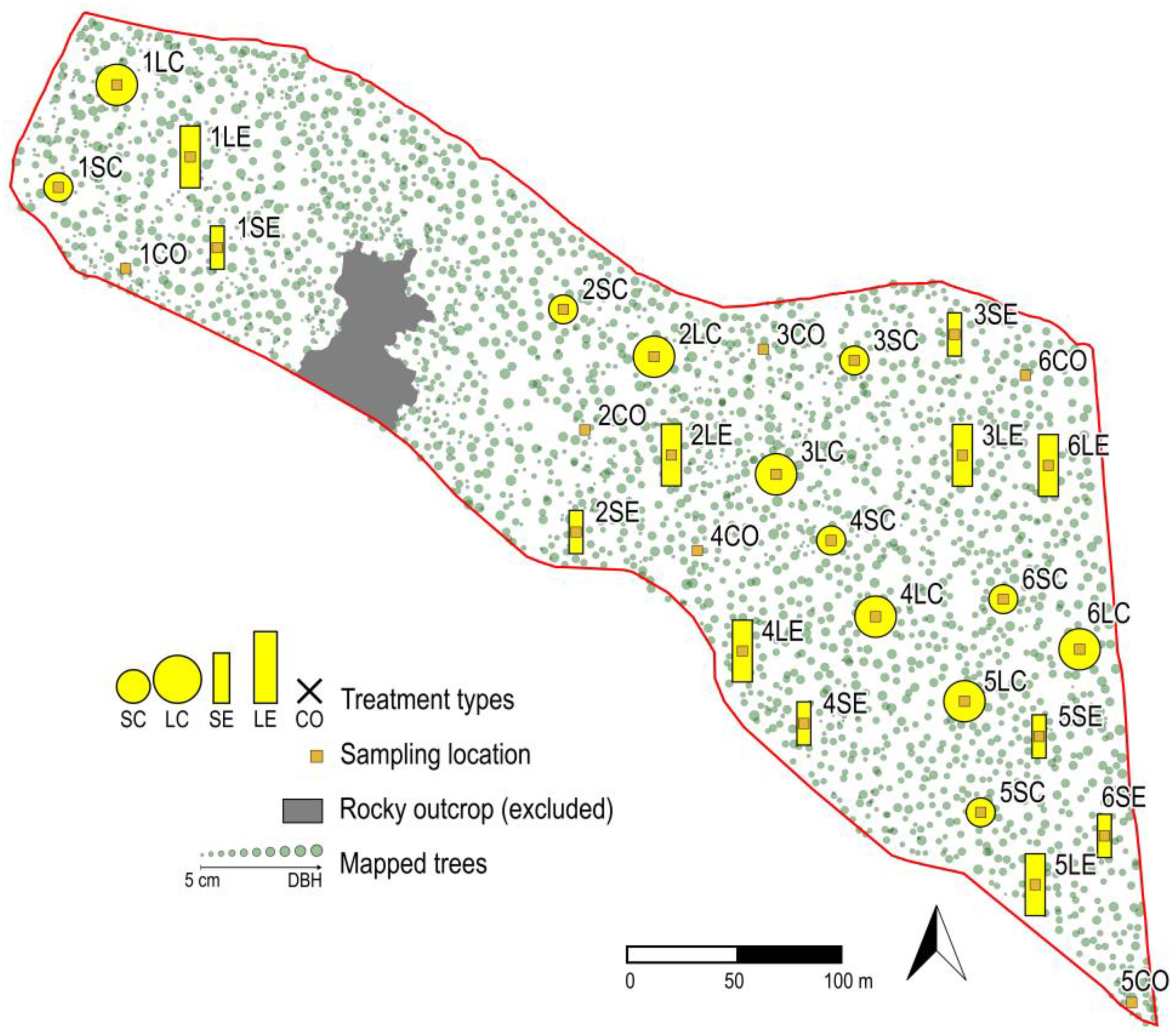
2.1. Study Area
2.2. Sampling Design and Data Collection
- Control (CO): mature, closed-canopy stand without any treatment;
- Large Circular (LC): a circular gap with a 10-m radius (i.e., total length of one mature tree) and the approximate size of 300 m2. This type may represent the most common approach for gap-cutting in the study area, although the managers achieve this size in several steps in time, and the shape might be slightly irregular due to spatial constraints;
- Small Circular (SC): a gap with a 7-m radius that corresponds to 2/3 of the length of a mature tree, with an area of 150 m2;
- Large Elongated (LE): a large gap of elongated type, also with N-S orientation (width: 10 m, length: 30 m) and in the same size as LC;
- Small Elongated (SE): a gap where trees were removed along a longitudinal axis (width: 7 m, length: 21 m) with N-S orientation to avoid the large extent of direct radiation and in the same size as SC.

