
Temporal Variation of Genetic Diversity in Rutilus rutilus Populations from Lithuania Using mtDNA Markers in the Context of Anthropogenic Activities

Abstract
:1. Introduction
2. Materials and Methods
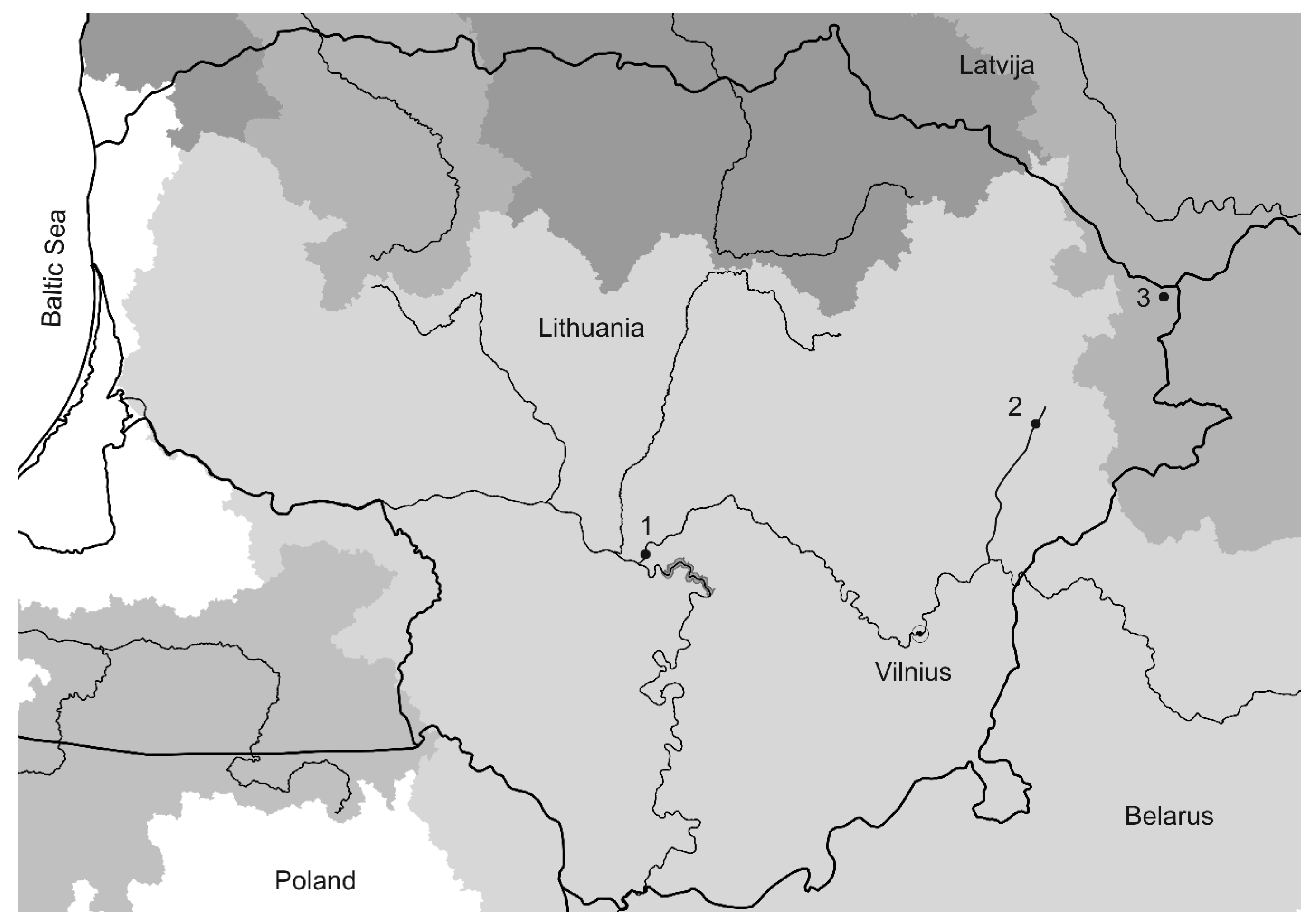
2.1. Sampling, DNA Extraction
2.2. PCR Conditions and Sequencing
2.3. Molecular Data Analysis
3. Results
3.1. Sequence Variation and Genetic Diversity
3.2. Haplotype Network Analysis
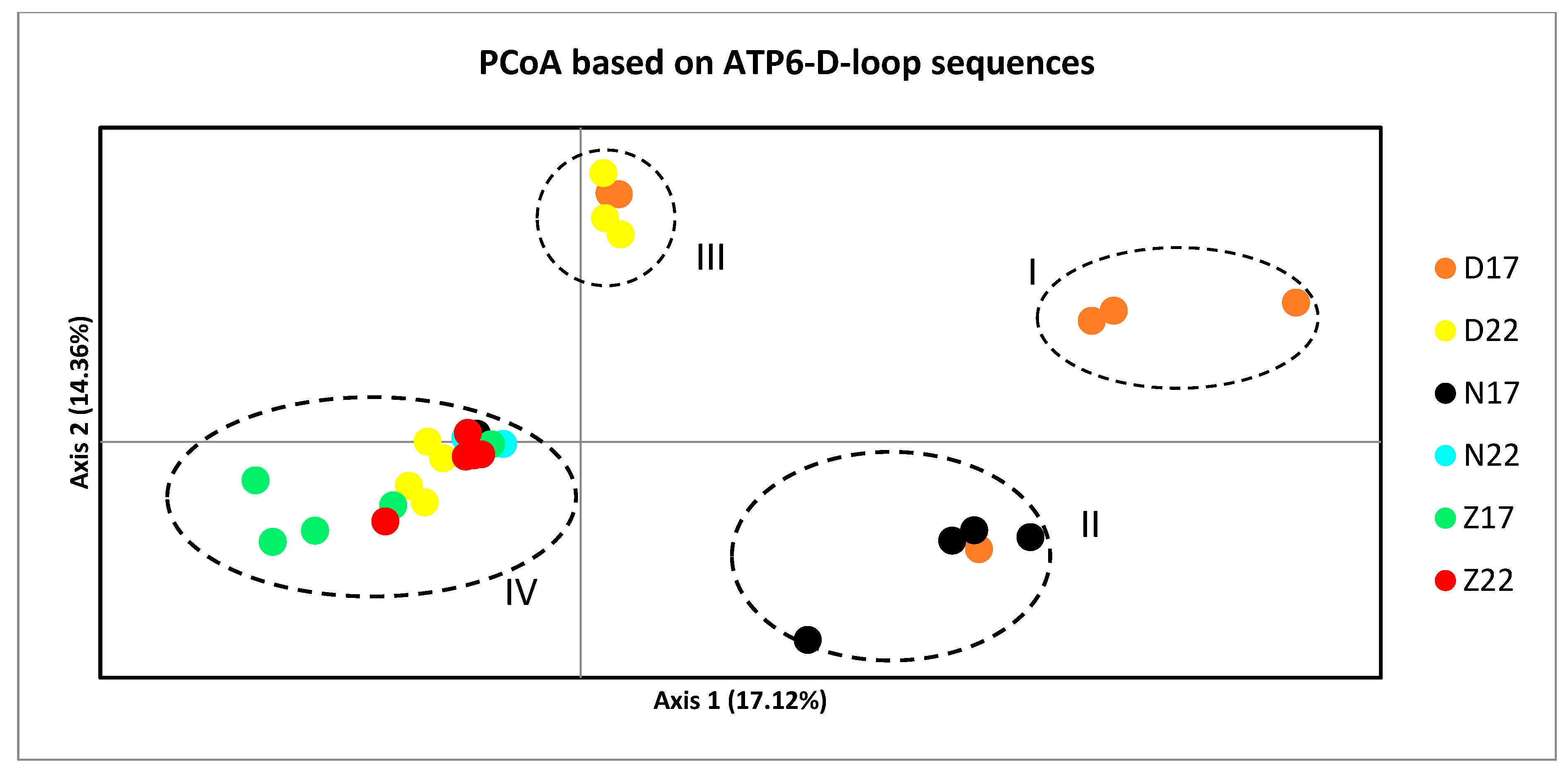
3.3. Population Genetic Structure
4. Discussion
4.1. Genetic Diversity
4.2. Population Genetic Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nater, A.; Arora, N.; Greminger, M.P.; van Schaik, C.P.; Singleton, I. Marked population structure and recent migration in the critically endangered Sumatran orangutan (Pongo abelii). J. Hered. 2013, 104, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar] [CrossRef]
- Baranyi, C.; Gollmann, G.; Bobin, M. Genetic and morphological variability in roach Rutilus rutilus, from Austria. Hydrobiologia 1997, 350, 13–23. [Google Scholar] [CrossRef]
- Raeymaekers, J.A.; Maes, G.E.; Geldof, S.; Hontis, I.; Nackaerts, K.; Volckaert, F.A. Modeling genetic connectivity in sticklebacks as a guideline for river restoration. Evol. Appl. 2008, 1, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Yamamoto, S. Effects of habitat fragmentation by damming on the persistence of stream-dwelling charr populations. Conserv. Biol. 2002, 16, 1318–1323. [Google Scholar] [CrossRef]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef] [PubMed]
- Coulter, D.P.; Sepúlveda, M.S.; Troy, C.D.; Höök, T.O. Thermal habitat quality of aquatic organisms near power plant discharges: Potential exacerbating effects of climate warming. Fish. Manag. Ecol. 2014, 21, 196–210. [Google Scholar] [CrossRef]
- Langford, T.E. Thermal discharges and pollution. In Encyclopedia of Ocean Sciences; Academic Press: Cambridge, MA, USA, 2001; pp. 2933–2940. [Google Scholar] [CrossRef]
- Dokulil, M.T. Impact of climate warming on European inland waters. Inland Waters 2013, 4, 27–40. [Google Scholar] [CrossRef]
- Hester, E.T.; Doyle, M.W. Human impacts to river temperature and their effects on biological processes: A quantitative synthesis. J. Am. Water Resour. Assoc. 2011, 47, 571–587. [Google Scholar] [CrossRef]
