
Phytoplankton Diversity and Blooms in Ephemeral Saline Lakes of Cyprus

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
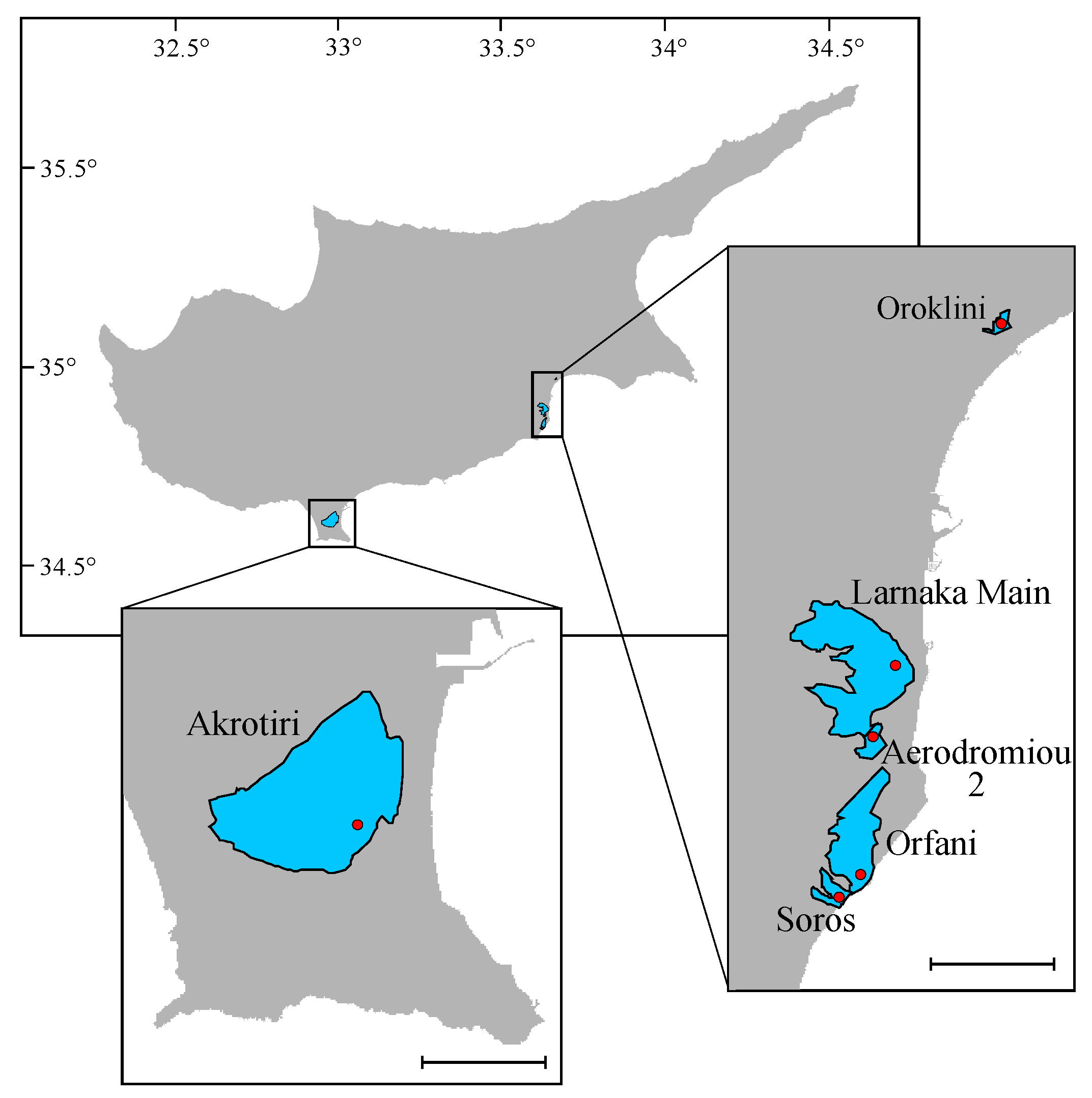
2.1. Study Area and Field Work
2.2. Microscopic Analysis
2.3. Land Use Coverage
2.4. Data Analysis
3. Results
3.1. Environmental Factors and Catchment Land Use Coverage
3.2. Phytoplankton Community Dynamics
3.2.1. Composition, Abundance and Biomass
3.2.2. Composition, Abundance and Biomass
Alpha Diversity
Beta Diversity
3.3. Environmental Effects on Phytoplankton Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, W.D. Conservation of salt lakes. Hydrobiologia 1993, 267, 291–306. [Google Scholar] [CrossRef]
- Saccò, M.; White, N.E.; Harrod, C.; Salazar, G.; Aguilar, P.; Cubillos, C.F.; Meredith, K.; Baxter, B.K.; Oren, A.; Anufriieva, E.; et al. Salt to conserve: A review on the ecology and preservation of hypersaline ecosystems. Biol. Rev. 2021, 96, 2828–2850. [Google Scholar] [CrossRef] [PubMed]
- Zadereev, E.; Lipka, O.; Karimov, B.; Krylenko, M.; Elias, V.; Pinto, I.S.; Alizade, V.; Anker, Y.; Feest, A.; Kuznetsova, D.; et al. Overview of past, current, and future ecosystem and biodiversity trends of inland saline lakes of Europe and Central Asia. Inland Waters 2020, 10, 438–452. [Google Scholar] [CrossRef]
- García, C.M.; García-Ruiz, R.; Rendón, M.; Niell, F.X.; Lucena, J. Hydrological cycle and interannual variability of the aquatic community in a temporary saline lake (Fuente de Piedra, Southern Spain). Hydrobiologia 1997, 345, 131–141. [Google Scholar] [CrossRef]
- Williams, W.D. Environmental threats to salt lakes and the likely status of inland saline ecosystems in 2025. Environ. Conserv. 2002, 29, 154–167. [Google Scholar] [CrossRef]
- Hammer, U.T. Saline Lake Ecosystems of the World; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1986; Volume 59. [Google Scholar]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.D.; Muyzer, G. Microbial diversity and biogeochemical cycling in soda lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef] [PubMed]
- Larson, C.A.; Belovsky, G.E. Salinity and nutrients influence species richness and evenness of phytoplankton communities in microcosm experiments from Great Salt Lake, Utah, USA. J. Plankton Res. 2013, 35, 1154–1166. [Google Scholar] [CrossRef]
- Williams, W.D. Salinity as a determinant of the structure of biological communities in salt lakes. Hydrobiologia 1998, 381, 191–201. [Google Scholar] [CrossRef]