2.3. Statistical Analysis
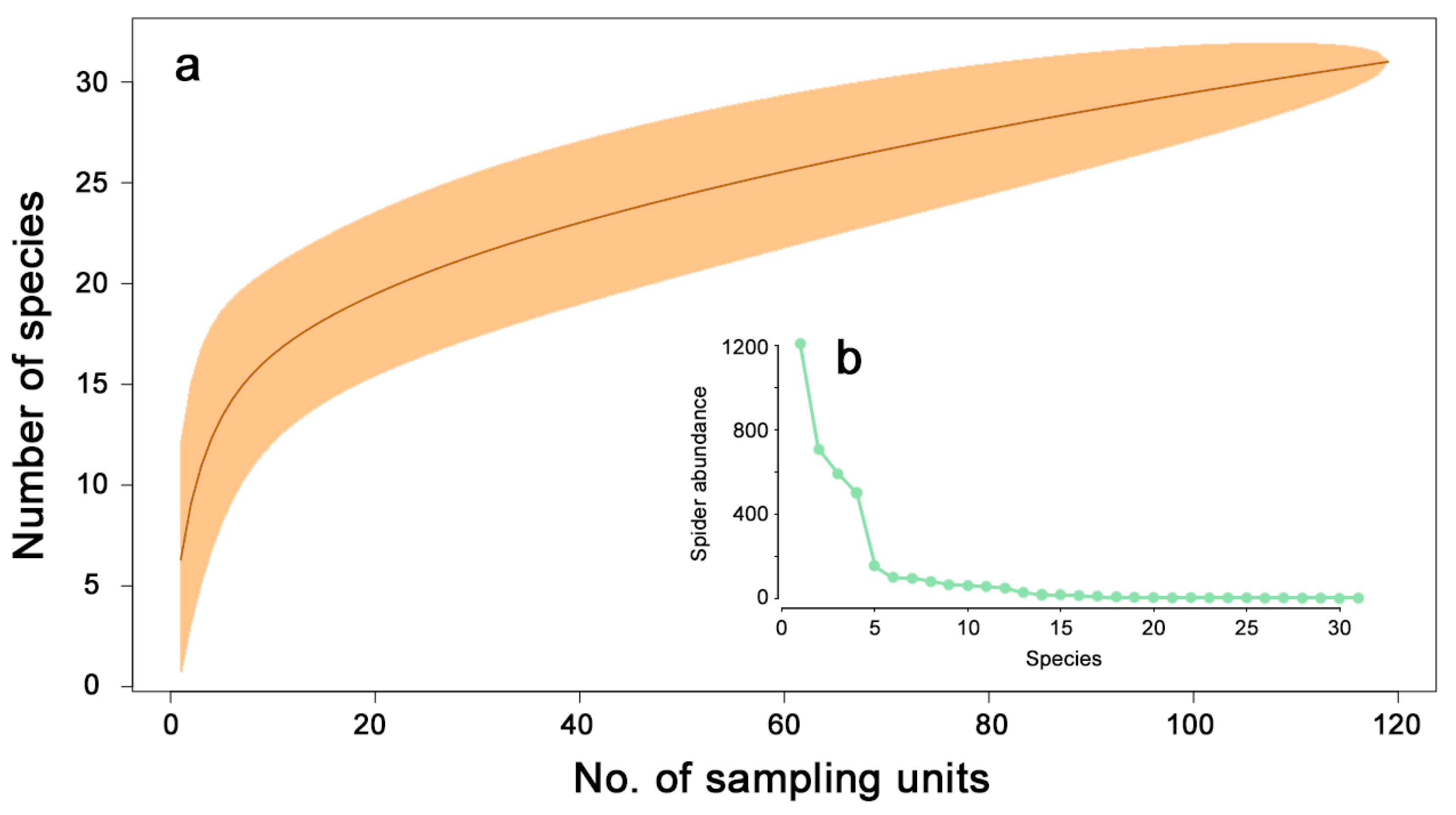
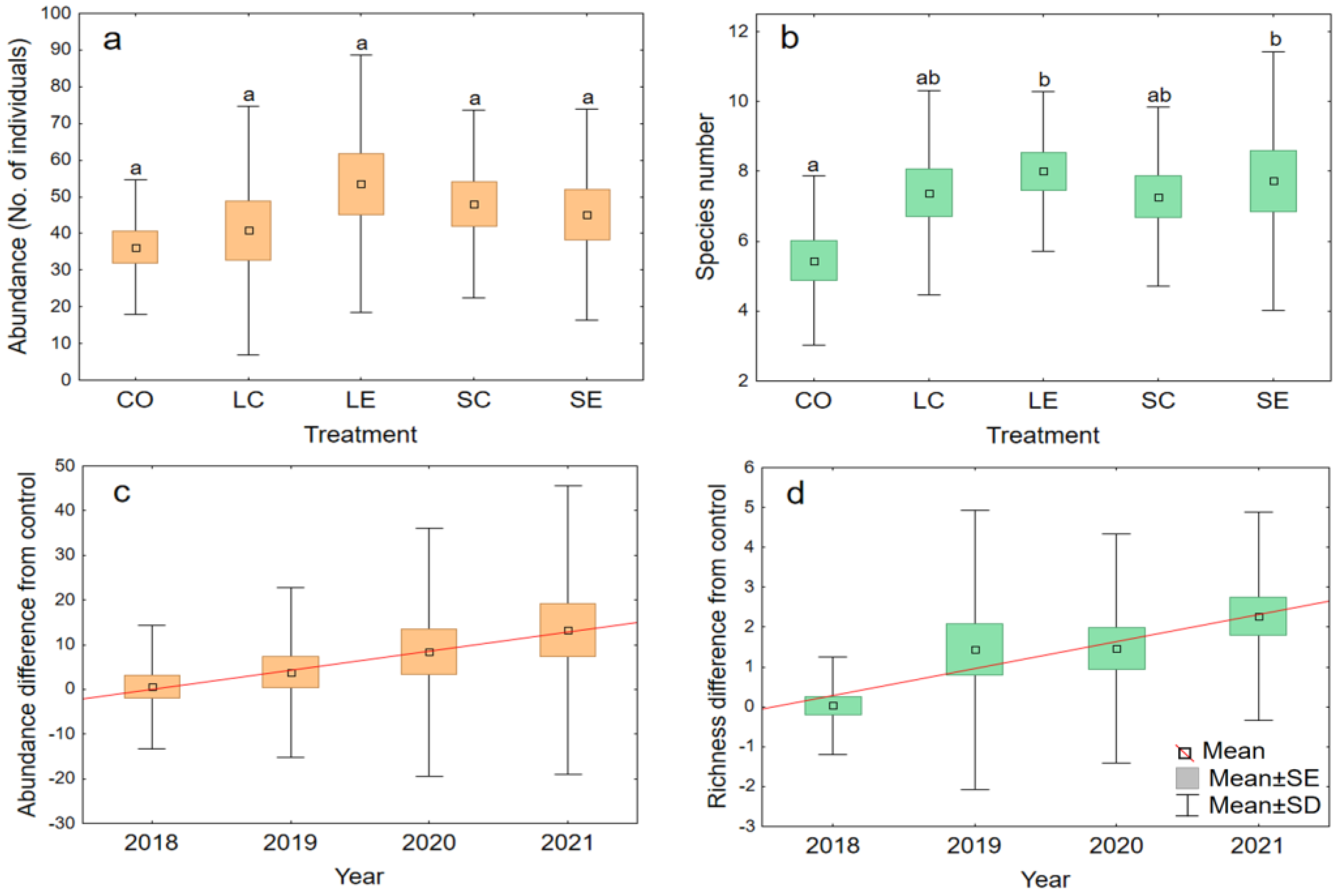
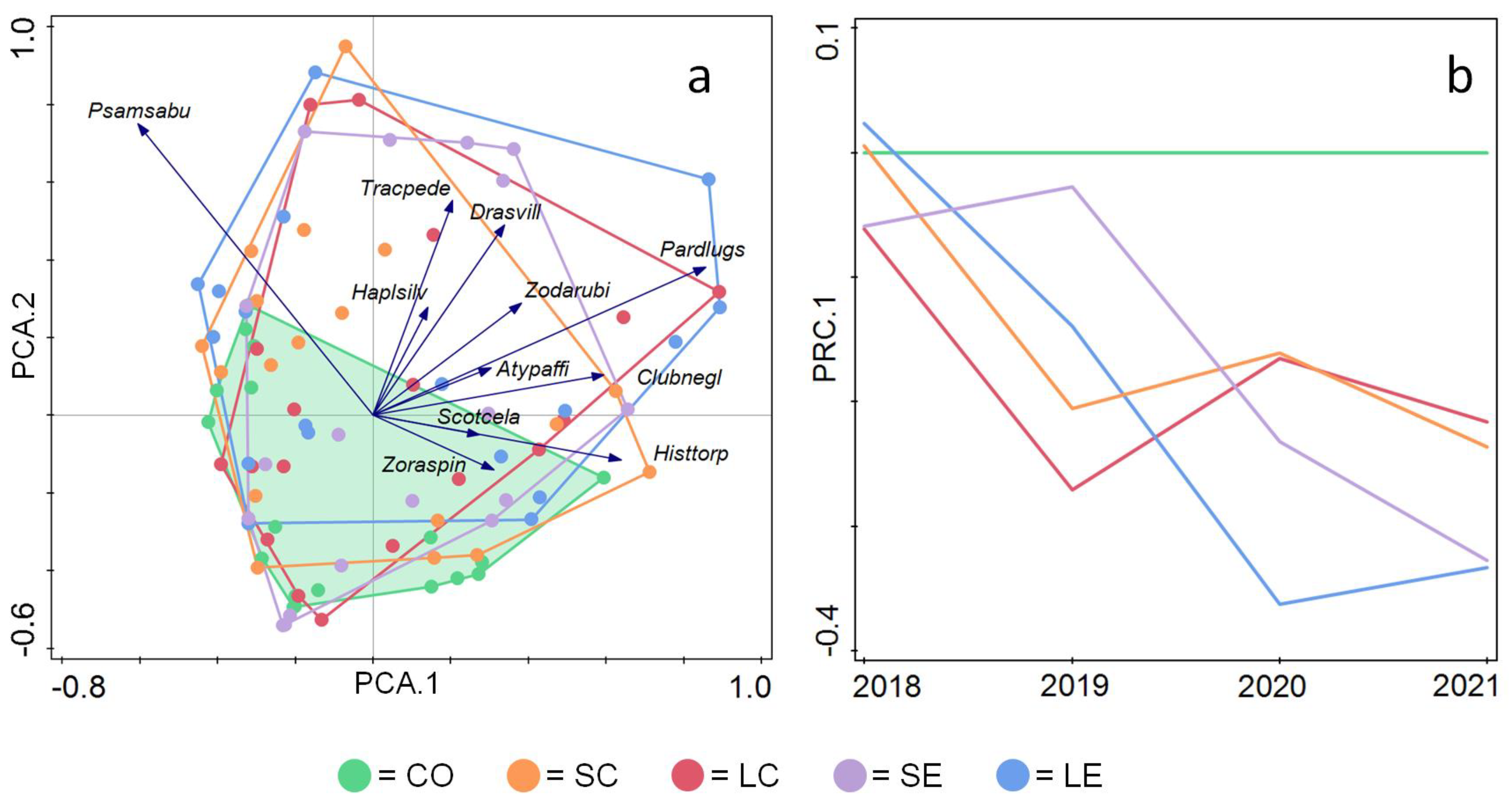
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species Name | Treatment | ||||
|---|---|---|---|---|---|---|
| CO | LC | LE | SC | SE | ||
| Agelenidae | Histopona torpida (C. L. Koch, 1834) | 16 | 13 | 27 | 25 | 14 |
| Agelenidae | Urocoras longispina (Kulczynski, 1897) | 324 | 166 | 204 | 280 | 233 |
| Araneidae | Cercidia prominens (Westring, 1851) | 1 | ||||
| Atypidae | Atypus affinis Eichwald, 1830 | 9 | 11 | 13 | 10 | 14 |
| Clubionidae | Clubiona neglecta O. P.-Cambridge, 1862 | 2 | 6 | 21 | 40 | 30 |
| Clubionidae | Clubiona terrestris Westring, 1851 | 1 | ||||
| Dysderidae | Dysdera erythrina (Walckenaer, 1802) | 10 | 6 | 19 | 2 | 10 |
| Dysderidae | Dysdera ninnii Canestrini, 1868 | 2 | 1 | 1 | 1 | 3 |
| Dysderidae | Harpactea rubicunda (C. L. Koch, 1838) | 15 | 5 | 3 | 4 | |
| Gnaphosidae | Drassyllus villicus (Thorell, 1875) | 4 | 38 | 49 | 30 | 33 |
| Gnaphosidae | Haplodrassus silvestris (Blackwall, 1833) | 8 | 12 | 18 | 12 | 12 |
| Gnaphosidae | Trachyzelotes pedestris (C. L. Koch, 1837) | 1 | 14 | 30 | 23 | 12 |
| Hahniidae | Cicurina cicur (Fabricius, 1793) | 1 | ||||
| Linyphiidae | Diplostyla concolor (Wider, 1834) | 1 | ||||
| Linyphiidae | Tapinopa longidens (Wider, 1834) | 1 | 1 | |||
| Linyphiidae | Tenuiphantes flavipes (Blackwall, 1854) | 1 | ||||
| Liocranidae | Agroeca brunnea (Blackwall, 1833) | 9 | 19 | 6 | 18 | 13 |
| Liocranidae | Liocranoeca striata (Kulczynski, 1882) | 1 | ||||
| Liocranidae | Scotina celans (Blackwall, 1841) | 1 | 3 | 4 | 4 | |
