
Dispersal Capabilities Do Not Parallel Ecology and Cryptic Speciation in European Cheliferidae Pseudoscorpions (Pseudoscorpiones: Cheliferidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxonomic Sampling
2.2. DNA Extraction and PCR Amplification
2.3. Phylogenetic Analyses
2.4. Molecular Species Delimitation
3. Results
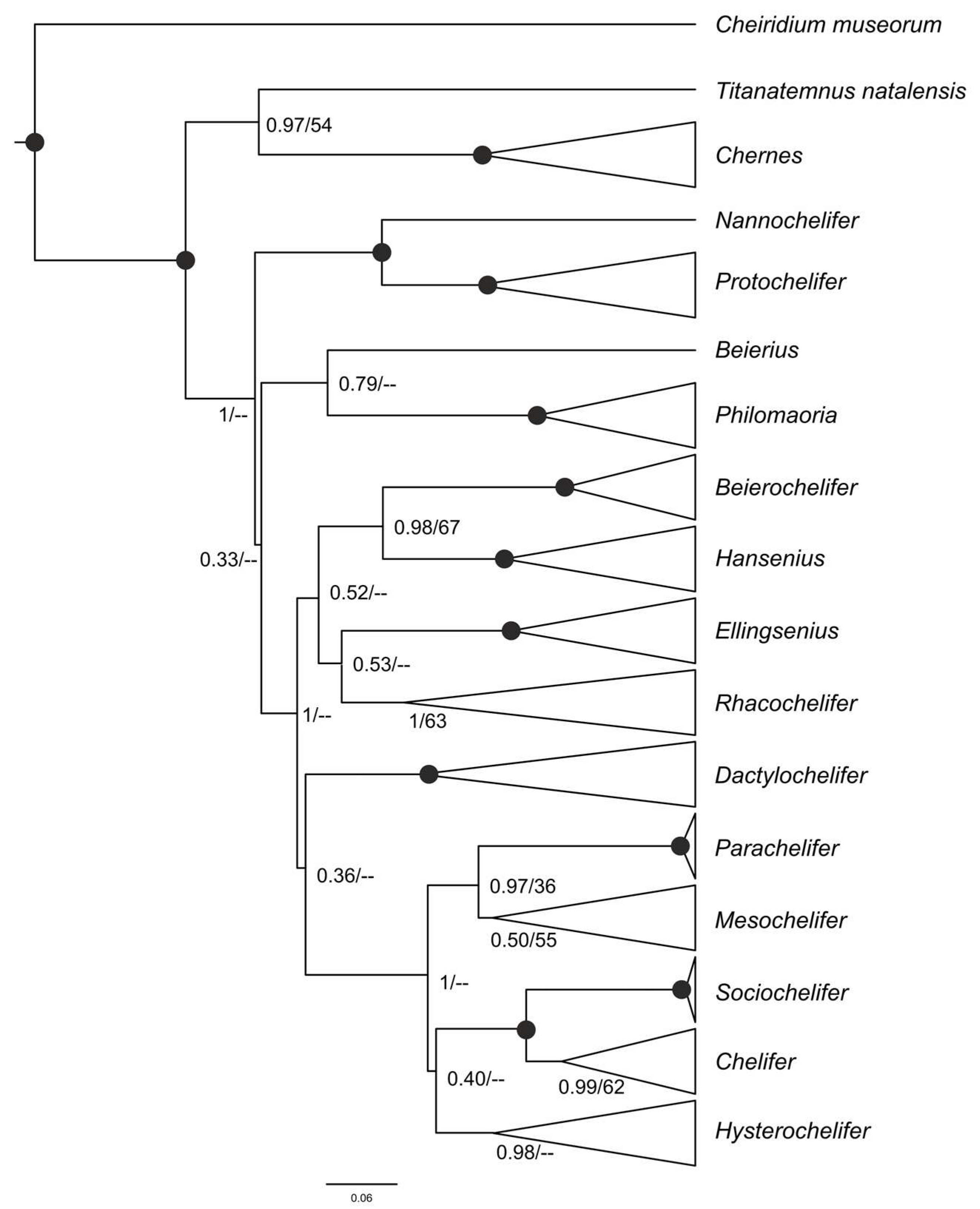
3.1. Taxonomic Sampling and Phylogenetic Analyses
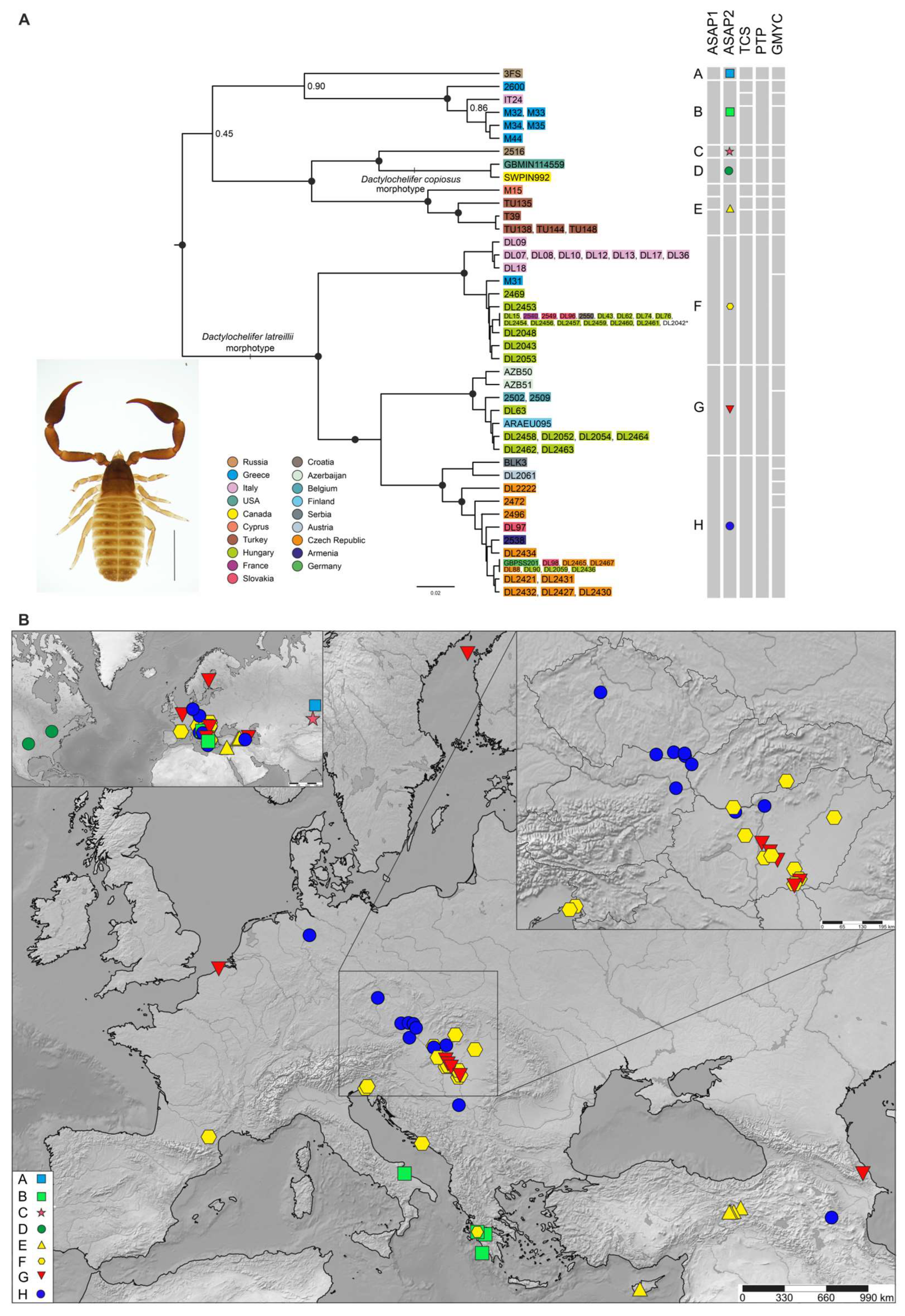
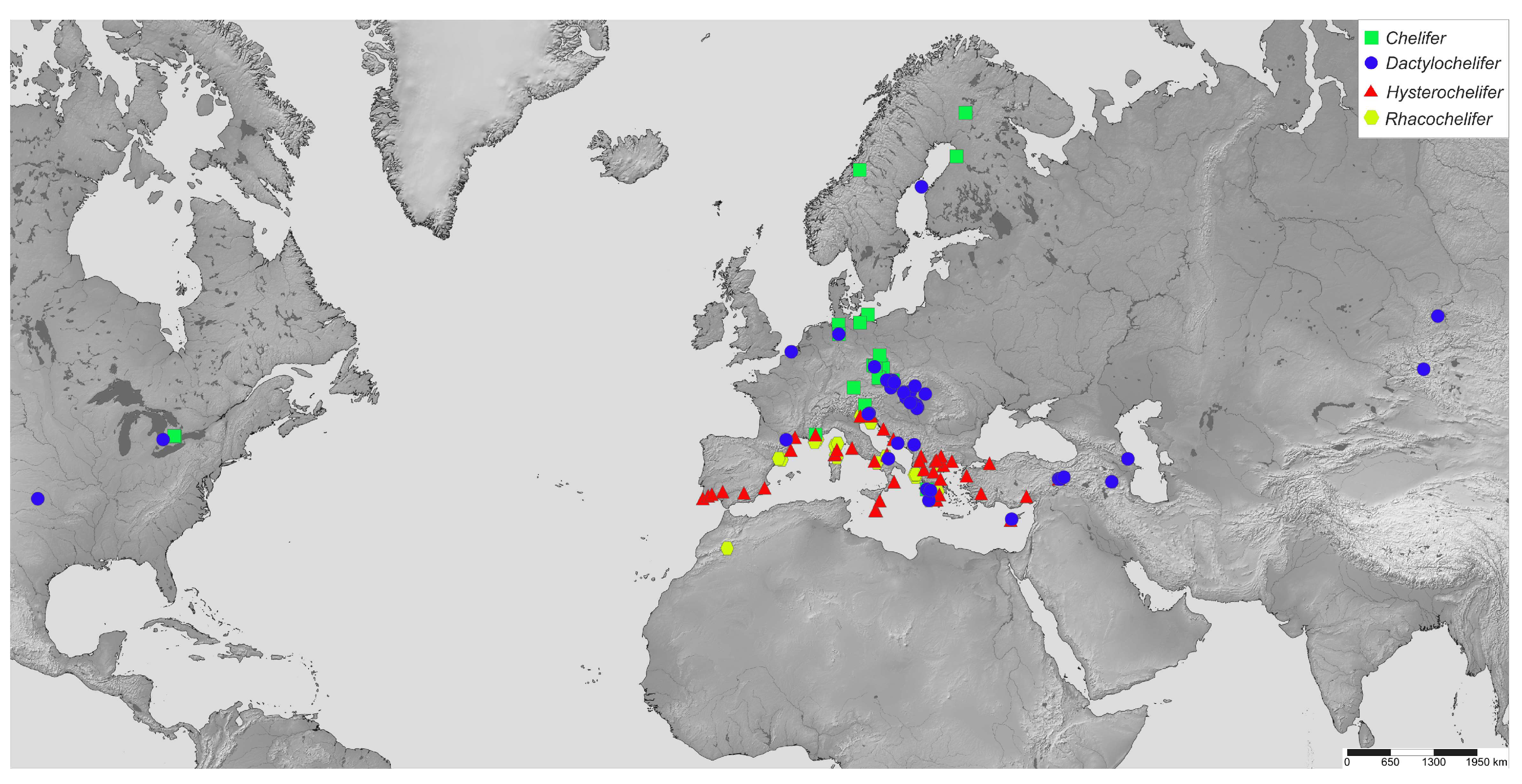
3.2. Molecular Species Delimitation and Geographic Distribution of the Lineages
3.2.1. Chelifer
3.2.2. Dactylochelifer
3.2.3. Hysterochelifer



3.2.4. Rhacochelifer
4. Discussion
4.1. Phylogenetic Relationships among Cheliferidae
4.2. Cheliferidae Diversity and Delimitation
4.2.1. Chelifer cancroides
4.2.2. Dactylochelifer
4.2.3. Hysterochelifer
4.2.4. Rhacochelifer
4.3. Effects of Dispersal Strategies on Distribution and Diversity of Cheliferids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacArthur, R. Patterns of species diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Wilson, R.J.; Thomas, C.D.; Fox, R.; Roy, D.B.; Kunin, W.E. Spatial patterns in species distributions reveal biodiversity change. Nature 2004, 432, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Arbogast, B.S.; Kenagy, G.J. Comparative phylogeography as an integrative approach to historical biogeography. J. Biogeogr. 