- Padisak, J.; Naselli-Flores, L. Phytoplankton in extreme environments: Importance and consequences of habitat permanency. Hydrobiologia 2021, 848, 157–176. [Google Scholar] [CrossRef]
- Alexe, M.; Șerban, G.; Baricz, A.; Andrei, A.; Cristea, A.; Battes, K.P.; Cîmpean, M.; Momeu, L.; Muntean, V.; Porav, S.; et al. Limnology and plankton diversity of salt lakes from Transylvanian Basin (Romania): A review. J. Limnol. 2017, 77, 17–34. [Google Scholar] [CrossRef]
- Ballot, A.; Krienitz, L.; Kotut, K.; Wiegand, C.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Cyanobacteria and cyanobacterial toxins in three alkaline Rift Valley lakes of Kenya—Lakes Bogoria, Nakuru and Elmenteita. J. Plankton Res. 2004, 26, 925–935. [Google Scholar] [CrossRef]
- Costa, N.B.; Kolman, M.A.; Giani, A. Cyanobacteria diversity in alkaline saline lakes in the Brazilian Pantanal wetland: A polyphasic approach. J. Plankton Res. 2016, 38, 1389–1403. [Google Scholar] [CrossRef]
- Stefanidou, N.; Katsiapi, M.; Tsianis, D.; Demertzioglou, M.; Michaloudi, E.; Moustaka-Gouni, M. Patterns in alpha and beta phytoplankton diversity along a conductivity gradient in coastal mediterranean lagoons. Diversity 2020, 12, 38. [Google Scholar] [CrossRef]
- Schagerl, M.; Oduor, S.O. Phytoplankton community relationship to environmental variables in three Kenyan Rift Valley saline-alkaline lakes. Mar. Freshw. Res. 2008, 59, 125–136. [Google Scholar] [CrossRef]
- Rad, F.A.; Aksoz, N.; Hejazi, M.A. Effect of salinity on cell growth and β-carotene production in Dunaliella sp. isolates from Urmia Lake in northwest of Iran. Afr. J. Biotechnol. 2011, 10, 2282–2289. [Google Scholar]
- Oren, A. The ecology of Dunaliella in high-salt environments. J. Biol. Res. Thessalon. 2014, 21, 23. [Google Scholar] [CrossRef]
- Williams, W.D. Comments on the so-called salt lakes of Greenland. Hydrobiologia 1991, 210, 67–74. [Google Scholar] [CrossRef]
- Salm, C.R.; Saros, J.E.; Martin, C.S.; Erickson, J.M. Patterns of seasonal phytoplankton distribution in prairie saline lakes of the northern Great Plains (USA). Saline Syst. 2009, 5, 1–13. [Google Scholar] [CrossRef]
- Lind, L.; Schuler, M.S.; Hintz, W.D.; Stoler, A.B.; Jones, D.K.; Mattes, B.M.; Relyea, R.A. Salty fertile lakes: How salinization and eutrophication alter the structure of freshwater communities. Ecosphere 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Afonina, E.Y.; Tashlykova, N.A. Plankton community and the relationship with the environment in saline lakes of On-Torey plain, Northeastern Mongolia. Saudi J. Biol. Sci. 2018, 25, 399–408. [Google Scholar] [CrossRef]