- Borell, Y.J.; Bernardo, D.; Blanco, G.; Vásquez, E.; Sánchez, J.A. Spatial and temporal variation of genetic diversity and estimation of effective population sizes in Atlantic salmon (Salmo salar, L.) populations from Asturias (Northern Spain) using microsatellites. Conserv. Gen. 2008, 9, 807–819. [Google Scholar] [CrossRef]
- Klerks, P.L.; Levinton, J.S. Rapid evolution of metal resistance in a benthic oligochaete inhabiting a metal-polluted site. Biol. Bull. 1989, 176, 135–141. [Google Scholar] [CrossRef]
- Oziolor, E.M.; Bigorgne, E.; Aguilar, L.; Usenko, S.; Matson, C.W. Evolved resistance to PCB- and PAH-induced cardiac teratogenesis, and reduced CYP1A activity in Gulf killifish (Fundulus grandis) populations from the Houston Ship Channel, Texas. Aquat. Toxicol. 2014, 150, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Reid, N.M.; Proestou, D.A.; Clark, B.W.; Warren, W.C.; Colbourne, J.K.; Shaw, J.R.; Karchner, S.I.; Hahn, M.E.; Nacci, D.; Oleksiak, M.F.; et al. The genomic landscape of rapid repeated evolutionary adaptation to toxic pollution in wild fish. Science 2016, 354, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Staton, S.C.; Cheviron, Z.A.; Rochette, N.; Catchen, J.; Losos, J.B.; Edwards, S.V. Winter storms drive rapid phenotypic, regulatory, and genomic shifts in the green anole lizard. Science 2017, 357, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Hermoso, V.; Clavero, M. Threatening processes and conservation management of endemic freshwater fish in the Mediterranean basin: A review. Mar. Freshw. Res. 2011, 62, 244–254. [Google Scholar] [CrossRef]
- Movchan, Y.V.; Smirnov, A.I. Fauna of Ukraine. Fishes. Cyprinid Fishes (Roach, Dace, Minnow, Rudd, Grass Carp, Asp, Verchovka, Tench, Undermouth, Gudgeon, Barbel). (Fauna Ukrainy. Ryby. Koropovi (Plitka, Yalets, Golijan, Krasnopirka, Amur, Bilyzna, Verkhova, Lyn, Chebachok Amurskyi); Kiev Naukova Dumka Publishing House: Kyiv, Ukraine, 1981; Volume 8, Part 1. [Google Scholar]
- Svanbäck, R.; Eklöv, P.; Fransson, R.; Holmgren, K. Intraspecific competition drives multiple species resource polymorphism in fish communities. Oikos 2008, 117, 114–124. [Google Scholar] [CrossRef]
- Virbickas, J. Lietuvos žuvys (Fishes of Lithuania); Ekologijos Institutas: Vilnius, Lithuania, 2000. [Google Scholar]
- Kawamura, K. “Handbook of European freshwater fishes” by M. Kottelat and J. Freyhof (2007). Ichthyol. Res. 2008, 55, 99. [Google Scholar] [CrossRef]