2001, 28, 819–825. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Barrow, L.N.; Lemmon, A.R.; Lemmon, E.M. Targeted sampling and target capture: Assessing phylogeographic concordance with genome-wide data. Syst. Biol. 2018, 67, 979–996. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.L.; Gaines, M.S. Evolution of dispersal: Theoretical models and empirical tests using birds and mammals. Annu. Rev. Ecol. Syst. 1990, 21, 449–480. [Google Scholar] [CrossRef]
- Nathan, R. Long-distance dispersal of plants. Science 2006, 313, 786–788. [Google Scholar] [CrossRef] [PubMed]
- Opatova, V.; Hamilton, C.A.; Hedin, M.; De Oca, L.M.; Král, J.; Bond, J.E. Phylogenetic systematics and evolution of the spider infraorder Mygalomorphae using genomic scale data. Syst. Biol. 2020, 69, 671–707. [Google Scholar] [CrossRef]
- Travis, J.M.; Dytham, C. Dispersal and climate change: A review of theory. In Dispersal Ecology and Evolution; Oxford University Press: Oxford, UK, 2012; pp. 337–348. [Google Scholar]
- Bartlow, A.W.; Agosta, S.J. Phoresy in animals: Review and synthesis of a common but understudied mode of dispersal. Biol. Rev. 2021, 96, 223–246. [Google Scholar] [CrossRef]
- Szymkowiak, P.; Górski, G.; Bajerlein, D. Passive dispersal in arachnids. Biol. Lett. 2007, 44, 75–101. [Google Scholar]