- Afonina, E.Y.; Tashlykova, N.A. Fluctuations in plankton community structure of endorheic soda lakes of southeastern Transbaikalia (Russia). Hydrobiologia 2020, 847, 1383–1398. [Google Scholar] [CrossRef]
- Afonina, E.Y.; Tashlykova, N.A. Plankton of Saline Lakes in Southeastern Transbaikalia: Transformation and Environmental Factors. Contemp. Probl. Ecol. 2019, 12, 155–170. [Google Scholar] [CrossRef]
- Moore, J.N. Recent desiccation of western Great Basin saline lakes: Lessons from Lake Abert, Oregon, USA. Sci. Total Environ. 2016, 554, 142–154. [Google Scholar] [CrossRef]
- Shadrin, N.; Zheng, M.; Oren, A. Past, present and future of saline lakes: Research for global sustainable development. Chin. J. Oceanol. Limnol. 2015, 33, 1349–1353. [Google Scholar] [CrossRef]
- Erol, A.; Randhir, T.O. Climatic change impacts on the ecohydrology of Mediterranean watersheds. Clim. Chang. 2012, 114, 319–341. [Google Scholar] [CrossRef]
- Zacharias, I.; Dimitriou, E.; Dekker, A.; Dorsman, E. Overview of temporary ponds in the Mediterranean region: Threats, management and conservation issues. J. Environ. Biol. 2007, 28, 1–9. [Google Scholar]
- Council of the European Commission. Council directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Communities Bruss. 1992, 206, 7–49. [Google Scholar]
- Ramsar List. The List of Wetlands of International Importance. 2023. Available online: https://www.ramsar.org/sites/default/files/2023-08/sitelist.pdf (accessed on 30 August 2023).
- Katsiapi, M.; Mazaris, A.D.; Charalampous, E.; Moustaka-Gouni, M. Watershed land use types as drivers of freshwater phytoplankton structure. In Phytoplankton Responses to Human Impacts at Different Scales, Hydrobiologia; Springer: Berlin/Heidelberg, Germany, 2012; pp. 121–131. [Google Scholar]
- Christia, C.; Tziortzis, I.; Fyttis, G.; Kashta, L.; Papastergiadou, E. A survey of the benthic aquatic flora in transitional water systems of Greece and Cyprus (Mediterranean Sea). Bot. Mar. 2011, 54, 169–178. [Google Scholar] [CrossRef]
- Karagianni, A.; Stamou, G.; Katsiapi, M.; Polykarpou, P.; Dörflinger, G.; Michaloudi, E. Zooplankton communities in Mediterranean temporary lakes: The case of saline lakes in Cyprus. Ann. Limnol. Int. J. Limnol. 2018, 54, 14. [Google Scholar] [CrossRef]
- European Commission. Directive 2000/60/EC of the European Parliament and of the Council Establishing a Framework for Community Action in the Field of Water Policy; European Commission: Brussels, Belgium, 2000.