- Lappalainen, A.; Rask, M.; Koponen, H.; Vesala, S. Relative abundance, diet and growth of perch (Perca fluviatilis) and roach (Rutilus rutilus) at Tvärminne, northern Baltic Sea, in 1975 and 1997: Responses to eutrophication? Boreal Env. Res 2001, 6, 107–108. [Google Scholar]
- Faulks, L.; Svanbäck, R.; Eklöv, P.; Östman, Ö. Genetic and morphological divergence along the littoral–pelagic axis in two common and sympatric fishes: Perch, Perca fluviatilis (Percidae) and roach, Rutilus rutilus (Cyprinidae). Biol. J. Linn. Soc. 2015, 114, 929–940. [Google Scholar] [CrossRef]
- Crookes, S.; Shaw, P.W. Isolation by distance and non-identical patterns of gene flow within two river populations of the freshwater fish Rutilus rutilus (L. 1758). Conserv. Genet. 2016, 17, 861–874. [Google Scholar] [CrossRef]
- Larmuseau, M.H.D.; Freyhof, J.; Volckaert, F.A.M.; Van Houdt, J.K.J. Matrilinear phylogeography and demographical patterns of Rutilus rutilus: Implications for taxonomy and conservation. J. Fish Biol. 2009, 75, 332–353. [Google Scholar] [CrossRef]
- Nedveckaite, T.; Motiejunas, S.; Kucinskas, V.; Mazeika, J.; Filistovic, V.; Jusciene, D.; Maceika, E.; Morkeliunas, L.; Hamby, D.M. Environmental releases of radioactivity and the incidence of thyroid disease at the Ignalina Nuclear Power Plant. Health Phys. 2000, 79, 666–674. [Google Scholar] [CrossRef]
- Thoresson, G. Guidelines for Coastal Monitoring. Kustrapport 1. National Board of Fisheries; Institute of Coastal Research: Öregrund, Sweden, 1993. [Google Scholar]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Sánchez-Delbarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef] [PubMed]
- Polzin, T.S.; Daneschmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef]
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations agai nst population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Tajima, F.V. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under linux and windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Petit-Marty, N.; Vázquez-Luis, M.; Hendriks, I.E. Use of the Nucleotide Diversity in COI Mitochondrial Gene as an Early Diagnostic of Conservation Status of Animal Species. Conserv. Lett. 2021, 14, e12756. [Google Scholar] [CrossRef]
- Demandt, M.H. Temporal changes in genetic diversity of isolated populations of perch and roach. Conserv. Genet. 2010, 11, 249–255. [Google Scholar] [CrossRef]