| Lycosidae | Aulonia albimana (Walckenaer, 1805) | 1 | 1 | |||
| Lycosidae | Pardosa lugubris s.str. (Walckenaer, 1802) | 27 | 203 | 183 | 73 | 107 |
| Lycosidae | Trochosa terricola Thorell, 1856 | 105 | 74 | 114 | 100 | 110 |
| Mimetidae | Ero furcata (Villers, 1789) | 1 | 1 | 1 | ||
| Miturgidae | Zora spinimana (Sundevall, 1833) | 1 | 5 | 7 | 3 | |
| Pisauridae | Pisaura mirabilis (Clerck, 1757) | 1 | 2 | |||
| Salticidae | Evarcha arcuata (Clerck, 1757) | 1 | 1 | |||
| Salticidae | Marpissa nivoyi (Lucas, 1846) | 1 | ||||
| Sparassidae | Micrommata virescens (Clerck, 1757) | 1 | ||||
| Tetragnathidae | Pachygnatha degeeri Sundevall, 1830 | 1 | ||||
| Thomisidae | Psammitis sabulosus (Hahn, 1832) | 115 | 118 | 192 | 145 | 137 |
| Zodariidae | Zodarion rubidum Simon, 1914 | 6 | 3 | 5 | 3 | |
| Total number of adult individuals | 650 | 700 | 891 | 780 | 749 | |
| Number of species | 17 | 23 | 19 | 19 | 23 | |
| Shannon diversity | 1.60 | 2.02 | 2.08 | 2.03 | 2.09 | |
References
- Kraus, D.; Krumm, F. Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; European Forest Institute: Freiburg, Germany, 2013. [Google Scholar]
- García-Nieto, A.P.; García-Llorente, M.; Iniesta-Arandia, I.; Martín-López, B. Mapping forest ecosystem services: From providing units to beneficiaries. Ecosyst. Serv. 2013, 4, 126–138. [Google Scholar] [CrossRef]
- Mori, A.S.; Lertzman, K.P.; Gustafsson, L.; Cadotte, M. Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology. J. Appl. Ecol. 2017, 54, 12–27. [Google Scholar] [CrossRef]
- Forest Europe. State of Europe’s Forests. In Proceedings of the Ministerial Conference on the Protection of Forests in Europe, Madrid, Spain, 20–21 October 2015. [Google Scholar]