- Fontaneto, D. Long-distance passive dispersal in microscopic aquatic animals. Mov. Ecol. 2019, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Opatova, V.; Šťáhlavský, F. Phoretic or not? Phylogeography of the pseudoscorpion Chernes hahnii (Pseudoscorpiones: Chernetidae). J. Arachnol. 2018, 46, 104–113. [Google Scholar] [CrossRef]
- Ohnishi, O.; Takenaka, M.; Okano, R.; Yoshitomi, H.; Tojo, K. Wide-scale gene flow, even in insects that have lost their flight ability: Presence of dispersion due to a unique parasitic ecological strategy of piggybacking hosts. Zool. Sci. 2021, 38, 122–139. [Google Scholar] [CrossRef] [PubMed]
- López-Estrada, E.K.; Sanmartín, I.; Uribe, J.E.; Abalde, S.; Jiménez-Ruiz, Y.; García-París, M. Mitogenomics and hidden-trait models reveal the role of phoresy and host shifts in the diversification of parasitoid blister beetles (Coleoptera: Meloidae). Mol. Ecol. 2022, 31, 2453–2474. [Google Scholar] [CrossRef] [PubMed]
- Christophoryová, J.; Krajčovičová, K.; Šťáhlavský, F.; Španiel, S.; Opatova, V. Integrative Taxonomy Approach Reveals Cryptic Diversity within the Phoretic Pseudoscorpion Genus Lamprochernes (Pseudoscorpiones: Chernetidae). Insects 2023, 14, 122. [Google Scholar] [CrossRef] [PubMed]
- World Pseudoscorpiones Catalog. Available online: http://wac.nmbe.ch (accessed on 4 September 2023).