- Hamilton, P.B.; Nicol, E.; De-Bastos, E.S.; Williams, R.J.; Sumpter, J.P.; Jobling, S.; Stevens, J.R.; Tyler, C.R. Populations of a cyprinid fish are self-sustaining despite widespread feminization of males. BMC Biol. 2014, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Ryota, Y.; Akira, G. Phylogeography of a freshwater Sculpin, Cottusnozawae from the northeastern part of Honshu Island, Japan. Ichthyol. Res. 2002, 49, 147–155. [Google Scholar] [CrossRef]
- Pourkazemi, M.; Skibinski, D.O.F.; Beardmore, J. Application of mtDNA d-loop region for the study of Russian sturgeon population structure from Iranian coastline of the Caspian Sea. J. Appl. Ichthyol. 1999, 15, 23–28. [Google Scholar] [CrossRef]
- Iervolino, F.; de Resende, E.K.; Hilsdorf, A.W.S. The lack of genetic differentiation of pacu (Piaractus mesopotamicus) populations in the Upper-Paraguay Basin revealed by the mitochondrial DNA D-loop region: Implications for fishery management. Fish. Res. 2010, 101, 27–31. [Google Scholar] [CrossRef]
- Padhi, A. Geographic variation within a tandemly repeated mitochondrial DNA D-loop region of a North American freshwater fish, Pylodictis olivaris. Gene 2014, 538, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Guo, X.; Liu, S.; Xiao, J.; Liu, Z.; Chen, Y.; Liu, Y. Maternal inheritance in polyploid fish inferred from mitochondrial ATPase genes analysis. Prog. Nat. Sci. 2009, 19, 693–698. [Google Scholar] [CrossRef]
- Garcia, D.A.; Lasso, C.A.; Morales, M.; Caballero, S.J. Molecular systematics of the freshwater stingrays (myliobatiformes: Potamotrygonidae) of the Amazon, Orinoco, Magdalena, Esequibo, Caribbean, and Maracaibo basins (Colombia–Venezuela): Evidence from three mitochondrial genes. Mitochondrial DNA Part A 2016, 27, 4479–4491. [Google Scholar] [CrossRef]
- Baltazar-Soares, M.; de Araújo Lima, A.R.; Silva, G. Targeted Sequencing of Mitochondrial Genes Reveals Signatures of Molecular Adaptation in a Nearly Panmictic Small Pelagic Fish Species. Genes 2021, 12, 91. [Google Scholar] [CrossRef]
- Yan, T.; Wang, X.; Li, S.; He, Z.; Luo, J.; Zhang, Q.; Yang, D.; He, Z. Genetic analysis of wild Ancherythroculter nigrocauda in tributaries and the main stream of the upper Yangtze River basin of China. Mitochondrial DNA A 2020, 31, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Jager, H.I.; Chandler, J.A.; Lepla, K.B.; Winkle, W.V. A theoretical study of river fragmentation by dams and its effects on white sturgeon populations. Environ. Biol. Fishes 2001, 60, 347–361. