
PFOS Induces Lipometabolism Change, Immune Defense, and Endocrine Disorders in Black-Spotted Frogs: Application of Transcriptome Profiling

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Exposure Strategy
2.4. RNA Extraction, cDNA Library Construction, and Sequencing
2.5. Transcriptomic Assembly and Gene Annotation
2.6. Analysis of Differentially Expressed Genes (DEGs)
3. Results
3.1. Transcriptome Sequencing and Assembly
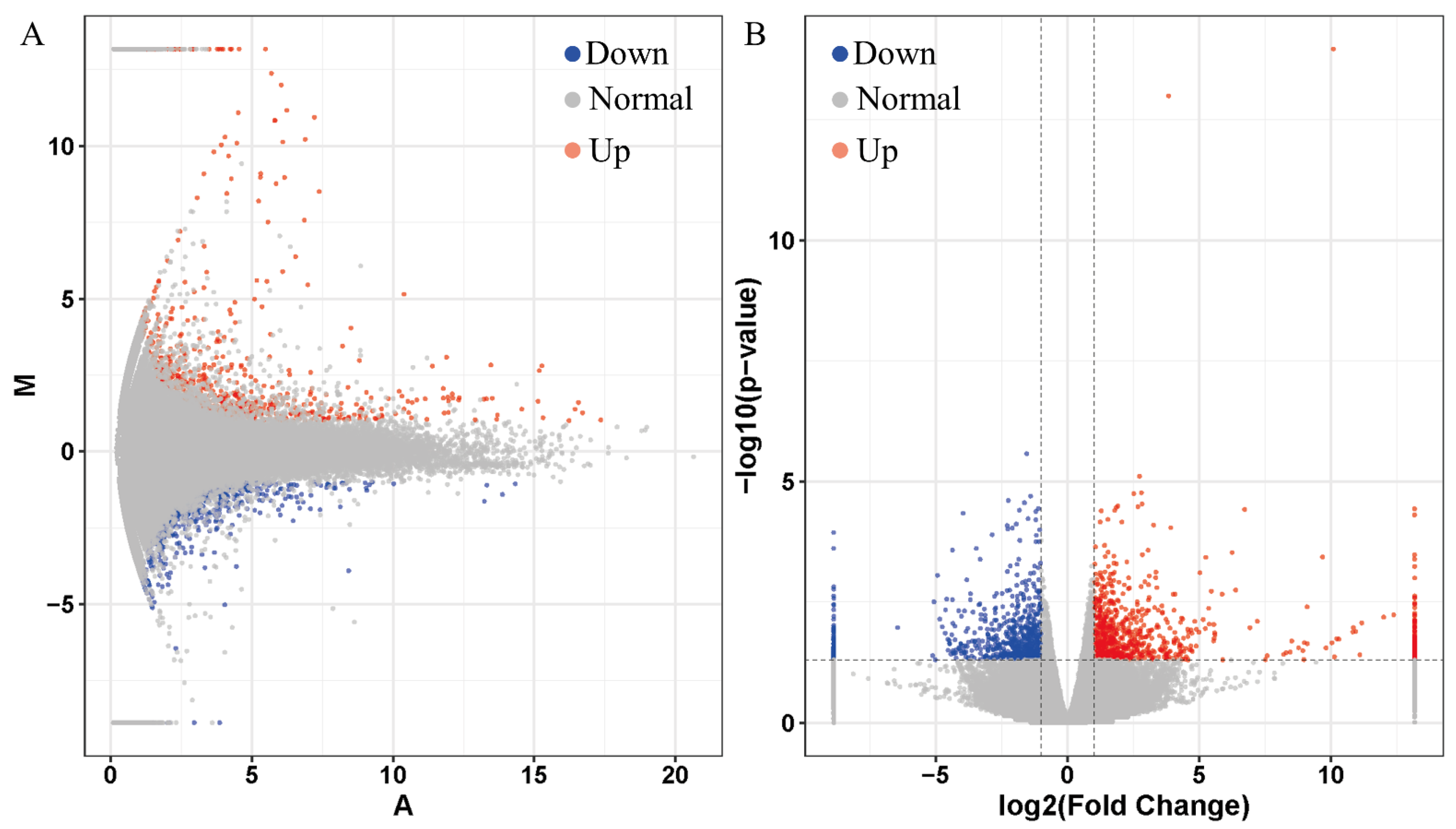
3.2. Identified of Differentially Expressed Genes (DEGs)
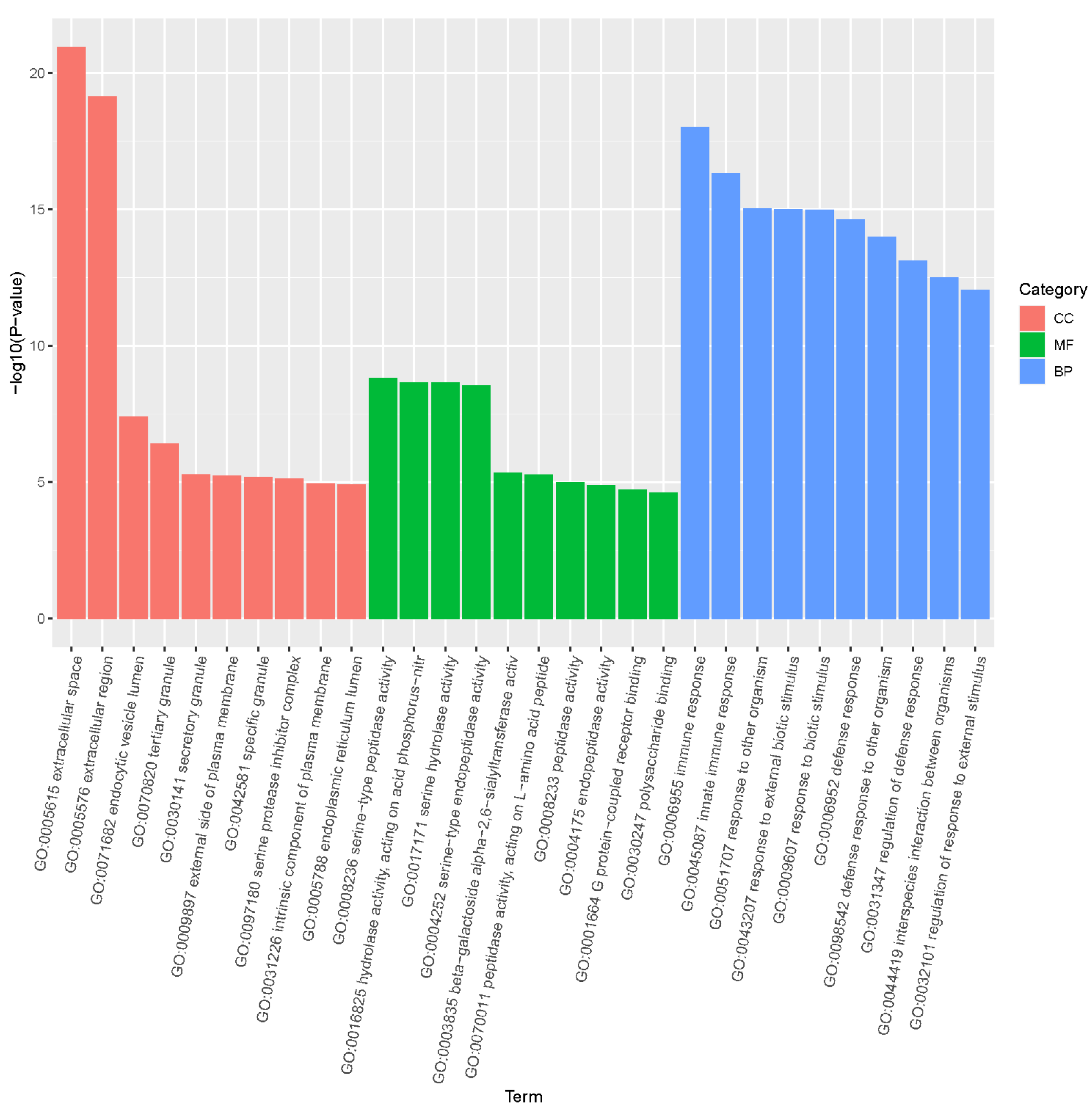
3.3. GO Analysis
3.4. KEGG Enrichment
4. Discussion
4.1. PPAR Signaling Pathway
4.2. Lipid Metabolism
4.3. Immune Defense
4.4. Endocrine System
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Blaustein, A.R.; Romansic, J.M.; Kiesecker, J.M.; Hatch, A.C. Ultraviolet radiation, toxic chemicals and amphibian population declines. Divers. Distrib. 2003, 9, 123–140. [Google Scholar] [CrossRef]
- Mann, R.M.; Hyne, R.V.; Choung, C.B.; Wilson, S.P. Amphibians and agricultural chemicals: Review of the risks in a complex environment. Environ. Pollut. 2009, 157, 2903–2927. [Google Scholar] [CrossRef] [PubMed]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-1. In IUCN Red List of Threatened Species (2020); IUCN: Gland, Switzerland, 2020; Available online: https://www.iucnredlist.org/ (accessed on 1 October 2020).