- Pommerening, A.; Murphy, S.T. A review of the history, definitions and methods of continuous cover forestry with special attention to afforestation and restocking. Forestry 2004, 77, 27–44. [Google Scholar] [CrossRef]
- Peura, M.; Burgas, D.; Eyvindson, K.; Repo, A.; Mönkkönen, M. Continuous cover forestry is a cost-efficient tool to increase multifunctionality of boreal production forests in Fennoscandia. Biol. Conserv. 2018, 217, 104–112. [Google Scholar] [CrossRef]
- Aszalós, R.; Thom, D.; Aakala, T.; Angelstam, P.; Brūmelis, G.; Gálhidy, L.; Gratzer, G.; Hlásny, T.; Katzensteiner, K.; Kovács, B.; et al. Natural disturbance regimes as a guide for sustainable forest management in Europe. Ecol. Appl. 2022, 32, e2596. [Google Scholar] [CrossRef]
- Schall, P.; Gossner, M.M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; Jung, K.; Baumgartner, V.; Blaser, S.; Böhm, S.; et al. The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. J. Appl. Ecol. 2018, 55, 267–278. [Google Scholar] [CrossRef]
- Mason, W.L.; Diaci, J.; Carvalho, J.; Valkonen, S. Continuous cover forestry in Europe: Usage and the knowledge gaps and challenges to wider adoption. Forestry 2021, 95, 1–12. [Google Scholar] [CrossRef]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’Amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. Forestry 2017, 90, 4–17. [Google Scholar] [CrossRef]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Lhotka, J.M.; Cunningham, R.A.; Stringer, J.W. Effect of silvicultural gap size on 51 year species recruitment, growth and volume yields in Quercus dominated stands of the Northern Cumberland Plateau, USA. Forestry 2018, 91, 451–458. [Google Scholar] [CrossRef]