- Hadjisterkotis, E.; Charalambides, M. The first evidence for the breeding of the Greater Flamingo Phoenicopterus ruber on Cyprus. Zeitschrift für Jagdwissenschaft. 2002, 48, 72–76. [Google Scholar]
- Ramsar Sites Information Service. Annotated List of Wetlands of International Importance: United Kingdom of Great Britain and Northern Ireland. 1999. Available online: https://rsis.ramsar.org/sites/default/files/rsiswp_search/exports/Ramsar-Sites-annotated-summary-United-Kingdom-of-Great-Britain-and-Northern-Ireland.pdf?1686509325 (accessed on 30 August 2023).
- Ramsar Sites Information Service. Annotated List of Wetlands of International Importance: Cyprus. 2008. Available online: https://rsis.ramsar.org/ris/1081 (accessed on 30 August 2023).
- Giosa, E.; Mammides, C.; Zotos, S. The importance of artificial wetlands for birds: A case study from Cyprus. PLoS ONE 2018, 13, e0197286. [Google Scholar] [CrossRef] [PubMed]
- Gucel, S.; Kadis, C.; Ozden, O.; Charalambidou, I.; Linstead, C.; Fuller, W.; Kounnamas, C.; Ozturk, M. Assessment of biodiversity differences between natural and artificial wetlands in Cyprus. Pak. J. Bot. 2012, 44, 213–224. [Google Scholar]
- Bold, H.C.; Wynne, M.J. Introduction to the Algae, 2nd ed.; Benjamin Cummings: San Francisco, CA, USA, 1984; 720p. [Google Scholar]
- Canter-Lund, H.; Lund, J.W. Freshwater Algae: Their Microscopic World Explored; Biopress Limited: Bristol, UK, 1994; 360p. [Google Scholar]
- Hallegraeff, G.M.; Bolch, C.J.S.; Hill, D.R.A.; Jameson, I.; Leroi, J.M.; McMinn, A.; Murray, S.; de Salas, M.F.; Saunders, K. Algae of Australia: Phytoplankton of Temperate Coastal Waters (Algae of Australia Series); Canberra & Melbourne: ABRS; CSIRO Publishing. 2010. Available online: https://hdl.handle.net/102.100.100/538278 (accessed on 30 August 2023).
- Hoppenrath, M.; Elbrächter, M.; Drebes, G. Marine Phytoplankton—Selected Microphytoplankton Species from the North Sea around Helgoland and Sylt; Kleine Senckenberg-Reihr, Bd 49; Schweizerbart Sche Vlgsb.: Stuttgart, Germany, 2010. [Google Scholar]
- Huber-Pestalozzi, G. Das Phytoplankton des Süsswassers. In August Thienemann, Die Binnengewässer; E. Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 1942; Volume 16/2, pp. 367–549. [Google Scholar]
- Kraberg, A.; Baumann, M.; Dürselen, C.-D. Coastal Phytoplankton: Photo Guide for Northern European Seas; VerlagDr. Friedrich Pfeil.: Munchen, Germany, 2010. [Google Scholar]
- Tikkanen, T. Kasviplanktonopas. Suomen Luonnonsuojelun Tuki Oy; Forssan kirjapaino Oy, Forssa: Helsinki, Finland, 1986. [Google Scholar]
- CEN 15204; Water Quality—Guidance Standard on the Enumeration of Phytoplankton Using Inverted Microscopy (Utermöhl Technique). European Committee for Standardization, Management Centre: Brussels, Belgium, 2006.
- CEN/EN 16695; Water Quality—Guidance on the Estimation of Microalgal Biovolume. European Committee for Standardization (CEN): Brussels, Belgium, 2015.
- Utermöhl, H. Zur Vervollkommnung der quantitative Phytoplanktonmethodik. Mitteilungen Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- European Environmental Agency. Corine Land Cover Technical Quide; Addendum; European Environmental Agency: Copenhagen, Denmark, 2000.