- Harvey, M.S. The smaller arachnid orders: Diversity, descriptions and distributions from Linnaeus to the present (1758 to 2007). Zootaxa 2007, 1668, 363–380. [Google Scholar] [CrossRef]
- Weygoldt, P. Biology of Pseudoscorpions; Harvard University Press: Cambridge, MA, USA, 1969. [Google Scholar]
- Legg, G. The possible significance of spermathecae in pseudoscorpions (Arachnida). Bull. Br. Arach. Soc. 1975, 3, 91–95. [Google Scholar]
- Muster, C.; Spelda, J.; Rulik, B.; Thormann, J.; von der Mark, L.; Astrin, J.J. The dark side of pseudoscorpion diversity: The German Barcode of Life campaign reveals high levels of undocumented diversity in European false scorpions. Ecol. Evol. 2021, 11, 13815–13829. [Google Scholar] [CrossRef]
- Zaragoza, J.A.; Šťáhlavský, F. A new Roncus species (Pseudoscorpiones: Neobisiidae) from Montseny Natural Park (Catalonia, Spain), with remarks on karyology. Zootaxa 2008, 1693, 27–40. [Google Scholar] [CrossRef]
- Kotrbová, J.; Opatova, V.; Gardini, G.; Šťáhlavský, F. Karyotype diversity of pseudoscorpions of the genus Chthonius (Pseudoscorpiones, Chthoniidae) in the Alps. Comp. Cytogenet. 2016, 10, 325–345. [Google Scholar]
- Zaragoza, J.A. Revision of the Ephippiochthonius complex in the Iberian Peninsula, Balearic Islands and Macaronesia, with proposed changes to the status of the Chthonius subgenera (Pseudoscorpiones, Chthoniidae). Zootaxa 2017, 4246, 1–221. [Google Scholar] [CrossRef] [PubMed]
- Hlebec, D.; Podnar, M.; Kučinić, M.; Harms, D. Molecular analyses of pseudoscorpions in a subterranean biodiversity hotspot reveal cryptic diversity and microendemism. Sci. Rep. 2023, 13, 430. [Google Scholar] [CrossRef] [PubMed]
- Colado, R.; García-Meseguer, A.J.; Mirón-Gatón, J.M.; Botella-Cruz, M.; Pallarés, S.; Sánchez-Fernández, D. Thermal tolerance and vulnerability to climate change in subterranean species: A case study using an Iberian endemic pseudoscorpion. Insect Conserv. Divers. 2022, 15, 181–190. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr.; Curcic, B.P.; Cokendolpher, J.C. Arthropod phoresy involving pseudoscorpions in the past and present. Acta Arachnol. 1998, 47, 79–96. [Google Scholar] [CrossRef]
- Zeh, D.W.; Zeh, J.A. On the function of harlequin beetle-riding in the pseudoscorpion, Cordylochernes scorpioides (Pseudoscorpionida: Chernetidae). J. Arachnol. 1992, 20, 47–51. [Google Scholar]
- Zeh, J.A.; Zeh, D.W. On the threshold of dispersal: Hitchhiking on a giant fly favours exaggerated male traits in a male-dimorphic pseudoscorpion. Biol. J. Linn. Soc. 2013, 108, 509–520. [Google Scholar] [CrossRef]
- Judson, M.L.I.; Heurtault, J. Nanolpium species (Garypoidea, Olpiidae) on grasses in southern Africa-a new niche for pseudoscorpions. Rev. Suisse Zool. 1996, 1, 321–326. [Google Scholar]
- Ranius, T.; Douwes, P. Genetic structure of two pseudoscorpion species living in tree hollows in Sweden. Anim. Biodivers. Conserv. 2002, 25, 67–74. [Google Scholar]
- Harvey, M. Pseudoscorpions of the World, Version 2.0. Available online: https://www.museum.wa.gov.au/catalo-gues/pseudoscorpions (accessed on 12 March 2023).