- Tobisch, T. Parent stand growth following gap and shelterwood cutting in a sessile oak-hornbeam forest. Acta Silvat. Lignar. Hung. 2010, 6, 33–48. [Google Scholar]
- Kovács, B.; Tinya, F.; Guba, E.; Németh, C.; Sass, V.; Bidló, A.; Ódor, P. The Short-Term Effects of Experimental Forestry Treatments on Site Conditions in an Oak–Hornbeam Forest. Forests 2018, 9, 406. [Google Scholar] [CrossRef]
- Kovács, B.; Tinya, F.; Németh, C.; Ódor, P. Unfolding the effects of different forestry treatments on microclimate in oak forests: Results of a 4-yr experiment. Ecol. Appl. 2020, 30, e02043. [Google Scholar] [CrossRef]
- Tinya, F.; Kovács, B.; Prättälä, A.; Farkas, P.; Aszalós, R.; Ódor, P. Initial understory response to experimental silvicultural treatments in a temperate oak-dominated forest. Eur. J. For. Res. 2018, 138, 65–77. [Google Scholar] [CrossRef]
- Elek, Z.; Kovács, B.; Aszalós, R.; Boros, G.; Samu, F.; Tinya, F.; Ódor, P. Taxon-specific responses to different forestry treatments in a temperate forest. Sci. Rep. 2018, 8, 16990. [Google Scholar] [CrossRef] [PubMed]
- Samu, F.; Elek, Z.; Kovács, B.; Fülöp, D.; Botos, E.; Schmera, D.; Aszalós, R.; Bidló, A.; Németh, C.; Sass, V.; et al. Resilience of spider communities affected by a range of silvicultural treatments in a temperate deciduous forest stand. Sci. Rep. 2021, 11, 20520. [Google Scholar] [CrossRef] [PubMed]
- Dövényi, Z. Magyarország Kistájainak katasztere [Cadastre of Hungarian Regions]; MTA Földrajztudományi Kutatóintézet: Budapest, Hungary, 2010. [Google Scholar]
