Fish Use of a Borland-Type Fish Lock in an Iberian River


Abstract
:1. Introduction
2. Materials and Methods
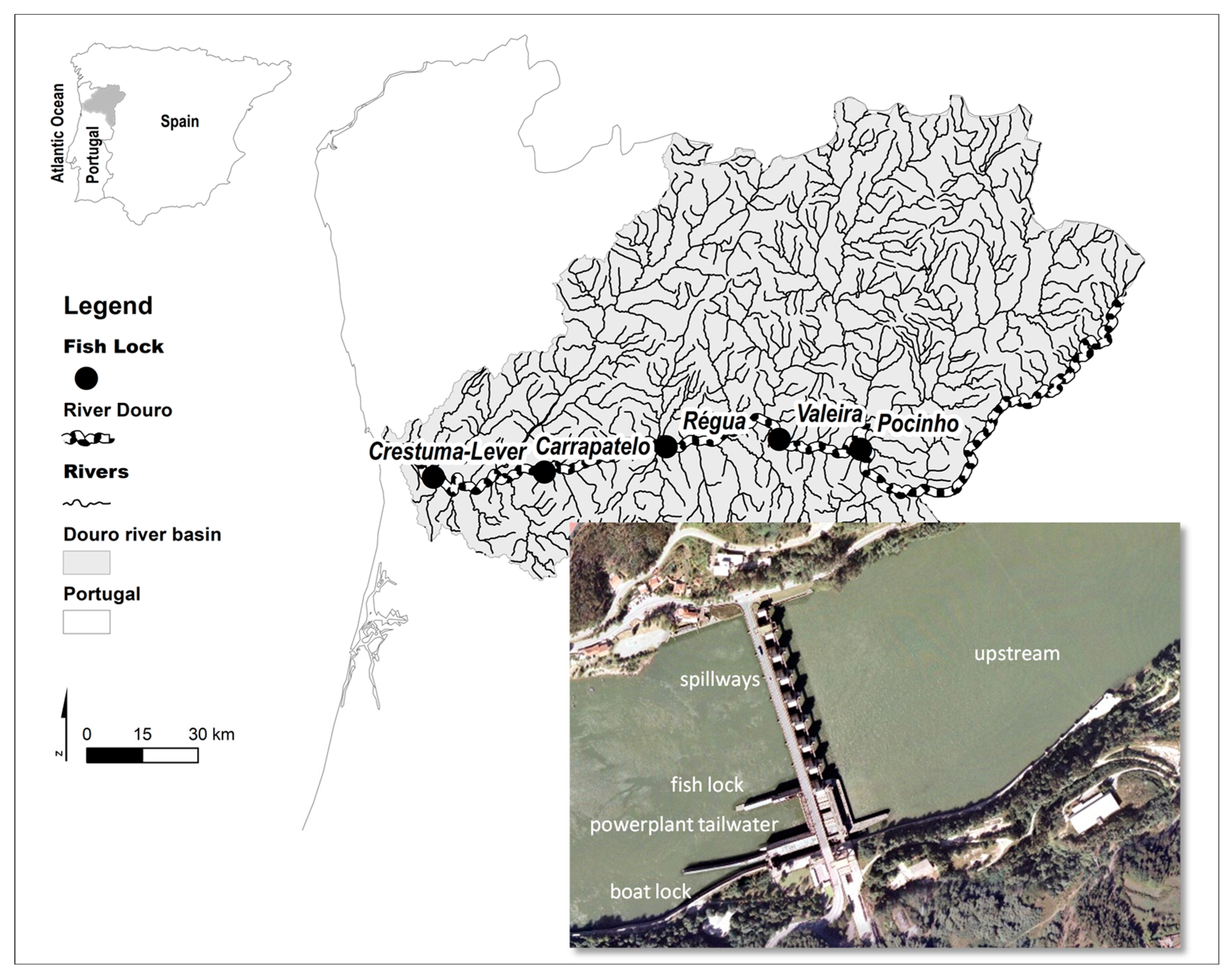
2.1. Study Area
2.2. Fish Lock Characteristics and Operation
2.3. Data Acquisition during Lock Cycles
2.4. Fish Captures by Professional Fishermen Downstream the Dam
2.5. Data Analysis
3. Results
3.1. Fish Lock
3.2. Creel Survey
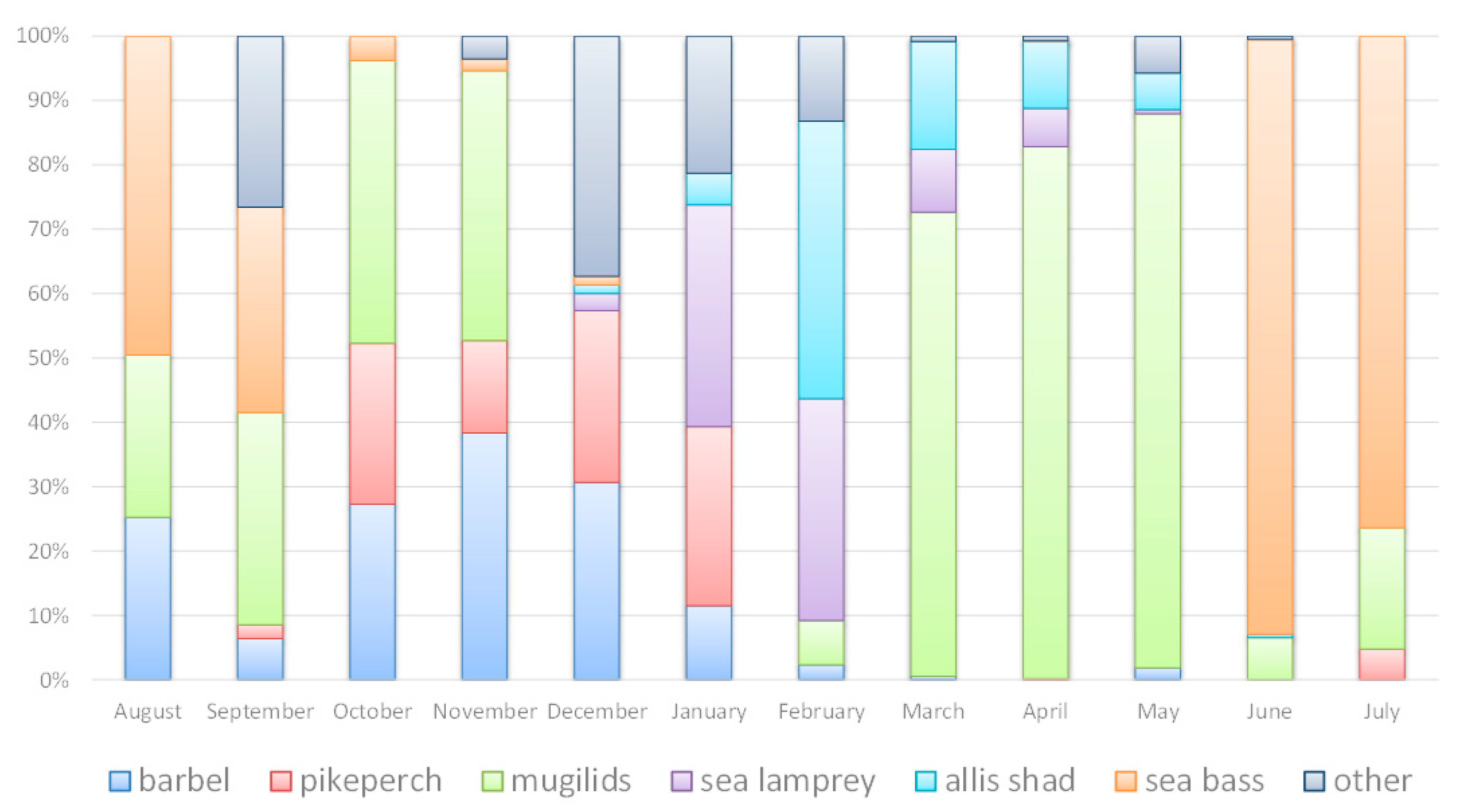
3.3. Selectivity in the Use of the Fish Lock
4. Discussion
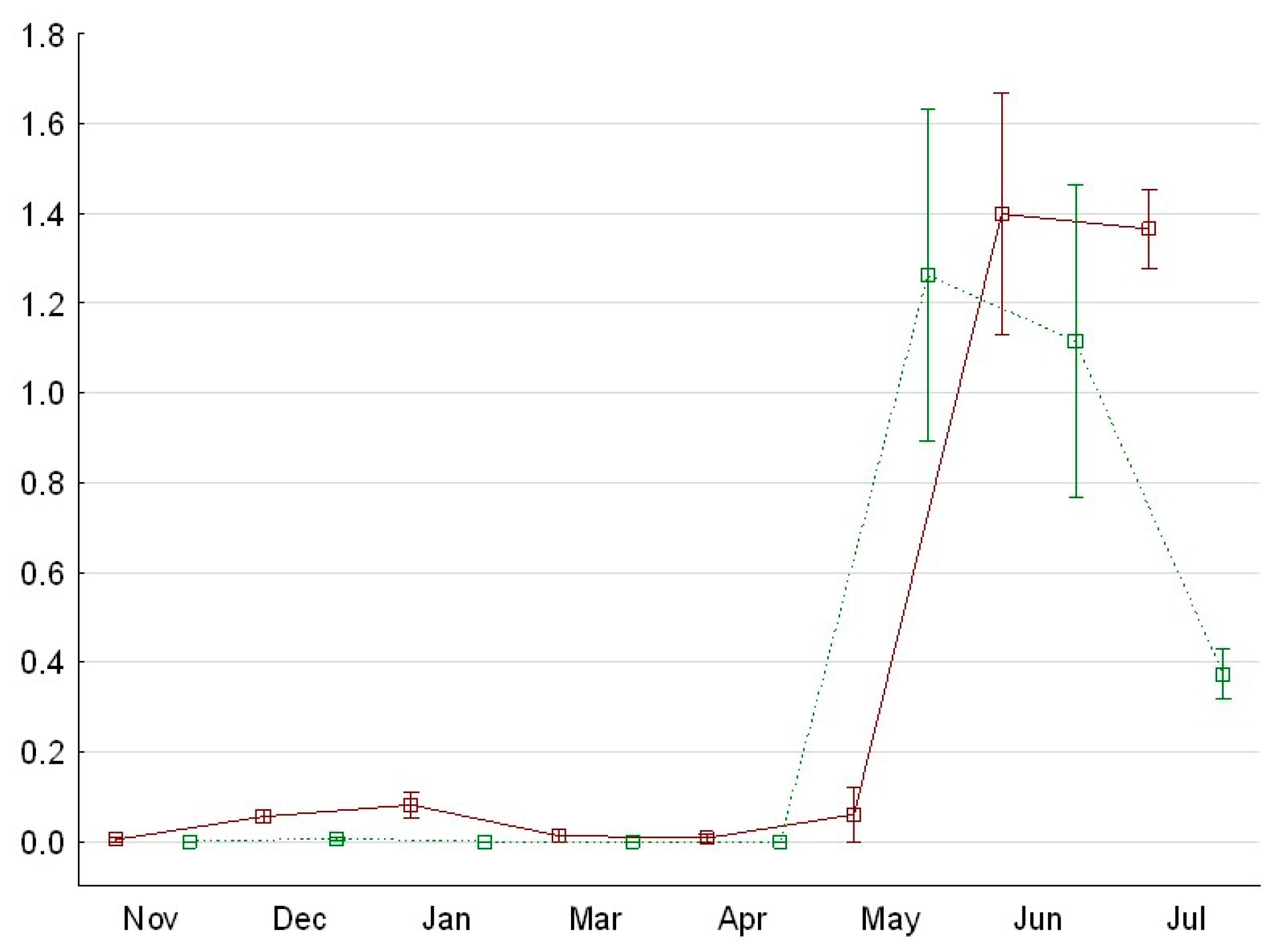
4.1. Temporal Use of the Fish Lock
4.2. Factors Influencing Lock Utilization by Fish
4.3. Selectivity of the Borland Fish Lock
4.4. Management Implications
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aitken, P.L.; Dickerson, L.H.; Menzies, W.J.M. Fish passes and screens at water power works. Proc. Inst. Civ. Eng. 1966, 32, 29–57. [Google Scholar] [CrossRef]
- Redeker, M. Retrofitting Fish Passes: International Best Practice, Current Research and Foreseeable Developments; U.S. Society on Dams: Denver, CO, USA, 2019; p. 15. [Google Scholar]
- O’Connor, W. Biology and Management of European Eel (Anguilla anguilla, L.) in the Shannon Estuary, Ireland. Ph.D. Thesis, National University of Ireland, Galway, Ireland, March 2003. [Google Scholar]
- Smith, D.L.; Nestler, J.M.; Maier, T.J. Planning Guide for Fish Passage at Pittsburgh District Dams; Technical Note ERDC WQTN-AM-16; Environmental Laboratory, Engineer Research and Development Center: Vicksburg, MS, USA, 2013. [Google Scholar]
- Travade, F.; Larinier, M. Fish Locks and Fish Lifts. Bull. Fr. Pêche Piscic. 2002, 364, 102–118. [Google Scholar] [CrossRef]
- Oldani, N.O.; Baigún, C.; Nestler, J.M.; Goodwin, R.A. Is Fish Passage Technology Saving Fish Resources in the Lower La Plata River Basin? Neotrop. Ichthyol. 2007, 5, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Stuart, I.G.; Berghuis, A.P.; Long, P.E.; Mallen-Cooper, M. Do fish locks have potential in tropical rivers? River Res. Appl. 2007, 23, 269–286. [Google Scholar] [CrossRef]