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography, the History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Allan, J.D.; Abell, R.; Hogan, Z.; Revenga, C.; Taylor, B.W.; Welcomme, R.L.; Winemiller, K. Overfishing of inland waters. BioScience 2005, 55, 1041–1051. [Google Scholar] [CrossRef]
- Ragauskas, A.; Butkauskas, D.; Sruoga, A. Investigation into genetic diversity of perch inhabiting Lake Drūkšiai and other water bodies of Lithuania on the basis of mtDNA analysis. Zool. Ecol. 2014, 24, 154–159. [Google Scholar] [CrossRef]
- Yang, L.; Hu, J.; Qin, C.; Zhang, Y.; Lu, R.; Meng, X.; Yang, G.; Yan, X.; Zhi, S.; Nie, G. Genetic structure analysis of Pseudorasbora parva in the four major river systems in Yunnan based on mitochondrial Cyt b. JFC 2020, 3, 339–350. [Google Scholar] [CrossRef]
- Soliman, T.; Aly, W.; Fahim, R.M.; Berumen, M.L.; Jenke-Kodama, H.; Bernardi, G. Comparative population genetic structure of redbelly tilapia (Coptodon zillii (Gervais, 1848)) from three different aquatic habitats in Egypt. Ecol. Evol. 2017, 7, 11092–11099. [Google Scholar] [CrossRef] [PubMed]
- Goldspink, C.R. The return of marked roach (Rutilus rutilus L.) to spawning grounds in Tjeukemeer, The Netherlands. J. Fish Biol. 1977, 11, 599–603. [Google Scholar] [CrossRef]
- Rowe, G.; Sweet, M.; Beebee, T.J.C. An Introduction to Molecular Ecology; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Zhang, F.; Duan, Y.; Cao, S.; Wang, J.; Tan, D. High genetic diversity in population of Lepturichthys fimbriata from the Yangtze River revealed by microsatellite DNA analysis. Chin. Sci. Bull. 2012, 57, 487–491. [Google Scholar] [CrossRef]
- Meldgaard, T.; Nielsen, E.E.; Loeschcke, V. Fragmentation by weirs in a riverine system: A study of genetic variation in time and space among populations of European grayling (Thymallus thymallus) in a Danish river system. Conserv. Genet. 2003, 4, 735–747. [Google Scholar] [CrossRef]
- Argüelles, R.; Toledo, M.; Martín, M.A. Study of the Tagus River and Entrepeñas reservoir ecosystem around the Trillo nuclear power plant using chemometric analysis: Influence on water, sediments, algae and fish. Chemosphere 2021, 279, 130532. [Google Scholar] [CrossRef]
- Rettman, A. Lithuania Nuclear Shutdown to Test EU-Russia Relations. Available online: EUobserver.com (accessed on 31 December 2009).