- Scheiner, S.M.; Gurevitch, J. Design and Analysis of Ecological Experiments, 2nd ed.; Oxford University Press: New York, NY, USA, 2001; p. 415. [Google Scholar]
- Sapia, M.; Lövei, G.L.; Elek, Z. Effects of varying sampling effort on the observed diversity of carabid (Coleoptera: Carabidae) assemblages in the Danglobe Project, Denmark. Entomol. Fenn. 2006, 17, 345–350. [Google Scholar] [CrossRef]
- Jimenez-Valverde, A.; Lobo, J.M. Establishing reliable spider (Araneae, Araneidae and Thomisidae) assemblage sampling protocols: Estimation of species richness, seasonal coverage and contribution of juvenile data to species richness and composition. Acta Oecol. 2006, 30, 21–32. [Google Scholar] [CrossRef]
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Kropf, C. Spiders of Europe. Version 07. 2022. Available online: https://www.araneae.nmbe.ch (accessed on 6 January 2023).
- World Spider Catalog. World Spider Catalog; Version 23.5; Natural History Museum Bern: Bern, Switzerland, 2022; Available online: http://wsc.nmbe.ch (accessed on 6 January 2023).
- Saska, P.; van der Werf, W.; Hemerik, L.; Luff, M.L.; Hatten, T.D.; Honek, A. Temperature effects on pitfall catches of epigeal arthropods: A model and method for bias correction. J. Appl. Ecol. 2013, 50, 181–189. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Simpson, G.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package; R package version 2.6-4. 2022. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 6 January 2023).
- R Core Team. R: A Language and Environment for Statistical Computing, version 4.2.2; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org (accessed on 6 January 2023).