- Vieira, M.; Bordalo, A. The Douro estuary (Portugal): A mesotidal salt wedge. Oceanol. Acta 2000, 23, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, C.; Teixeira, C.; Teixeira, R.; Machado, A.; Azevedo, I.; Bordalo, A. Dissolved organic carbon and nitrogen dynamics in the Douro River estuary, Portugal. Cienc. Mar. 2008, 34, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Lobón-Cerviá, J.; Benigno, E.; Rincón, P.A. Historical Changes in the Fish Fauna of the River Duero basin. In Historical Change of Large Alluvial Rivers: Western Europe; Petts, G.E., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 1989; pp. 221–232. [Google Scholar]
- Ribeiro, F.; Collares-Pereira, M.; Moyle, P. Non-native fish in the fresh waters of Portugal, Azores and Madeira Islands: A growing threat to aquatic biodiversity. Fish. Manag. Ecol. 2009, 16, 255–264. [Google Scholar] [CrossRef]
- Martelo, J.; da Costa, L.; Ribeiro, D.; Gago, J.; Magalhães, M.; Gante, H.; Alves, M.; Cheoo, G.; Gkenas, C.; Banha, F.; et al. Evaluating the range expansion of recreational non-native fishes in Portuguese freshwaters using scientific and citizen science data. BioInvasions Rec. 2021, 10, 378–389. [Google Scholar] [CrossRef]
- Collares-Pereira, M.J.; Aves, M.J.; Ribeiro, F.; Domingos, I.; Almeida, P.R.; Costa, L.; Gante, H.; Filipe, A.F.; Aboim, M.A.; Rodrigues, P.M.; et al. Guia dos Peixes de Água Doce e Migradores de Portugal Continental; Edições Afrontamento: Lisboa, Portugal, 2021. [Google Scholar]
- Godinho, F.N.; Pinheiro, P.J. Occurrence of Atlantic salmon in the Douro River. Recent evidence. J. Appl. Ichthyol. 2019, 35, 1300–1302. [Google Scholar] [CrossRef]
- Rodrigues, S.M.; Silva, D.; Cunha, J.; Pereira, R.; Freitas, V.; Ramos, S. Environmental influences, particularly river flow alteration, on larval fish assemblages in the Douro Estuary, Portugal. Reg. Stud. Mar. Sci. 2022, 56, 102617. [Google Scholar] [CrossRef]
- Nicola, G.G.; Elvira, B.; Almodovar, A. Dams and fish passage facilities in the large rivers of Spain: Effects on migratory species. Large Rivers 1996, 10, 375–379. [Google Scholar] [CrossRef]
- StatSoft. STATISTICA for Windows (Computer Program Manual); StatSoft, Inc.: Tulsa, OK, USA, 2000. [Google Scholar]
- Bochechas, J. Condições de Funcionamento e de Eficácia de Eclusas Para Peixes: Casos das Barragens de Crestuma-Lever e de Belver. Master’s Thesis, Instituto Superior Técnico, Lisbon, Portugal, 1995. [Google Scholar]