- Beier, M. Revision der Chernetidae (Pseudoscorp.). Zool. Jahrbücher Abt. Für Syst. Okol. Und Geogr. Der Tiere 1933, 64, 509–548. [Google Scholar]
- Harvey, M.S. The phylogeny and classification of the Pseudoscorpionida (Chelicerata: Arachnida). Invertebr. Syst. 1992, 6, 1373–1435. [Google Scholar] [CrossRef]
- Beier, M. Ordnung Pseudoscorpionidea (Afterskorpione). In Bestimmungsbücher zur Bodenfauna Europas; Akademie-Verlag: Berlin, Germany, 1963; Volume 1, pp. 1–313. [Google Scholar]
- Christophoryova, J.; Šťáhlavský, F.; Fedor, P. An updated identification key to the pseudoscorpions (Arachnida: Pseudoscorpiones) of the Czech Republic and Slovakia. Zootaxa 2011, 2876, 35–48. [Google Scholar] [CrossRef]
- Harvey, M.S. Catalogue of the Pseudoscorpionida; Manchester University Press: Manchester, UK, 1990. [Google Scholar]
- Kekkonen, M.; Hebert, P.D. DNA barcode-based delineation of putative species: Efficient start for taxonomic workflows. Mol. Ecol. Resour. 2014, 14, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, T.P.; Hugg, L.; Zeh, J.A.; Zeh, D.W. Mitochondrial DNA sequencing reveals extreme genetic differentiation in a cryptic species complex of neotropical pseudoscorpions. Mol. Phylogenetics Evol. 1997, 7, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Ohira, H.; Sato, K.; Tsutsumi, T.; Kaneko, S.; Choi, H.-J. DNA barcoding suggested the existence of cryptic species and high biodiversity of South Korean pseudoscorpions (Arachnida, Pseudoscorpiones). J. Asia-Pac. Biodivers. 2018, 11, 399–407. [Google Scholar] [CrossRef]
- Ohira, H.; Kaneko, S.; Faulks, L.; Tsutsumi, T. Unexpected species diversity within Japanese Mundochthonius pseudoscorpions (Pseudoscorpiones: Chthoniidae) and the necessity for improved species diagnosis revealed by molecular and morphological examination. Invertebr. Syst. 2018, 32, 259–277. [Google Scholar] [CrossRef]
- Harrison, S.E.; Guzik, M.T.; Harvey, M.S.; Austin, A.D. Molecular phylogenetic analysis of Western Australian troglobitic chthoniid pseudoscorpions (Pseudoscorpiones: Chthoniidae) points to multiple independent subterranean clades. Invertebr. Syst. 2014, 28, 386–400. [Google Scholar] [CrossRef]
- Murienne, J.; Harvey, M.S.; Giribet, G. First molecular phylogeny of the major clades of Pseudoscorpiones (Arthropoda: Chelicerata). Mol. Phylogenetics Evol. 2008, 49, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Benavides, L.R.; Cosgrove, J.G.; Harvey, M.S.; Giribet, G. Phylogenomic interrogation resolves the backbone of the Pseudoscorpiones tree of life. Mol. Phylogenetics Evol. 2019, 139, 106509. [Google Scholar] [CrossRef]
- Pfeiler, E.; Bitler, B.G.; Castrezana, S.; Matzkin, L.M.; Markow, T.A. Genetic diversification and demographic history of the cactophilic pseudoscorpion Dinocheirus arizonensis from the Sonoran Desert. Mol. Phylogenetics Evol. 2009, 52, 133–141. [Google Scholar] [CrossRef]
- Harms, D.; Roberts, J.D.; Harvey, M.S. Climate variability impacts on diversification processes in a biodiversity hotspot: A phylogeography of ancient pseudoscorpions in south-western Australia. Zool. J. Linn. Soc. 2019, 186, 934–949. [Google Scholar] [CrossRef]
- Harms, D. The origins of diversity in ancient landscapes: Deep phylogeographic structuring in a pseudoscorpion (Pseudotyrannochthoniidae: Pseudotyrannochthonius) reflects Plio-Pleistocene climate fluctuations. Zool. Anz. 2018, 273, 112–123. [Google Scholar] [CrossRef]
- Lorenz, M.; Loria, S.F.; Harvey, M.S.; Harms, D. The Hercules pseudoscorpions from Madagascar: A systematic study of Feaellidae (Pseudoscorpiones: Feaelloidea) highlights regional endemism and diversity in one of the “hottest” biodiversity hotspots. Arthropod Syst. Phylogeny 2022, 80, 649–691. [Google Scholar] [CrossRef]
- DeSalle, R.; Goldstein, P. Review and interpretation of trends in DNA barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef]
- Yu, J.; Wu, X.; Liu, C.; Newmaster, S.; Ragupathy, S.; Kress, W.J. Progress in the use of DNA barcodes in the identification and classification of medicinal plants. Ecotoxicol. Environ. Saf. 2021, 208, 111691. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Wheeler, D.L. GenBank. Nucleic Acids Res. 2012, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed]