- Whittaker, R.H. Dominance and diversity in land plant communities. Science 1965, 147, 250–260. [Google Scholar] [PubMed]
- SAS Institute. JMP Statistics and Graphics Guide, Release 6; SAS Institute Inc.: Cary, NC, USA, 2005. [Google Scholar]
- Smilauer, P.; Leps, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; p. 282. [Google Scholar]
- ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012; p. 496. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge: Oxford, UK, 2013. [Google Scholar]
- Van den Brink, P.J.; Braak, C.J.F.T. Principal response curves: Analysis of time-dependent multivariate responses of biological community to stress. Environ. Toxicol. Chem. 1999, 18, 138–148. [Google Scholar] [CrossRef]
- Boros, G.; Kovács, B.; Ódor, P. Green tree retention enhances negative short-term effects of clear-cutting on enchytraeid assemblages in a temperate forest. Appl. Soil Ecol. 2019, 136, 106–115. [Google Scholar] [CrossRef]
- Beck, J.; Pfiffner, L.; Ballesteros-Mejia, L.; Blick, T.; Luka, H. Revisiting the indicator problem: Can three epigean arthropod taxa inform about each other’s biodiversity? Divers. Distrib. 2013, 19, 688–699. [Google Scholar] [CrossRef]
- Smith, G.F.; Gittings, T.; Wilson, M.; French, L.; Oxbrough, A.; O’Donoghue, S.; O’Halloran, J.; Kelly, D.L.; Mitchell, F.J.G.; Kelly, T.; et al. Identifying practical indicators of biodiversity for stand-level management of plantation forests. Biodivers. Conserv. 2008, 17, 991–1015. [Google Scholar] [CrossRef]
- Pearce, J.L.; Venier, L.A. The use of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) as bioindicators of sustainable forest management: A review. Ecol. Indic. 2006, 6, 780–793. [Google Scholar] [CrossRef]
- Connell, J.H. Intermediate-disturbance hypothesis. Science 1979, 204, 1344–1345. [Google Scholar]
- Mackey, R.L.; Currie, D.J. The Diversity–Disturbance Relationship: Is It Generally Strong and Peaked? Ecology 2001, 82, 3479–3492. [Google Scholar] [CrossRef]
- Hughes, A.R.; Byrnes, J.E.; Kimbro, D.L.; Stachowicz, J.J. Reciprocal relationships and potential feedbacks between biodiversity and disturbance. Ecol. Lett. 2007, 10, 849–864. [Google Scholar] [CrossRef]
- Pinzon, J.; Spence, J.R.; Langor, D.W. Responses of ground-dwelling spiders (Araneae) to variable retention harvesting practices in the boreal forest. For. Ecol. Manag. 2012, 266, 42–53. [Google Scholar] [CrossRef]
- Pinzon, J.; Spence, J.R.; Langor, D.W. Effects of prescribed burning and harvesting on ground-dwelling spiders in the Canadian boreal mixedwood forest. Biodivers. Conserv. 2013, 22, 1513–1536. [Google Scholar] [CrossRef]
- Munevar, A.; Rubio, G.D.; Zurita, G.A. Changes in spider diversity through the growth cycle of pine plantations in the semi-deciduous Atlantic forest: The role of prey availability and abiotic conditions. For. Ecol. Manag. 2018, 424, 536–544. [Google Scholar] [CrossRef]
- Welch, K.D.; Haynes, K.F.; Harwood, J.D. Microhabitat evaluation and utilization by a foraging predator. Anim. Behav. 2013, 85, 419–425. [Google Scholar] [CrossRef]
- Samu, F.; Kadar, F.; Onodi, G.; Kertesz, M.; Sziranyi, A.; Szita, E.; Fetyko, K.; Neidert, D.; Botos, E.; Altbacker, V. Differential ecological responses of two generalist arthropod groups, spiders and carabid beetles (Araneae, Carabidae), to the effects of wildfire. Community Ecol. 2010, 11, 129–139. [Google Scholar] [CrossRef]
- Uetz, G.W.; Unzicker, J.D. Pitfall trapping in ecological studies of wandering spiders. J. Arachnol. 1976, 3, 101–111. [Google Scholar]
- Lang, A. The pitfalls of pitfalls: A comparison of pitfall trap catches and absolute density estimates of epigeal invertebrate predators in arable land. J. Pest Sci. 2000, 73, 99–106. [Google Scholar]
- Topping, C.J.; Sunderland, K.D. Limitations to the use of pitfall traps in ecological studies, exemplified by a study of spiders in a field of winter wheat. J. Appl. Ecol. 1992, 29, 485–491. [Google Scholar] [CrossRef]