- Hammer, Ø.; Harper, D.A. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Baselga, A. Multiplicative partition of true diversity yields independent alpha and beta components; additive partition does not. Ecology 2010, 91, 1974–1981. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer. PRIMER-e, Plymouth; Plymouth Marine Laboratory: Plymouth, UK, 2006; 866p. [Google Scholar]
- Dowle, M.; Srinivasan, A.; Gorecki, J.; Chirico, M.; Stetsenko, P.; Short, T.; Lianoglou, S.; Antonyan, E.; Bonsch, M.; Parsonage, H.; et al. Data. Table R Package, Version 1.14.0. 2021. Available online: https://cran.r-project.org/web/packages/data.table/index.html (accessed on 30 August 2023).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.5-7. 2020. Available online: https://cran.r-project.org/ (accessed on 30 August 2023).
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; pp. 1–269. [Google Scholar]
- Diego, F.; Yamila, B.; Gisela, M.; Patricia, M. Controlling factors in planktonic communities over a salinity gradient in high-altitude lakes. Ann. Limnol. Int. J. Limnol. 2015, 51, 261–272. [Google Scholar] [CrossRef]
- Li, Z.; Gao, Y.; Wang, S.; Lu, Y.; Sun, K.; Jia, J.; Wang, Y. Phytoplankton community response to nutrients along lake salinity and altitude gradients on the Qinghai-Tibet Plateau. Ecol. Indic. 2021, 128, 107848. [Google Scholar] [CrossRef]
- Ramdani, M.; Elkhiati, N.; Flower, R.J.; Thompson, J.R.; Chouba, L.; Kraiem, M.M.; Ayache, F.; Ahmed, M.H. Environmental influences on the qualitative and quantitative composition of phytoplankton and zooplankton in North African coastal lagoons. Hydrobiologia 2009, 622, 113–131. [Google Scholar] [CrossRef]
- LAWA. German Guidance Document for the Implementation of the EC Water Framework Directive. 2003. Available online: http://www.lawa.de/Publikationen.html (accessed on 30 April 2003).
- Jellison, R.; Zadereev, Y.S.; DasSarma, P.A.; Melack, J.M.; Rosen, M.R.; Degermendzhy, A.G.; DasSarma, S.; Zambrana, G. Conservation and Management Challenges of Saline Lakes: A Review of Five Experience Briefs; Lake Basin Management Initiative: Thematic Paper. 2004. Available online: http://www.worldlakes.org/uploads/ISSLR_Review_of_Salt_Lakes.pdf (accessed on 30 August 2023).
- Alvarez, S.; Diaz, P.; Lopez-Archilla, A.I.; Guerrero, M.C. Phytoplankton composition and dynamics in three shallow temporary salt lakes (Monegros, Spain). J. Arid Environ. 2006, 65, 553–571. [Google Scholar] [CrossRef]
- Temponeras, M.; Kristiansen, J.; Moustaka-Gouni, M. Seasonal variation in phytoplankton composition and physical-chemical features of the shallow Lake Doïrani, Macedonia, Greece. In The Trophic Spectrum Revisited: The Influence of Trophic State on the Assembly of Phytoplankton Communities, Proceedings of the 11th Workshop of the International Association of Phytoplankton Taxonomy and Ecology (IAP), Shrewsbury, UK, 15–23 August 1998; Springer: Dordrecht, The Netherlands, 2000; pp. 109–122. [Google Scholar]