- Dewaard, J.R.; Levesque-Beaudin, V.; Dewaard, S.L.; Ivanova, N.V.; McKeown, J.T.; Miskie, R.; Naik, S.; Perez, K.H.; Ratnasingham, S.; Sobel, C.N.; et al. Expedited assessment of terrestrial arthropod diversity by coupling Malaise traps with DNA barcoding. Genome 2019, 62, 85–95. [Google Scholar] [CrossRef]
- Senapati, D.; Bhattacharya, M.; Kar, A.; Chini, D.S.; Das, B.K.; Patra, B.C. Environmental DNA (eDNA): A promising biological survey tool for aquatic species detection. In Proceedings of the Zoological Society; Springer: Berlin/Heidelberg, Germany, 2019; pp. 211–228. [Google Scholar]
- Schenk, J.; Kleinbölting, N.; Traunspurger, W. Comparison of morphological, DNA barcoding, and metabarcoding characterizations of freshwater nematode communities. Ecol. Evol. 2020, 10, 2885–2899. [Google Scholar] [CrossRef]
- Unger, S.D.; Williams, L.A.; Diaz, L.; Jachowski, C.B. DNA barcoding to assess diet of larval eastern hellbenders in North Carolina. Food Webs 2020, 22, e00134. [Google Scholar] [CrossRef]
- Johnson, N.S.; Lewandoski, S.A.; Merkes, C. Assessment of sea lamprey (Petromyzon marinus) diet using DNA metabarcoding of feces. Ecol. Indic. 2021, 125, 107605. [Google Scholar] [CrossRef]
- Fisher, B.L.; Smith, M.A. A revision of Malagasy species of Anochetus Mayr and Odontomachus Latreille (Hymenoptera: Formicidae). PLoS ONE 2008, 3, e1787. [Google Scholar] [CrossRef] [PubMed]
- Alcántar-Escalera, F.J.; García-Varela, M.; Vázquez-Domínguez, E.; de León, G.P.-P. Using DNA barcoding to link cystacanths and adults of the acanthocephalan Polymorphus brevis in central Mexico. Mol. Ecol. Resour. 2013, 13, 1116–1124. [Google Scholar] [PubMed]
- Grzywacz, A.; Jarmusz, M.; Walczak, K.; Skowronek, R.; Johnston, N.P.; Szpila, K. DNA barcoding identifies unknown females and larvae of Fannia R.-D. (Diptera: Fanniidae) from carrion succession experiment and case report. Insects 2021, 12, 381. [Google Scholar] [CrossRef] [PubMed]
- DeWaard, J.R.; Ratnasingham, S.; Zakharov, E.V.; Borisenko, A.V.; Steinke, D.; Telfer, A.C.; Perez, K.H.J.; Sones, J.E.; Young, M.R.; Levesque-Beaudin, V.; et al. A reference library for Canadian invertebrates with 1.5 million barcodes, voucher specimens, and DNA samples. Sci. Data 2019, 6, 308. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vaamonde, C.; Kirichenko, N.; Cama, A.; Doorenweerd, C.; Godfray, H.C.J.; Guiguet, A.; Gomboc, S.; Huemer, P.; Landry, J.-F.; Laštůvka, A.; et al. Evaluating DNA barcoding for species identification and discovery in European gracillariid moths. Front. Ecol. Evol. 2021, 9, 626752. [Google Scholar] [CrossRef]
- Hajibabaei, M.; Singer, G.A.; Hebert, P.D.; Hickey, D.A. DNA barcoding: How it complements taxonomy, molecular phylogenetics and population genetics. TRENDS Genet. 2007, 23, 167–172. [Google Scholar] [CrossRef] [PubMed]
- James, S.W.; Porco, D.; Decaens, T.; Richard, B.; Rougerie, R.; Erseus, C. DNA barcoding reveals cryptic diversity in Lumbricus terrestris L., 1758 (Clitellata): Resurrection of L. herculeus (Savigny, 1826). PLoS ONE 2010, 5, e15629. [Google Scholar] [CrossRef] [PubMed]
- Lynghammar, A.; Christiansen, J.S.; Griffiths, A.M.; Fevolden, S.-E.; Hop, H.; Bakken, T. DNA barcoding of the northern Northeast Atlantic skates (Chondrichthyes, Rajiformes), with remarks on the widely distributed starry ray. Zool. Scr. 2014, 43, 485–495. [Google Scholar] [CrossRef]
- Brasier, M.J.; Harle, J.; Wiklund, H.; Jeffreys, R.M.; Linse, K.; Ruhl, H.A.; Glover, A.G. Distributional patterns of polychaetes across the West Antarctic based on DNA barcoding and particle tracking analyses. Front. Mar. Sci. 2017, 4, 356. [Google Scholar] [CrossRef]
- Chamberlin, J.C. A synoptic revision of the generic classification of the chelonethid family Cheliferidae Simon. (Arachnida). Can. Entomol. 1932, 64, 35–39. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270 (Suppl. S1), S96–S99. [Google Scholar] [CrossRef]
- Shorthouse, D.P. SimpleMappr, an online tool to produce publication-quality point maps. 2010. Available online: https://www.simplemappr.net (accessed on 18 June 2023).