- Jiang, Z.; Jiang, J.; Wang, Y.; Zhang, E.; Zhang, Y.; Li, L.; Xie, F.; Cai, B.; Cao, L.; Zheng, G.; et al. Red list of china’s vertebrates. Biodivers. Sci. 2016, 24, 500–551. [Google Scholar]
- Zipkin, E.; DiRenzo, G.; Ray, J.; Rossman, S.; Lips, K. Tropical snake diversity collapses after widespread amphibian loss. Science 2020, 367, 814–816. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Garner, T.W. Chytrid fungi and global amphibian declines. Nat. Rev. Microbiol. 2020, 18, 332–343. [Google Scholar] [CrossRef]
- McCrink-Goode, M. Pollution: A global threat. Environ. Int. 2014, 68, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Orton, F.; Tyler, C. Do hormone-modulating chemicals impact on reproduction and development of wild amphibians? Biol. Rev. 2015, 90, 1100–1117. [Google Scholar] [CrossRef]
- Schellenberger, S.; Liagkouridis, I.; Awad, R.; Khan, S.; Plassmann, M.; Peters, G.; Benskin, J.P.; Cousins, I.T. An outdoor aging study to investigate the release of per- and polyfluoroalkyl substances (pfas) from functional textiles. Environ. Sci. Technol. 2022, 56, 3471–3479. [Google Scholar] [CrossRef]
- Hamid, H.; Li, L.Y.; Grace, J.R. Review of the fate and transformation of per- and polyfluoroalkyl substances (pfass) in landfills. Environ. Pollut. 2018, 235, 74–84. [Google Scholar] [CrossRef]
- Kaltenberg, E.M.; Dasu, K.; Lefkovitz, L.F.; Thorn, J.; Schumitz, D. Sampling of freely dissolved per- and polyfluoroalkyl substances (pfas) in surface water and groundwater using a newly developed passive sampler. Environ. Pollut. 2023, 318, 120940. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, J.; Guo, H.; Sheng, N.; Guo, Y.; Dai, J. Comparative hepatotoxicity of a novel perfluoroalkyl ether sulfonic acid, nafion byproduct 2 (h-pfmo2osa), and legacy perfluorooctane sulfonate (pfos) in adult male mice. Environ. Sci. Technol. 2022, 56, 10183–10192. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Liu, Z.; Yang, H.; Lu, L.; Chen, R.; Zhang, X.; Zhong, Y.; Zhang, H. Per- and polyfluoroalkyl substances (pfass) impair lipid metabolism in Rana nigromaculata: A field investigation and laboratory study. Environ. Sci. Technol. 2022, 56, 13222–13232. [Google Scholar] [CrossRef]
- Cui, Q.; Pan, Y.; Zhang, H.; Sheng, N.; Wang, J.; Guo, Y.; Dai, J. Occurrence and tissue distribution of novel perfluoroether carboxylic and sulfonic acids and legacy per/polyfluoroalkyl substances in black-spotted frog (Pelophylax nigromaculatus). Environ. Sci. Technol. 2018, 52, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wu, H.; Liu, F.; Yang, H.; Shen, L.; Chen, J.; Zhang, X.; Zhong, Y.; Zhang, H.; Liu, Z. Assessing the hepatotoxicity of pfoa, pfos, and 6:2 cl-pfesa in black-spotted frogs (Rana nigromaculata) and elucidating potential association with gut microbiota. Environ. Pollut. 2022, 312, 120029. [Google Scholar] [CrossRef]
- Zhou, Z.; Liang, Y.; Shi, Y.; Xu, L.; Cai, Y. Occurrence and transport of perfluoroalkyl acids (pfaas), including short-chain pfaas in tangxun lake, china. Environ. Sci. Technol. 2013, 47, 9249–9257. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Malinowski, C.R.; Sepúlveda, M.S. Emerging trends in nanoparticle toxicity and the significance of using daphnia as a model organism. Chemosphere 2021, 291, 132941. [Google Scholar] [CrossRef]