- Katsiapi, M.; Moustaka-Gouni, M.; Michaloudi, E.; Kormas, K.A. Phytoplankton and water quality in a Mediterranean drinking-water reservoir (Marathonas Reservoir, Greece). Environ. Monit. Assess. 2011, 181, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Belovsky, G.E.; Stephens, D.; Perschon, C.; Birdsey, P.; Paul, D.; Naftz, D.; Baskin, R.; Larson, C.; Mellison, C.; Luft, J.; et al. The Great Salt Lake Ecosystem (Utah, USA): Long term data and a structural equation approach. Ecosphere 2011, 2, 1–40. [Google Scholar] [CrossRef]
- Hodgson, D.A.; Vyverman, W.; Sabbe, K. Limnology and biology of saline lakes in the Rauer Islands, eastern Antarctica. Antarct. Sci. 2001, 13, 255–270. [Google Scholar] [CrossRef]
- Kebede, E.; Willén, E. Phytoplankton in a Salinity-Alkalinity Series of Lakes in the Ethiopian Rift Valley; Acta Universitatis Upsaliensis: Uppsala, Sweden, 1998; Volume 89, pp. 63–96. [Google Scholar]
- Moustaka-Gouni, M. Phytoplankton succession and diversity in a warm monomictic, relatively shallow lake: Lake Volvi, Macedonia, Greece. Hydrobiologia 1993, 249, 33–42. [Google Scholar] [CrossRef]
- Katsiapi, M.; Moustaka-Gouni, M.; Vardaka, E.; Kormas, K.A. Different phytoplankton descriptors show asynchronous changes in a shallow urban lake (L. Kastoria, Greece) after sewage diversion. Fundam. Appl. Limnol. 2013, 182, 219–230. [Google Scholar] [CrossRef]
- Chrisostomou, A.; Moustaka-Gouni, M.; Sgardelis, S.; Lanaras, T. Air-dispersed phytoplankton in a Mediterranean river-reservoir system (Aliakmon-Polyphytos, Greece). J. Plankton Res. 2009, 31, 877–884. [Google Scholar] [CrossRef]
- Genitsaris, S.; Moustaka-Gouni, M.; Kormas, K.A. Airborne microeukaryote colonists in experimental water containers: Diversity, succession, life histories and established food webs. Aquat. Microb. Ecol. 2011, 62, 139–152. [Google Scholar] [CrossRef]
- Genitsaris, S.; Stefanidou, N.; Beeri-Shlevin, Y.; Viner-Mozzini, Y.; Moustaka-Gouni, M.; Ninio, S.; Sukenik, A. Air-dispersed aquatic microorganisms show establishment and growth preferences in different freshwater colonisation habitats. FEMS Microbiol. Ecol. 2021, 97, fiab122. [Google Scholar] [CrossRef]
- Özkan, K.; Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Liboriussen, L.; Svenning, J.C. Contrasting roles of water chemistry, lake morphology, land-use, climate and spatial processes in driving phytoplankton richness in the Danish landscape. Hydrobiologia 2013, 710, 173–187. [Google Scholar] [CrossRef]
- Batanero, G.L.; León-Palmero, E.; Li, L.; Green, A.J.; Rendón-Martos, M.; Suttle, C.A.; Reche, I. Flamingos and drought as drivers of nutrients and microbial dynamics in a saline lake. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Alfonso, M.B.; Zunino, J.; Piccolo, M.C. Impact of water input on plankton temporal dynamics from a managed shallow saline lake. Ann. Limnol. Int. J. Limnol. 2017, 53, 391–400. [Google Scholar] [CrossRef]
- Olmos, J.; Ochoa, L.; Paniagua-Michel, J.; Contreras, R. DNA fingerprinting differentiation between β-carotene hyperproducer strains of Dunaliella from around the world. Saline Syst. 2009, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dolapsakis, N.P.; Tafas, T.; Abatzopoulos, T.J.; Ziller, S.; Economou-Amilli, A. Abundance and growth response of microalgae at Megalon Embolon solar saltworks in northern Greece: An aquaculture prospect. J. Appl. Phycol. 2005, 17, 39–49. [Google Scholar] [CrossRef]