- Ersts, P.J. Geographic Distance Matrix Generator Version 1.23; American Museum of Natural History: New York, NY, USA, 2012; Available online: https://biodiversityinformatics.amnh.org/open_source/gdmg (accessed on 11 July 2023).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit 1 from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Drummond. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 1–8. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- FigTree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 4 September 2023).
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Ezard, T.; Fujisawa, T.; Barraclough, T.G. Splits: Species’ limits by threshold statistics. R Package Version 2009, 1, r29. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Santibáñez-López, C.E.; González-Santillán, E.; Monod, L.; Sharma, P.P. Phylogenomics facilitates stable scorpion systematics: Reassessing the relationships of Vaejovidae and a new higher-level classification of Scorpiones (Arachnida). Mol. Phylogenetics Evol. 2019, 135, 22–30. [Google Scholar] [CrossRef]
- Benavides, L.R.; Pinto-da-Rocha, R.; Giribet, G. The phylogeny and evolution of the flashiest of the armored harvestmen (Arachnida: Opiliones). Syst. Biol. 2021, 70, 648–659. [Google Scholar] [CrossRef]
- Dupuis, J.R.; Roe, A.D.; Sperling, F.A. Multi-locus species delimitation in closely related animals and fungi: One marker is not enough. Mol. Ecol. 2012, 21, 4422–4436. [Google Scholar] [CrossRef]
- Edwards, D.L.; Knowles, L.L. Species detection and individual assignment in species delimitation: Can integrative data increase efficacy? Proc. R. Soc. B Biol. Sci. 2014, 281, 20132765. [Google Scholar] [CrossRef] [PubMed]
- Carstens, B.C.; Pelletier, T.A.; Reid, N.M.; Satler, J.D. Satler. How to fail at species delimitation. Mol. Ecol. 2013, 22, 4369–4383. [Google Scholar] [CrossRef] [PubMed]
- Hlebec, D.; Harms, D.; Kučinić, M.; Harvey, M.S. Integrative taxonomy of the pseudoscorpion family Chernetidae (Pseudoscorpiones: Cheliferoidea): Evidence for new range-restricted species in the Dinaric Karst. Zool. J. Linn. Soc. 2023. [Google Scholar] [CrossRef]
- Arabi, J.; Judson, M.L.I.; Deharveng, L.; Lourenço, W.R.; Cruaud, C.; Hassanin, A. Nucleotide composition of CO1 sequences in Chelicerata (Arthropoda): Detecting new mitogenomic rearrangements. J. Mol. Evol. 2012, 74, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Astrin, J.J.; Höfer, H.; Spelda, J.; Holstein, J.; Bayer, S.; Hendrich, L.; Huber, B.A.; Kielhorn, K.-H.; Krammer, H.-J.; Lemke, M.; et al. Towards a DNA barcode reference database for spiders and harvestmen of Germany. PLoS ONE 2016, 11, e0162624. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Huang, J.-X.; Liu, H.-H.; Chang, C.-H. Two New Pseudoscorpion Species of the Coastal Genus Garypus L. Koch, 1873 (Garypidae) and an Updated Checklist of the Pseudoscorpiones of Taiwan. Zool. Stud. 2022, 61, e24. [Google Scholar] [PubMed]
- Levi, H.W. Notes on the life history of the pseudoscorpion Chelifer cancroides (Linn.) (Chelonethida). Trans. Am. Microsc. Soc. 1948, 67, 290–298. [Google Scholar] [CrossRef]
- Stemme, T.; Pfeffer, S.E. Anatomy of the nervous system in Chelifer cancroides (Arachnida: Pseudoscorpiones) with a distinct sensory pathway associated with the pedipalps. Insects 2021, 13, 25. [Google Scholar] [CrossRef]
- Harvey, M.S. A review and redescription of the cosmopolitan pseudoscorpion Chelifer cancroides (Pseudoscorpiones: Cheliferidae). J. Arachnol. 2014, 42, 86–104. [Google Scholar] [CrossRef]
- Krämer, J.; Lüddecke, T.; Marner, M.; Maiworm, E.; Eichberg, J.; Hardes, K.; Schäberl, T.F.; Vilcinskas, A.; Predel, R. Antimicrobial, insecticidal and cytotoxic activity of linear venom peptides from the pseudoscorpion Chelifer cancroides. Toxins 2022, 14, 58. [Google Scholar] [CrossRef]
- Červená, M.; Krajčovičová, K.; Christophoryová, J. Recent data about diversity and distribution of pseudoscorpions (Arachnida: Pseudoscorpiones) collected from different habitat types in Slovakia. Klapalekiana 2020, 56, 1–18. [Google Scholar]