- Santos, J.M.; Ferreira, M.T.; Godinho, F.N.; Bochechas, J. Performance of fish lift recently built at the Touvedo Dam on the Lima River, Portugal. J. Appl. Ichthyol. 2002, 18, 118–123. [Google Scholar] [CrossRef]
- Travade, F.; Larinier, M.; Boyer-Bernard, S.; Dartiguelongue, J. Performance of Four Fish Pass Installations Recently Built on Two Rivers in South-west France. In Proceedings of the Symposium on Fish Migration and Fish Bypass Channels, Vienna, Austria, 24–27 September 1996; Fishing News Books: Surrey, UK, 1998. [Google Scholar]
- Belo, A.F.; Cardoso, G.; Pereira, E.; Quintella, B.R.; Mateus, C.S.; Alexandre, C.M.; Batista, C.; Telhado, A.; Quadrado, M.F.; Almeida, P.R. Fish pass use by shads (Alosa alosa L. and Alosa fallax, Lacépède, 1803): Implications for monitoring and Management. Ecohydrology 2021, 14, e2292. [Google Scholar] [CrossRef]
- Pereira, E.; Quintella, B.R.; Lança, M.J.; Alexandre, C.M.; Mateus, C.S.; Pedro, S.; Belo, A.F.; Rato, A.S.; Quadrado, M.F.; Telhado, A.; et al. Temporal patterns of the catadromous thinlip grey mullet migration in freshwater. Ecohydrology 2021, 14, e2345. [Google Scholar] [CrossRef]
- McEvoy, B. Video Counting of Adult Salmon at Ardnacrusha Fish Pass; Fisheries Research Centre Internal Report: Dublin, Ireland, 1993. [Google Scholar]
- Lucas, M.C.; Baras, E. Methods for studying spatial behaviour of freshwater fishes in the natural environment. Fish Fish. 2000, 1, 283–316. [Google Scholar] [CrossRef]
- Feunteun, E.; Laffaille, P.; Robinet, T.; Briand, C.; Baisez, A.; Olivier, J.M.; Acou, A. A review of upstream migration and movements in inland waters by anguillid eels: Towards a general theory. In Eel Biology; Aida, K., Tsukamoto, K., Yanauchi, K., Eds.; Springer: Tokyo, Japan, 2003; pp. 191–213. [Google Scholar]
- Welsh, S.A.; Liller, H.L. Environmental correlates of upstream migration of yellow-phase American eels in the Potomac River drainage. Trans. Am. Fish. Soc. 2013, 142, 483–491. [Google Scholar] [CrossRef]
- Mameri, D.; Rivaes, R.; Oliveira, J.M.; Pádua, J.; Ferreira, M.T.; Santos, J.M. Passability of Potamodromous Species through a Fish Lift at a Large Hydropower Plant (Touvedo, Portugal). Sustainability 2020, 12, 172. [Google Scholar] [CrossRef] [Green Version]