- Heidelberg, K.B.; Nelson, W.C.; Holm, J.B.; Eisenkolb, N.; Andrade, K.; Emerson, J.B. Characterization of eukaryotic microbial diversity in hypersaline Lake Tyrrell, Australia. Front. Microbiol. 2013, 4, 115. [Google Scholar] [CrossRef] [PubMed]
- Moscatello, S.; Belmonte, G. Egg banks in hypersaline lakes of the South-East Europe. Saline Syst. 2009, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Gurevich, P.; Anati, D.A.; Barkan, E.; Luz, B. A bloom of Dunaliella parva in the Dead Sea in 1992: Biological and biogeochemical aspects. Hydrobiologia 1995, 297, 173–185. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- De Deckker, P. Australian salt lakes: Their history, chemistry, and biota—A review. Hydrobiologia 1983, 105, 231–244. [Google Scholar] [CrossRef]
- Mogany, T.; Swalaha, F.M.; Allam, M.; Mtshali, P.S.; Ismail, A.; Kumari, S.; Bux, F. Phenotypic and genotypic characterization of an unique indigenous hypersaline unicellular cyanobacterium, Euhalothece sp. nov. Microbiol. Res. 2018, 211, 47–56. [Google Scholar] [CrossRef]
- Bik, H.M.; Porazinska, D.L.; Creer, S.; Caporaso, J.G.; Knight, R.; Thomas, W.K. Sequencing our way towards understanding global eukaryotic biodiversity. Trends Ecol. Evol. 2012, 27, 233–243. [Google Scholar] [CrossRef]
- Oren, A. A hundred years of Dunaliella research: 1905–2005. Saline Syst. 2005, 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.R.O.B.; Moura, Y.A.S.; Converti, A.; Porto, A.L.F.; Marques, D.D.A.V.; Bezerra, R.P. Assessment of the potential of Dunaliella microalgae for different biotechnological applications: A systematic review. Algal Res. 2021, 58, 102396. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Altitude (m a.s.l.) | Latitude (N) | Longitude (E) | Area (km2) | Catchment Area (km2) |
|---|---|---|---|---|---|
| Aerodromiou 2 | −1.0 | 34.878997 | 33.620442 | 0.31 | 1.84 |
| Akrotiri | −6.0 | 34.619061 | 32.976002 | 11.90 | 91.52 |
| Larnaka Main | −1.6 | 34.894315 | 33.622746 | 4.81 | 58.73 |
| Orfani | −0.5 | 34.860969 | 33.619213 | 2.24 | 10.09 |
| Oroklini | 3.1 | 34.969443 | 33.654534 | 0.15 | 2.77 |
| Soros | −0.5 | 34.845219 | 33.613079 | 0.38 | 10.62 |
| Factor/Lake | Aerodromiou 2 | Akrotiri | Larnaka Main | Orfani | Oroklini | Soros |
|---|---|---|---|---|---|---|
| Artificial surfaces (%) | 83 | 22 | 28 | 45 | 56 | 19 |
| Agricultural areas (%) | 0 | 36 | 57 | 26 | 27 | 77 |
| Forest and semi-natural areas (%) | 0 | 28 | 7 | 7 | 12 | 0 |
| Wetlands | 17 | 14 | 8 | 22 | 5 | 4 |
| Aerodromiou 2 | Akrotiri | Larnaka Main | Orfani | Oroklini | Soros | |
|---|---|---|---|---|---|---|
| BACILLARIOPHYTA (DIATOMS) | ||||||
| Bacillariophyceae | ||||||
| Cylindrotheca closterium (Ehrenberg) Reimann & J.C.Lewin | ● | ● | ○ | ○ | ○ | ○ |
| Fragillaria (Lyngbye) sp. | ○ | ○ | ○ | ○ | ||
| Nitzschia (Hassal) sp. | ● | ○ | ○ | ○ | ● | ● |
| Mediophyceae | ||||||
| Chaetoceros (Ehrenberg) sp. | ○ | ○ | ||||
| Cyclotella [(Kützing) Brébisson] sp. | ● | ● | ○ | ○ | ○ | ○ |
| Unidentified centric diatom | ○ | ○ | ||||