- Lin, H.; Feng, Y.; Zheng, Y.; Han, Y.; Yuan, X.; Gao, P.; Zhang, H.; Zhong, Y.; Liu, Z. Transcriptomic analysis reveals the hepatotoxicity of perfluorooctanoic acid in black-spotted frogs (Rana nigromaculata). Diversity 2022, 14, 971. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Z.; Tang, S.; Li, D.; Jiang, Q.; Zhang, T. Transcriptional response provides insights into the effect of chronic polystyrene nanoplastic exposure on daphnia pulex. Chemosphere 2020, 238, 124563. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Pérez, E.; Jiang, Q.; Chen, Q.; Jiao, Y.; Huang, Y.; Yang, Y.; Zhao, Y. Polystyrene nanoplastic induces oxidative stress, immune defense, and glycometabolism change in daphnia pulex: Application of transcriptome profiling in risk assessment of nanoplastics. J. Hazard. Mater. 2021, 402, 123778. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Sepúlveda, M.S.; Jiang, Q.; Jiao, Y.; Chen, Q.; Huang, Y.; Tian, J.; Zhao, Y. Development of an adverse outcome pathway for nanoplastic toxicity in daphnia pulex using proteomics. Sci. Total Environ. 2021, 766, 144249. [Google Scholar] [CrossRef]
- Davidsen, N.; Ramhøj, L.; Lykkebo, C.A.; Kugathas, I.; Poulsen, R.; Rosenmai, A.K.; Evrard, B.; Darde, T.A.; Axelstad, M.; Bahl, M.I.; et al. Pfos-induced thyroid hormone system disrupted rats display organ-specific changes in their transcriptomes. Environ. Pollut. 2022, 305, 119340. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Jorquera, I.A.; Colli-Dula, R.C.; Kroll, K.; Jayasinghe, B.S.; Parachu Marco, M.V.; Silva-Sanchez, C.; Toor, G.S.; Denslow, N.D. Blood transcriptomics analysis of fish exposed to perfluoro alkyls substances: Assessment of a non-lethal sampling technique for advancing aquatic toxicology research. Environ. Sci. Technol. 2018, 53, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Minhós, T.; Ferreira da Silva, M.; Bersacola, E.; Galat, G.; Galat-Luong, A.; Mayhew, M.; Starin, E. International Union for Conservation of Nature (IUCN) Red List Assessment, Temminck’s Red Colobus (Piliocolobus badius temminckii); e.T18247A92648587; IUCN: Gland, Switzerland, 2020. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Trinity: Reconstructing a full-length transcriptome without a genome from rna-seq data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using diamond. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. Kegg: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: Hmmer3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. Edger: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Kolde, R.; Kolde, M.R. Package ‘pheatmap’. R Package. 2018. Available online: http://www2.uaem.mx/r-mirror/web/packages/pheatmap/pheatmap.pdf (accessed on 1 October 2022).
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T. Kegg for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef]
- Mandal, P.K. Dioxin: A review of its environmental effects and its aryl hydrocarbon receptor biology. J. Comp. Physiol. B 2005, 175, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Barak, Y.; Nagy, L.; Liao, D.; Tontonoz, P.; Evans, R.M. Ppar-γ dependent and independent effects on macrophage-gene expression in lipid metabolism and inflammation. Nat. Med. 2001, 7, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.C.; Farese, R.V., Jr. Lipid droplets and cellular lipid metabolism. Annu. Rev. Biochem. 2012, 81, 687. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Lv, W.W.; Huang, Y.H.; Fan, B.; Li, Y.M.; Zhao, Y.L. Effects of cadmium on lipid metabolism in female estuarine crab, chiromantes dehaani. Comp. Biochem. Physiol. C-Toxicol. Pharmacol. 2016, 188, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lv, W.; Jiao, Y.; Liu, Z.; Li, Y.; Cai, M.; Wu, D.; Zhou, W.; Zhao, Y. Effects of exposure to waterborne polystyrene microspheres on lipid metabolism in the hepatopancreas of juvenile redclaw crayfish, cherax quadricarinatus. Aquat. Toxicol. 2020, 224, 105497. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Le Grand, F.; Bideau, A.; Huvet, A.; Paul-Pont, I.; Soudant, P. Nanoplastics exposure modulate lipid and pigment compositions in diatoms. Environ. Pollut. 2020, 262, 114274. [Google Scholar] [CrossRef]