- White, E.M.; Knights, B. Environmental factors affecting migration of the European eel in the Rivers Severn and Avon, England. J. Fish Biol. 1997, 50, 1104–1116. [Google Scholar] [CrossRef]
- Santos, J.M.; Rivaes, R.; Oliveira, J.; Ferreira, T. Improving yellow eel upstream movements with fish lifts. J. Ecohydraul. 2016, 1, 50–61. [Google Scholar] [CrossRef]
- Fernández-Delgado, C.; Drake, P.; Arias, A.M.; García-González, D. Peces de Doñana Y su Entorno; Ministerio de Medio Ambiente, Ed.; Colección OAPN, Naturaleza y Parques Nacionales: Madrid, Spain, 2000; p. 272. [Google Scholar]
- Santos, J.M.; Ferreira, M.T.; Godinho, F.N.; Bochechas, J. Efficacy of a nature-like bypass channel in a Portuguese lowland river. J. Appl. Ichthyol. 2005, 21, 381–388. [Google Scholar] [CrossRef]
- Ordeix, M.; Pou-Rovira, Q.; Sellarès, N.; Bardina, M.; Casamitjana, A.; Solà, C.; Munné, A. Fish pass assessment in the rivers of Catalonia (NE Iberian Peninsula). A case study of weirs associated with hydropower plants and gauging stations. Limnetica 2011, 30, 405–426. [Google Scholar] [CrossRef]
- Aparicio, E.; Pintor, C.; Durán, C.; Carmona-Catot, G. Fish passage assessment at the most downstream barrier of the Ebro River (NE Iberian Peninsula). Limnetica 2012, 31, 37–46. [Google Scholar] [CrossRef]
- Lopez, V.; Franch, N.; Pou, Q.; Clavero, M.; Gaya, N.; Queral, J.M. Atles dels Peixos del Delta de l’Ebre. Collecci·ó Tècnica, 3. Generalitat de Catalunya, 1st ed.; Departament d’Agricultura, Ramaderia, Pesca I Medi Natural: Catalonia, Spain, 2015; pp. 1–224. [Google Scholar]
- Rodriguez-Ruiz, A.; Granado-Lorencio, C. Spawning period and migration of three species of cyprinids in a stream with Mediterranean regimen (SW Spain). J. Fish Biol. 1992, 41, 545–556. [Google Scholar] [CrossRef]
- Sato, Y.; Bazzoli, N.; Rizzo, E.; Boschi, M.B.; Miranda, M.O.T. Influence of the Abaeté River on the reproductive success of the neotropical migratory teleost Prochilodus argenteus in the São Francisco River, downstream from the Três Marias Dam, southeastern Brazil. River Res. Appl. 2005, 21, 939–950. [Google Scholar] [CrossRef]
- Jonsson, B.; Jonsson, N.; Hansen, L.P. Factors affecting river entry of adult Atlantic salmon in a small river. J. Fish Biol. 2007, 71, 943–956. [Google Scholar] [CrossRef]