- Björn, H. Science and Politics of Foreign Aid: Swedish Environmental Support to the Baltic States; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003. [Google Scholar] [CrossRef]
- Mažeika, J.; Taminskas, J. Water system of the lake Druksiai Transboundary Catchment under anthropogenic pressure. Pol. Geol. Inst. Spec. Pap. 2005, 18, 71–78. [Google Scholar]
- Verones, F.; Hanafiah, M.M.; Pfister, S.; Huijbregts, M.A.J.; Pelletier, G.J.; Koehler, A. Characterization factors for thermal pollution in freshwater aquatic environments. Environ. Sci. Technol. 2010, 44, 9364–9369. [Google Scholar] [CrossRef] [PubMed]
- Astrauskas, A.; Bernotas, E.; Didrikas, T. The impacts of the Ignalina Nuclear Power Plant effluent on fishes in Lithuania. Ital. J. Zool. 1998, 65, 461–464. [Google Scholar] [CrossRef]
- Kesminas, V.; Olechnovičienė, J. Fish community changes in the cooler of the Ignalina Nuclear Power Plant. Ekologija 2008, 54, 124–131. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity: The search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef]
- Dowle, E.J.; Morgan-Richards, M.; Trewick, S.A. Molecular evolution and the latitudinal biodiversity gradient. Heredity 2013, 110, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.M. A latitudinal gradient for genetic diversity. Science 2016, 353, 1494–1495. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Locations | Collection Date | Population Code | Number of Samples | Location |
|---|---|---|---|---|
| Lake Drūkšiai | 2017 | D17 | 19 | 55°38′45.7″ N 26°35′53.3″ E |
| 2022 | D22 | 15 | ||
| Neris River | 2017 | N17 | 11 | 54°57′24.8″ N 23°55′41.9″ E |
| 2022 | N22 | 12 | ||
| Žeimena River | 2017 | Z17 | 12 | 55°14′26.6″ N 25°58′48.3″ E |
| 2022 | Z22 | 15 |
| Population Code | h | h’ | Hd | S | π | K | Neutrality Test | ||
|---|---|---|---|---|---|---|---|---|---|
| Tajima’s D | Fu’s Fs | ||||||||
| D-loop | D17 | 10 | 6 | 0.8245 | 8 | 0.0038 | 1.8011 | −0.7288 | −5.3827 ** |
| D22 | 7 | 4 | 0.8761 | 4 | 0.0025 | 1.6190 | 0.8942 | −2.5570 * | |
| N17 | 7 | 4 | 0.8727 | 6 | 0.0033 | 1.5636 | −0.9370 | −3.7031 ** | |
| N22 | 6 | 2 | 0.6818 | 7 | 0.0030 | 1.4090 | −1.5344 | −2.2953 * | |
| Z17 | 4 | 2 | 0.7424 | 4 | 0.0027 | 1.2575 | −0.1778 | −0.1272 | |
| Z22 | 4 | 2 | 0.4667 | 3 | 0.0011 | 0.5142 | −1.3165 | −1.7972 * | |
| Total | 26 | 20 | 0.7938 | 14 | 0.0033 | 1.5140 | −0.6334 | −2.6438 | |
| ATP6 | D17 | 5 | 4 | 0.3859 | 4 | 0.0009 | 0.4210 | −1.8612 * | −3.5705 * |
| D22 | 2 | 0 | 0.3428 | 1 | 0.0007 | 0.3428 | 0.2350 | 0.5966 | |
| N17 | 3 | 1 | 0.4727 | 2 | 0.0011 | 0.5090 | −0.7781 | −0.6587 | |
| N22 | 2 | 0 | 0.3030 | 1 | 0.0006 | 0.3030 | −0.1949 | 0.2973 | |
| Z17 | 2 | 1 | 0.1666 | 1 | 0.0003 | 0.1666 | −1.1405 | −0.4756 | |
| Z22 | 3 | 1 | 0.2571 | 2 | 0.0005 | 0.2666 | −1.4905 * | −1.5463 * | |
| Total | 9 | 7 | 0.3207 | 8 | 0.0008 | 0.3445 | −0.8717 | −0.8928 * | |