- Rowe, D.L.; Dunn, K.A.; Adkins, R.M.; Honeycutt, R.L. Molecular clocks keep dispersal hypotheses afloat: Evidence for trans-Atlantic rafting by rodents. J. Biogeogr. 2010, 37, 305–324. [Google Scholar] [CrossRef]
- Beier, M. Pseudoscorpionidea II. Subord. C. Cheliferinea; Tierreich, 58, i–xxi+ 1–294; De Gruyter: Berlin, Germany; Boston, MA, USA, 1932. [Google Scholar]
- Novák, J.; Danyi, L. Catalogue of the type material of pseudoscorpions (Arachnida: Pseudoscorpiones) deposited in the Hungarian Natural History Museum, Budapest. Zootaxa 2018, 4527, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Menge, A. Uber die Scheerenspinnen, Chernetidae. Schr. Naturf. Ges. Danz. 1855, 5, 1–41. [Google Scholar]
- Caputi, L.; Andreakis, N.; Mastrototaro, F.; Cirino, P.; Vassillo, M.; Sordino, P. Cryptic speciation in a model invertebrate chordate. Proc. Natl. Acad. Sci. USA 2007, 104, 9364–9369. [Google Scholar] [CrossRef] [PubMed]
- Wattier, R.; Mamos, T.; Copilaş-Ciocianu, D.; Jelić, M.; Ollivier, A.; Chaumot, A.; Danger, M.; Felten, V.; Piscart, C.; Žganec, K.; et al. Continental-scale patterns of hyper-cryptic diversity within the freshwater model taxon Gammarus fossarum (Crustacea, Amphipoda). Sci. Rep. 2020, 10, 16536. [Google Scholar] [CrossRef] [PubMed]
- Erséus, C.; Gustafsson, D. Cryptic speciation in clitellate model organisms. In Annelids in Modern Biology; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 31–46. [Google Scholar]
- Heneberg, P.; Bogusch, P.; Astapenková, A. Reed galls serve as an underestimated but critically important resource for an assemblage of aculeate hymenopterans. Biol. Conserv. 2014, 172, 146–154. [Google Scholar] [CrossRef]
- Bogusch, P.; Macek, J.; Janšta, P.; Kubík, Š.; Řezáč, M.; Holý, K.; Malenovský, I.; Baňař, P.; Mikát, M.; Astapenková, A.; et al. Industrial and post-industrial habitats serve as critical refugia for pioneer species of newly identified arthropod assemblages associated with reed galls. Biodivers. Conserv. 2016, 25, 827–863. [Google Scholar] [CrossRef]
- Bogusch, P.; Heneberg, P.; Astapenková, A. Habitat requirements of wetland bees and wasps: Several reed-associated species still rely on peaty meadows and other increasingly rare wetland habitats. Wetl. Ecol. Manag. 2020, 28, 921–936. [Google Scholar] [CrossRef]
- Schwentner, M.; Giribet, G. Phylogeography, species delimitation and population structure of a Western Australian short-range endemic mite harvestman (Arachnida: Opiliones: Pettalidae: Karripurcellia). Evol. Syst. 2018, 2, 81–87. [Google Scholar] [CrossRef]
- Štundlová, J.; Šmíd, J.; Nguyen, P.; Šťáhlavský, F. Cryptic diversity and dynamic chromosome evolution in Alpine scorpions (Euscorpiidae: Euscorpius). Mol. Phylogenetics Evol. 2019, 134, 152–163. [Google Scholar] [CrossRef]
- Moulds, T.A.; Murphy, N.; Adams, M.; Reardon, T.; Harvey, M.S.; Jennings, J.; Austin, A.D. Phylogeography of cave pseudoscorpions in southern Australia. J. Biogeogr. 2007, 34, 951–962. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Just, P.; Šťáhlavský, F.; Bogusch, P.; Astapenková, A.; Opatova, V. Dispersal Capabilities Do Not Parallel Ecology and Cryptic Speciation in European Cheliferidae Pseudoscorpions (Pseudoscorpiones: Cheliferidae). Diversity 2023, 15, 1040. https://doi.org/10.3390/d15101040
Just P, Šťáhlavský F, Bogusch P, Astapenková A, Opatova V. Dispersal Capabilities Do Not Parallel Ecology and Cryptic Speciation in European Cheliferidae Pseudoscorpions (Pseudoscorpiones: Cheliferidae). Diversity. 2023; 15(10):1040. https://doi.org/10.3390/d15101040
Chicago/Turabian StyleJust, Pavel, František Šťáhlavský, Petr Bogusch, Alena Astapenková, and Vera Opatova. 2023. "Dispersal Capabilities Do Not Parallel Ecology and Cryptic Speciation in European Cheliferidae Pseudoscorpions (Pseudoscorpiones: Cheliferidae)" Diversity 15, no. 10: 1040. https://doi.org/10.3390/d15101040