- Boavida, I.; Jesus, J.B.; Pereira, V.; Santos, C.; Lopes, M.; Cortes, R.M.V. Fulfilling spawning flow requirements for potamodromous cyprinids in a restored river segment. Sci. Total Environ. 2018, 635, 567–575. [Google Scholar] [CrossRef]
- Wright, G.V.; Wright, R.M.; Kemp, P.S. Impact of tide gates on the migration of adult European eels, Anguilla anguilla. Estuaries Coasts 2015, 38, 2031–2043. [Google Scholar] [CrossRef] [Green Version]
- Kroes, R.; Van Loon, E.E.; Goverse, E.; Schiphouwer, M.E.; van der Geest, H.G. Attraction of migrating glass eel (Anguilla anguilla) by freshwater flows from water pumping stations in an urbanized delta system. Sci. Total Environ. 2020, 714, 136818. [Google Scholar] [CrossRef]
- Sauriau, P.G.; Robin, J.P.; Marchand, J. Effects of the Excessive Organic Enrichment of the Loire Estuary on the Downstream Migratory Patterns of the Amphihaline Grey Mullet Liza Ramada (Pisces, Mugilidae). In Changes in Fluxes in Estuaries (ECSA22/ERF Symposium, Plymouth, September 1992); Dyer, K.R., Orth, R.J., Eds.; Olsen & Olsen: Fredensborg, Denmark, 1994; pp. 349–356. [Google Scholar]
- Jellyman, D.J.; Ryan, C.M. Seasonal migration of elvers (Anguilla spp.) into Lake Pounui, New Zealand, 1974–1978. N. Z. J. Mar. Freshw. Res. 1983, 17, 1–15. [Google Scholar] [CrossRef]
- Jonsson, N. Influence of water flow, water temperature and light on fish migration in rivers. Nord. J. Freshw. Res. 1991, 66, 20–35. [Google Scholar]
- Pereira, E.; Cardoso, G.R.; Quintella, B.R.; Mateus, C.S.; Alexandre, C.M.; Oliveira, R.L.; Belo, A.F.; Telhado, A.; Quadrado, M.F.; Batista, C.M.; et al. Proposals for optimizing sea lamprey passage through a vertical-slot fishway. Ecohydrology 2019, 12, e2087. [Google Scholar] [CrossRef]
- Griffioen, A.B.; vanKeeken, O.A.; Hamer, A.L.; Winter, H.V. Passage efficiency and behaviour of sea lampreys (Petromyzon marinus, Linnaeus 1758) at a large marine–freshwater barrier. River Res. Appl. 2022, 38, 906–916. [Google Scholar] [CrossRef]
- Almeida, P.R. Estuarine movement patterns of adult thin-lipped grey mullet, Liza ramada (Risso) (Pisces, Mugilidae), observed by ultrasonic tracking. J. Exp. Mar. Biol. Ecol. 1996, 202, 137–150. [Google Scholar] [CrossRef]