| ATP6-D-loop | D17 | 12 | 8 | 0.8713 | 12 | 0.0024 | 2.2222 | −1.2836 | −7.4378 ** |
| D22 | 8 | 6 | 0.9142 | 5 | 0.0016 | 1.9619 | 0.8148 | −3.1514 * | |
| N17 | 7 | 4 | 0.8727 | 7 | 0.0019 | 1.7818 | −1.0325 | −3.2539 * | |
| N22 | 6 | 2 | 0.8181 | 5 | 0.0013 | 1.2424 | −0.9201 | −2.6663 ** | |
| Z17 | 7 | 6 | 0.9090 | 7 | 0.0024 | 2.2727 | −0.0763 | −2.1626 | |
| Z22 | 6 | 3 | 0.6476 | 7 | 0.0011 | 1.0476 | −1.8487 * | −2.6661 * | |
| Total | 35 | 29 | 0.9036 | 26 | 0.0022 | 2.0981 | −0.7244 | −3.5564 * | |
| Sequence | Source of Variation | d.f. | Sum of Squares | Percentage of Variance, % | PhiPT |
|---|---|---|---|---|---|
| D-loop | |||||
| Whole | Among populations | 5 | 13.681 | 18 | 0.177 *** |
| Within populations | 78 | 53.629 | 82 | ||
| Total | 83 | 67.310 | 100 | ||
| 2017-year group | Among populations | 2 | 6.126 | 17 | 0.174 *** |
| Within populations | 39 | 30.945 | 83 | ||
| Total | 41 | 37.071 | 100 | ||
| 2022-year group | Among populations | 2 | 3.531 | 13 | 0.128 *** |
| Within populations | 39 | 22.683 | 87 | ||
| Total | 41 | 26.214 | 100 | ||
| ATP6 | |||||
| Whole | Among populations | 5 | 1.113 | 2 | 0.022 |
| Within populations | 78 | 13.185 | 98 | ||
| Total | 83 | 14.298 | 100 | ||
| 2017-year group | Among populations | 2 | 0.510 | 3 | 0.027 |
| Within populations | 39 | 7.252 | 97 | ||
| Total | 41 | 7.762 | 100 | ||
| 2022-year group | Among populations | 2 | 0.186 | 0 | −0.029 |
| Within populations | 39 | 5.933 | 100 | ||
| Total | 41 | 6.119 | 100 | ||
| ATP6-D-loop | |||||
| Whole | Among populations | 5 | 17.762 | 18 | 0.177 *** |
| Within populations | 78 | 69.309 | 82 | ||
| Total | 83 | 87.071 | 100 | ||
| 2017-year group | Among populations | 2 | 8.615 | 18 | 0.184 *** |
| Within populations | 39 | 41.409 | 82 | ||
| Total | 41 | 50.024 | 100 | ||
| 2022-year group | Among populations | 2 | 3.862 | 11 | 0.109 *** |
| Within populations | 39 | 27.900 | 89 | ||
| Total | 41 | 31.762 | 100 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butkauskas, D.; Ignatavičienė, I.; Ragauskas, A.; Rakauskas, V. Temporal Variation of Genetic Diversity in Rutilus rutilus Populations from Lithuania Using mtDNA Markers in the Context of Anthropogenic Activities. Diversity 2023, 15, 1113. https://doi.org/10.3390/d15111113
Butkauskas D, Ignatavičienė I, Ragauskas A, Rakauskas V. Temporal Variation of Genetic Diversity in Rutilus rutilus Populations from Lithuania Using mtDNA Markers in the Context of Anthropogenic Activities. Diversity. 2023; 15(11):1113. https://doi.org/10.3390/d15111113
Chicago/Turabian StyleButkauskas, Dalius, Ieva Ignatavičienė, Adomas Ragauskas, and Vytautas Rakauskas. 2023. "Temporal Variation of Genetic Diversity in Rutilus rutilus Populations from Lithuania Using mtDNA Markers in the Context of Anthropogenic Activities" Diversity 15, no. 11: 1113. https://doi.org/10.3390/d15111113