- Lee, M.-C.; Park, J.C.; Lee, J.-S. Effects of environmental stressors on lipid metabolism in aquatic invertebrates. Aquat. Toxicol. 2018, 200, 83–92. [Google Scholar] [CrossRef]
- Zeng, Z.; Song, B.; Xiao, R.; Zeng, G.; Gong, J.; Chen, M.; Xu, P.; Zhang, P.; Shen, M.; Yi, H. Assessing the human health risks of perfluorooctane sulfonate by in vivo and in vitro studies. Environ. Int. 2019, 126, 598–610. [Google Scholar] [CrossRef]
- Liang, L.; Pan, Y.; Bin, L.; Liu, Y.; Huang, W.; Li, R.; Lai, K.P. Immunotoxicity mechanisms of perfluorinated compounds pfoa and pfos. Chemosphere 2022, 291, 132892. [Google Scholar] [CrossRef]
- Liu, Z.; Lin, H.; Zheng, Y.; Feng, Y.; Shi, C.; Zhu, R.; Shen, X.; Han, Y.; Zhang, H.; Zhong, Y. Perfluorooctanoic acid and perfluorooctanesulfonic acid induce immunotoxicity through the nf-κb pathway in black-spotted frog (Rana nigromaculata). Chemosphere 2023, 313, 137622. [Google Scholar] [CrossRef]
- Han, X.; Alam, M.N.; Cao, M.; Wang, X.; Cen, M.; Tian, M.; Lu, Y.; Huang, Q. Low levels of perfluorooctanoic acid exposure activates steroid hormone biosynthesis through repressing histone methylation in rats. Environ. Sci. Technol. 2022, 56, 5664–5672. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Reads No. | Bases (bp) | Q30 (bp) | N (%) | Q20 (%) | Q30 (%) | Clean Reads No. | Clean Data (bp) | Clean Reads % | Clean Data % |
|---|---|---|---|---|---|---|---|---|---|---|
| C1 | 48,625,956 | 7,293,893,400 | 6,792,623,194 | 0.009645 | 97.43 | 93.12 | 44,947,858 | 6,742,178,700 | 92.43 | 92.43 |
| C2 | 36,803,570 | 5,520,535,500 | 5,107,266,173 | 0.00963 | 97.15 | 92.51 | 33,650,206 | 5,047,530,900 | 91.43 | 91.43 |
| C3 | 45,578,514 | 6,836,777,100 | 6,372,523,669 | 0.009515 | 97.43 | 93.2 | 40,731,048 | 6,109,657,200 | 89.36 | 89.36 |
| C4 | 48,676,036 | 7,301,405,400 | 6,780,579,574 | 0.009638 | 97.27 | 92.86 | 44,445,616 | 6,666,842,400 | 91.3 | 91.3 |
| C5 | 38,909,934 | 5,836,490,100 | 5,422,004,285 | 0.009612 | 97.29 | 92.89 | 35,623,160 | 5,343,474,000 | 91.55 | 91.55 |
| P1 | 39,576,224 | 5,936,433,600 | 5,495,635,542 | 0.009637 | 97.14 | 92.57 | 36,199,482 | 5,429,922,300 | 91.46 | 91.46 |
| P2 | 44,026,002 | 6,603,900,300 | 6,133,830,390 | 0.009438 | 97.29 | 92.88 | 40,301,200 | 6,045,180,000 | 91.53 | 91.53 |
| P3 | 44,797,236 | 6,719,585,400 | 6,231,277,140 | 0.009436 | 97.21 | 92.73 | 40,892,672 | 6,133,900,800 | 91.28 | 91.28 |
| P4 | 37,467,758 | 5,620,163,700 | 5,198,500,671 | 0.009623 | 97.11 | 92.49 | 34,366,456 | 5,154,968,400 | 91.72 | 91.72 |
| P5 | 38,538,978 | 5,780,846,700 | 5,355,863,388 | 0.009672 | 97.14 | 92.64 | 35,310,926 | 5,296,638,900 | 91.62 | 91.62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, C.; Yang, H.; Xu, M.; Hua, T.; He, M.; Yang, Y.; Hou, X.; Zhang, H.; Liu, Z. PFOS Induces Lipometabolism Change, Immune Defense, and Endocrine Disorders in Black-Spotted Frogs: Application of Transcriptome Profiling. Diversity 2023, 15, 196. https://doi.org/10.3390/d15020196
Shi C, Yang H, Xu M, Hua T, He M, Yang Y, Hou X, Zhang H, Liu Z. PFOS Induces Lipometabolism Change, Immune Defense, and Endocrine Disorders in Black-Spotted Frogs: Application of Transcriptome Profiling. Diversity. 2023; 15(2):196. https://doi.org/10.3390/d15020196
Chicago/Turabian StyleShi, Chaoli, Hongmei Yang, Mengxuan Xu, Tianyang Hua, Mengfan He, Yuchen Yang, Xiaoyu Hou, Hangjun Zhang, and Zhiquan Liu. 2023. "PFOS Induces Lipometabolism Change, Immune Defense, and Endocrine Disorders in Black-Spotted Frogs: Application of Transcriptome Profiling" Diversity 15, no. 2: 196. https://doi.org/10.3390/d15020196