- Buddle, C.M.; Higgins, S.; Rypstra, A.L. Ground-dwelling spider assemblages inhabiting riparian forests and hedgerows in an agricultural landscape. Am. Midl. Nat. 2004, 151, 15–26. [Google Scholar]
- McIver, J.D.; Parsons, G.L.; Moldenke, A.R. Litter spider succession after clear-cutting in a western coniferous forest. Can. J. For. Res. 1992, 22, 984–992. [Google Scholar] [CrossRef]
- Bali, L.; Andrési, D.; Tuba, K.; Szinetár Csaba, M. Comparing pitfall trapping and suction sampling data collection for ground-dwelling spiders in artificial forest gaps. Arachnol. Mitt. 2019, 58, 23–28. [Google Scholar]
- Samu, F.; Lengyel, G.; Szita, É.; Bidló, A.; Ódor, P. The effect of forest stand characteristics on spider diversity and species composition in deciduous-coniferous mixed forests. J. Arachnol. 2014, 42, 135–141. [Google Scholar] [CrossRef]
- Siewers, J.; Schirmel, J.; Buchholz, S. The efficiency of pitfall traps as a method of sampling epigeal arthropods in litter rich forest habitats. Eur. J. Entomol. 2014, 111, 69–74. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. The gap theory in forest dynamics. Bot. Mag. 1992, 105, 375–383. [Google Scholar] [CrossRef]
- Tinya, F.; Kovács, B.; Bidló, A.; Dima, B.; Király, I.; Kutszegi, G.; Lakatos, F.; Mag, Z.; Márialigeti, S.; Nascimbene, J.; et al. Environmental drivers of forest biodiversity in temperate mixed forests–A multi-taxon approach. Sci. Total Environ. 2021, 795, 148720. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Burivalova, Z.; Koh, L.P.; Hellweg, S. Impact of Forest Management on Species Richness: Global Meta-Analysis and Economic Trade-Offs. Sci. Rep. 2016, 6, 23954. [Google Scholar] [CrossRef] [Green Version]
- Cernecka, L.; Mihal, I.; Gajdos, P.; Jarcuska, B. The effect of canopy openness of European beech (Fagus sylvatica) forests on ground-dwelling spider communities. Insect Conserv. Divers. 2020, 13, 250–261. [Google Scholar] [CrossRef]
- Oxbrough, A.G.; Gittings, T.; O’Halloran, J.; Giller, P.S.; Smith, G.F. Structural indicators of spider communities across the forest plantation cycle. For. Ecol. Manag. 2005, 212, 171–183. [Google Scholar] [CrossRef]
- Galle, R.; Szabo, A.; Csaszar, P.; Torma, A. Spider assemblage structure and functional diversity patterns of natural forest steppes and exotic forest plantations. For. Ecol. Manag. 2018, 411, 234–239. [Google Scholar] [CrossRef]
- Andresi, D.; Bali, L.; Tuba, K.; Szinetar, C. Comparative study of ground beetle and ground-dwelling spider assemblages of artificial gap openings. Community Ecol. 2018, 19, 133–140. [Google Scholar] [CrossRef]
- Dorow, W.H.O.; Blick, T.; Pauls, S.U.; Schneider, A. Waldbindung ausgewählter Tiergruppen Deutschlands; BfN-Skripten 544: Bonn, Germany, 2019; p. 388. [Google Scholar]
- Biteniekyté, M.; Relys, V. Epigeic spider communities of a peat bog and adjacent habitats. Rev. Iber. Aracnol. 2008, 15, 81–87. [Google Scholar]
- Samu, F.; Horváth, A.; Neidert, D.; Botos, E.; Szita, É. Metacommunities of spiders in grassland habitat fragments of an agricultural landscape. Basic Appl. Ecol. 2018, 31, 92–103. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samu, F.; Elek, Z.; Růžičková, J.; Botos, E.; Kovács, B.; Ódor, P. Can Gap-Cutting Help to Preserve Forest Spider Communities? Diversity 2023, 15, 240. https://doi.org/10.3390/d15020240
Samu F, Elek Z, Růžičková J, Botos E, Kovács B, Ódor P. Can Gap-Cutting Help to Preserve Forest Spider Communities? Diversity. 2023; 15(2):240. https://doi.org/10.3390/d15020240
Chicago/Turabian StyleSamu, Ferenc, Zoltán Elek, Jana Růžičková, Erika Botos, Bence Kovács, and Péter Ódor. 2023. "Can Gap-Cutting Help to Preserve Forest Spider Communities?" Diversity 15, no. 2: 240. https://doi.org/10.3390/d15020240