| Unidentified pennate diatom | ○ | ○ | ||||
| CHLOROPHYTA | ||||||
| Chlorodendrophyceae | ||||||
| Tetraselmis (F.Stein) sp. | ○ | |||||
| Chlorophyceae | ||||||
| Ankyra (Fott) sp. | ○ | ○ | ○ | ○ | ||
| Chlamydomonas (Ehrenberg) sp. | ○ | ○ | ● | |||
| Dunaliella (Teodoresco) spp. | ● | ○ | ● | ● | ● | ● |
| Kirchneriella (Schmidle) sp. | ○ | ○ | ||||
| Monoraphidium (Komárková-Legnerová) sp. | ○ | ○ | ||||
| Schroederia (Lemmermann) sp. | ○ | ○ | ● | ○ | ● | ○ |
| Pyramimonadophyceae | ||||||
| Pseudoscourfieldia marina (J.Throndsen) Manton | ● | ● | ● | ● | ● | |
| Trebouxiophyceae | ||||||
| Crucigeniella (Lemmermann) sp. | ○ | |||||
| Didymocystis (Korshikov) sp. | ● | |||||
| Oocystis (Nägeli ex A. Braun) sp. | ● | ○ | ||||
| Planctonema lauterbornii (Schmidle) | ○ | |||||
| Unidentified coccoid microeukaryotes | ○ | ● | ● | ○ | ● | ○ |
| CRYPTISTA | ||||||
| Cryptophyceae | ||||||
| Rhodomonas (G.Karsten) sp. | ○ | |||||
| Cryptomonas (Ehrenberg) sp. | ○ | ○ | ○ | ● | ● | ○ |
| CYANOBACTERIA | ||||||
| Cyanophyceae | ||||||
| Anabaena (Bory ex Bornet & Flahault) sp. | ○ | ○ | ○ | ○ | ○ | |
| Aphanizomenon (Morren ex Bornet & Flahault) sp. | ○ | ○ | ○ | |||
| Chroococcus (Nägeli) sp. | ○ | ○ | ○ | |||
| Jaagenima? (Anagnostidis & Komarek) sp. | ○ | ○ | ○ | ○ | ○ | ○ |
| Euhalothece (F.Garcia-Pichel, U.Nübel & G.Muyzer, 1998, non. inval.) sp. | ● | |||||
| Oscillatoria (Vaucher ex Gomont) sp. | ○ | ○ | ○ | ○ | ○ | |
| Planktothrix (Anagnostidis & Komárek) sp. | ○ | |||||
| Pseudanabaena (Lauterborn) sp. | ○ | ○ | ○ | ○ | ● | ○ |
| Unidentified coccoid cyanobacteria | ● | ● | ||||
| EUGLENOZOA | ||||||
| Euglenophyceae | ||||||
| Euglena (Ehrenberg) sp. | ○ | ● | ○ | |||
| Unidentified euglenophytes | ○ | |||||
| HAPTOPHYTA | ||||||
| Coccolithophyceae | ||||||
| Prymnesium parvum (N.Carter) | ● | |||||
| CHAROPHYTA | ||||||
| Closteriaceae | ||||||
| Closterium (Nitzsch ex Ralfs) sp. | ○ | |||||
| MIOZOA | ||||||
| Dinophyceae | ||||||
| Amphidinium (Claperède & Lachmann) sp. | ○ | ○ | ||||
| Gymnodynium (F.Stein) sp. | ● | ○ | ○ | ○ | ○ | |
| Peridiniella (C.A.Kofoid & J.R.Michener) sp. | ● | ○ | ○ | ○ | ○ | |
| Peridinium (Ehrenberg) sp. | ○ | ○ | ||||
| Unidentified dinoflagellate | ● |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polykarpou, P.; Katsiapi, M.; Genitsaris, S.; Stefanidou, N.; Dörflinger, G.; Moustaka-Gouni, M.; Economou-Amilli, A.; Raitsos, D.E. Phytoplankton Diversity and Blooms in Ephemeral Saline Lakes of Cyprus. Diversity 2023, 15, 1204. https://doi.org/10.3390/d15121204
Polykarpou P, Katsiapi M, Genitsaris S, Stefanidou N, Dörflinger G, Moustaka-Gouni M, Economou-Amilli A, Raitsos DE. Phytoplankton Diversity and Blooms in Ephemeral Saline Lakes of Cyprus. Diversity. 2023; 15(12):1204. https://doi.org/10.3390/d15121204
Chicago/Turabian StylePolykarpou, Polina, Matina Katsiapi, Savvas Genitsaris, Natassa Stefanidou, Gerald Dörflinger, Maria Moustaka-Gouni, Athena Economou-Amilli, and Dionysios E. Raitsos. 2023. "Phytoplankton Diversity and Blooms in Ephemeral Saline Lakes of Cyprus" Diversity 15, no. 12: 1204. https://doi.org/10.3390/d15121204