- Wirjoatmodjo, S.; Pitcher, T.J. Flounders follow the tides to feed: Evidence from ultrasonic tracking in an estuary. Estuar. Coast. Shelf Sci. 1984, 19, 231–241. [Google Scholar] [CrossRef]
- Trancart, T.; Lambert, P.; Rochard, E.; Daverat, F.; Coustillas, J.; Roqueplo, C. Alternative flood tide transport tactics in catadromous species: Anguilla anguilla, Liza ramada and Platichthys flesus. Estuar. Coast. Shelf Sci. 2012, 99, 191–198. [Google Scholar] [CrossRef]
- Sugeha, H.Y.; Arai, T.; Miller, M.J.; Limbong, D.; Tsukamoto, K. Inshore migration of the tropical eels Anguilla spp. recruiting to the Poigar River estuary on north Sulawesi Island. Mar. Ecol. Prog. Ser. 2001, 221, 233–243. [Google Scholar] [CrossRef]
- Hammond, S.D.; Welsh, S.A. Seasonal Movements of Large Yellow American Eels Downstream of a Hydroelectric Dam, Shenandoah River, West Virginia. In Eels at the Edge: Science, Status, and Conservation Concerns; Casselman, J.M., Cairns, D.K., Eds.; American Fisheries Society: Bethesda, MD, USA, 2009; Volume 58, pp. 309–323. [Google Scholar]
- Torricelli, P.; Tongiorgi, P.; Almansi, P. Migration of grey mullet fry into the Arno River: Seasonal appearance, daily activity, and feeding rhythms. Fish. Res. 1982, 1, 219–234. [Google Scholar] [CrossRef]
- Albertini-Berhaut, J. Rythme alimentaire chez les jeunes Mugil capito (Téléostééns Mugilidae) dans le Golfe de Marseille. Tehys 1979, 9, 79–82. [Google Scholar]
- Almeida, P.R. Feeding ecology of Liza ramada (Risso 1810) (Pisces: Mugilidae) in a south-western estuary of Portugal. Estuar. Coast. Shelf Sci. 2003, 57, 313–323. [Google Scholar] [CrossRef]
- Almeida, P.R.; Quintella, B.R.; Mateus, C.S.; Alexandre, C.M.; Pedro, S. Diadromous Fish in Portugal: Status, Threats and Management Guidelines. In Desenvolvimento Sustentável do Oceano: Uma Utopia útil; Bebiano, T., Carvalho, I., Gameiro, J., Guerreiro, J., Eds.; Universidade do Algarve: Faro, Portugal, 2018; pp. 189–213. [Google Scholar]
- Cabral, M.J.; Almeida, J.; Almeida, P.R.; Dellinger, T.; Ferrand de Almeida, N.; Oliveira, M.; Palmeirim, J.M.; Queiroz, A.I.; Rogado l Santos Reis, M. (Eds.) Livro Vermelho dos Vertebrados de Portugal; Instituto de Conservação da Natureza: Lisboa, Portugal, 2005; pp. 1–660. [Google Scholar]
- Larinier, M.; Travade, F. The design of fishways for shad. Bull. Fr. Pêche Piscic 2002, 364, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dam and Reservoir | |
| Installed capacity | 117 MW (three Kaplan turbines) |
| Maximum discharged volume at each turbine | 440 m3/s |
| Height of the crest | 25.50 m |
| Average flow at the dam | 450 m3/s |
| Inundated area at pool level | 12.98 km2 |
| Total volume | 22.5 hm3 |
| Douro river | |
| Total length | 897 km |
| Catchment area | 97,600 km2 |
| Average total discharge | 11,349 hm3 |
| Phase Duration | |
| Attraction/fishing phase | 60 min |
| Filling stage | 20–25 min |
| Passing stage | 45 min |
| Emptying phase | 25–30 min |
| Other characteristics | |
| Flow during the attraction phase | 0.5–1.0 m3/s |
| Velocity at the entrance during the attraction phase (variable according to the downstream water level and the position of the upstream sluice gate) | 1.2–2.6 m/s−1 |
| Velocity inside the lock during fish passing | 0.3 m/s |
| Month | Total Number of Cycles | Number of Cycles with Fish | Mugilidae | European Eel | Iberian Barbel | Pumpkin Seed Sunfish | Largemouth Bass | Bleak | European Flounder | Unidentified Cypriniformes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U | D | U | D | U | D | U | D | U | D | U | D | U | D | U | D | |||
| November 2016 | 72 | 12 | 0 | 0 | 1 | 1 | 0 | 0 | 11 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| December 2016 | 240 | 44 | 6 | 1 | 160 | 3 | 0 | 0 | 0 | 3 | 0 | 0 | 17 | 0 | 0 | 0 | 6 | 5 |
| January 2017 | 116 | 10 | 0 | 0 | 158 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| March 2017 | 85 | 1 | 79 | 0 | 11 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| April 2017 | 82 | 1 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| May 2017 | 16 | 11 | 3264 | 0 | 8 | 0 | 1263 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| June 2017 | 13 | 13 | 6491 | 0 | 177 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 |
| July 2017 | 146 | 142 | 2719 | 2 | 44,104 | 1 | 9 | 0 | 159 | 0 | 9 | 0 | 221 | 7 | 3 | 0 | 55 | 6 |
| TOTAL | 770 | 234 | 12,559 | 3 | 44,622 | 6 | 1272 | 0 | 170 | 4 | 9 | 0 | 255 | 7 | 3 | 0 | 61 | 11 |
| European Eel | |||
| F | p | Adjusted R2 | |
| Day/night | 4.84 | <0.05 | 0.01 |
| Moon phase | 40.41 | <0.01 | 0.1 |
| Cycle type | 0.06 | NS | - |
| Month | 125.14 | <0.01 | 0.53 |
| Tide height | 8.66 | <0.01 | 0.02 |
| Discharged flow | 295.9 | <0.01 | 0.27 |
| Mugilids | |||
| F | p | Adjusted R2 | |
| Day/night | 15.74 | <0.01 | 0.02 |
| Moon phase | 6.74 | <0.01 | 0.01 |
| Cycle type | 0.21 | NS | - |
| Month | 49.2 | <0.01 | 0.31 |
| Tide height | 8.07 | <0.01 | 0.02 |
| Discharged flow | 68.85 | <0.01 | 0.09 |
| European Eel | Month | Discharged Flow | Moon Phase |
| F | 57.4 | 0.54 | 15.75 |
| Partial η2 | 0.35 | <0.01 | 0.06 |
| Mugilids | Month | Discharged flow | Moon phase |
| F | 36.2 | 0.26 | |
| Partial η2 | 0.25 | <0.01 |
| Iberian Barbel | Pikeperch | Mugilids | Sea Lamprey | Allis Shad | Twaite Shad | European Sea Bass | European Flounder | Others * | |
|---|---|---|---|---|---|---|---|---|---|
| Aug | 7.3 | 0.0 | 7.3 | 0.0 | 0.0 | 0.0 | 14.3 | 0.0 | 0.0 |
| Sep | 3.0 | 1.0 | 15.5 | 0.0 | 0.0 | 0.0 | 15.0 | 1.5 | 11.0 |
| Oct | 9.0 | 8.3 | 14.5 | 0.0 | 0.0 | 0.0 | 1.3 | 0.0 | 0.0 |
| Nov | 8.0 | 3.0 | 8.8 | 0.0 | 0.0 | 0.0 | 0.4 | 0.0 | 0.8 |
| Dec | 2.6 | 2.2 | 0.0 | 0.2 | 0.1 | 0.1 | 0.1 | 1.4 | 1.7 |
| Jan | 0.3 | 0.7 | 0.0 | 0.9 | 0.1 | 0.0 | 0.0 | 0.3 | 0.2 |
| Feb | 0.1 | 0.0 | 0.2 | 1.0 | 1.2 | 0.0 | 0.0 | 0.4 | 0.0 |
| Mar | 0.1 | 0.0 | 12.5 | 1.7 | 2.9 | 0.0 | 0.0 | 0.2 | 0.0 |
| Apr | 0.0 | 0.0 | 12.0 | 0.9 | 1.5 | 0.1 | 0.0 | 0.0 | 0.0 |
| May | 0.4 | 0.0 | 20.4 | 0.1 | 1.4 | 1.4 | 0.0 | 0.0 | 0.0 |
| Jun | 0.0 | 0.0 | 2.4 | 0.0 | 0.2 | 0.0 | 34.0 | 0.0 | 0.4 |
| Jul | 0.0 | 2.4 | 9.4 | 0.0 | 0.0 | 0.0 | 37.9 | 0.0 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godinho, F.N.; Pinheiro, P.J.; Benites, L. Fish Use of a Borland-Type Fish Lock in an Iberian River. Diversity 2023, 15, 178. https://doi.org/10.3390/d15020178
Godinho FN, Pinheiro PJ, Benites L. Fish Use of a Borland-Type Fish Lock in an Iberian River. Diversity. 2023; 15(2):178. https://doi.org/10.3390/d15020178
Chicago/Turabian StyleGodinho, Francisco N., Paulo J. Pinheiro, and Liliana Benites. 2023. "Fish Use of a Borland-Type Fish Lock in an Iberian River" Diversity 15, no. 2: 178. https://doi.org/10.3390/d15020178