

Cestode Diversity of Shrews on the Kamchatka Peninsula and Paramushir Island

Abstract
:1. Introduction
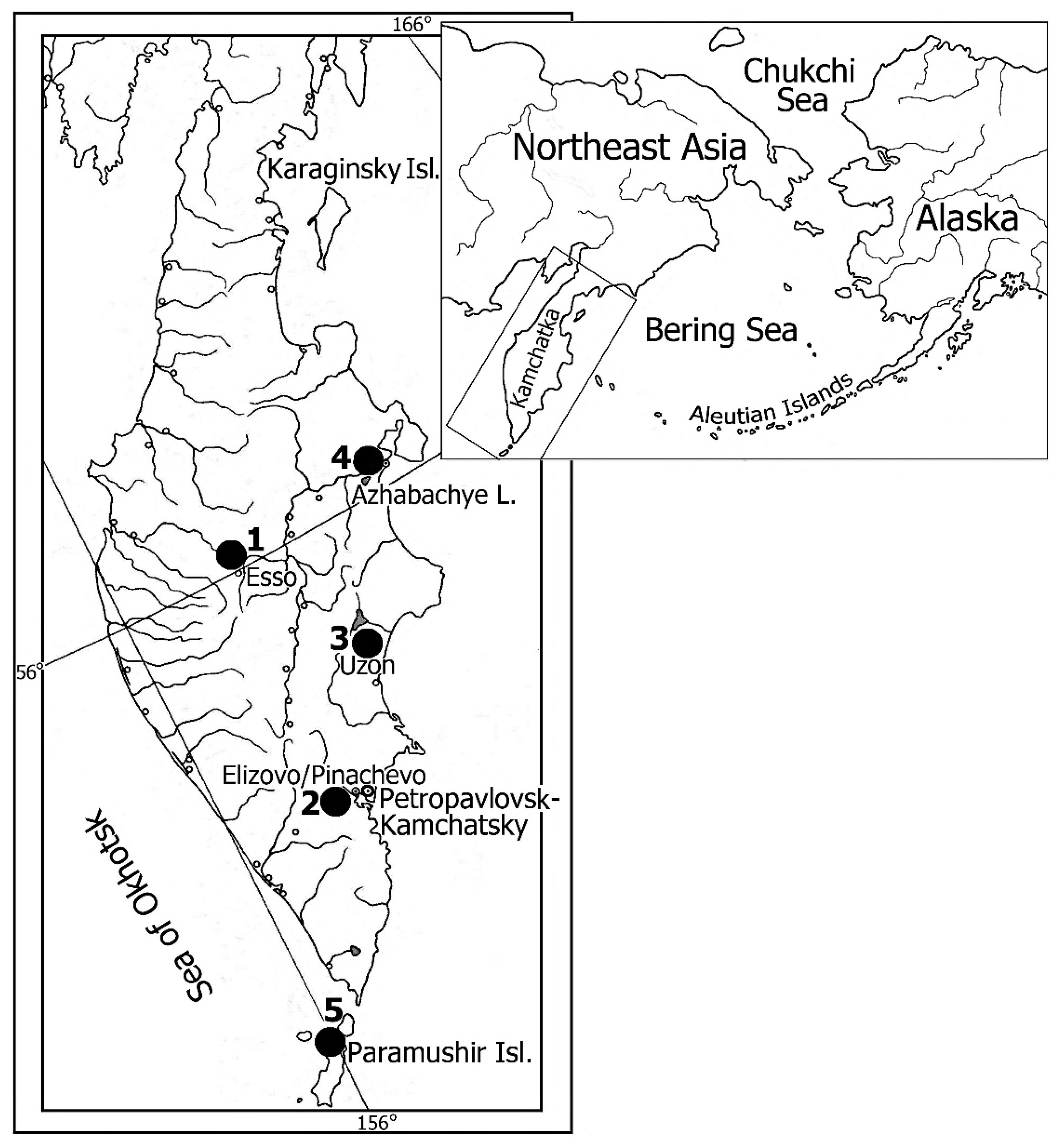
2. Materials and Methods
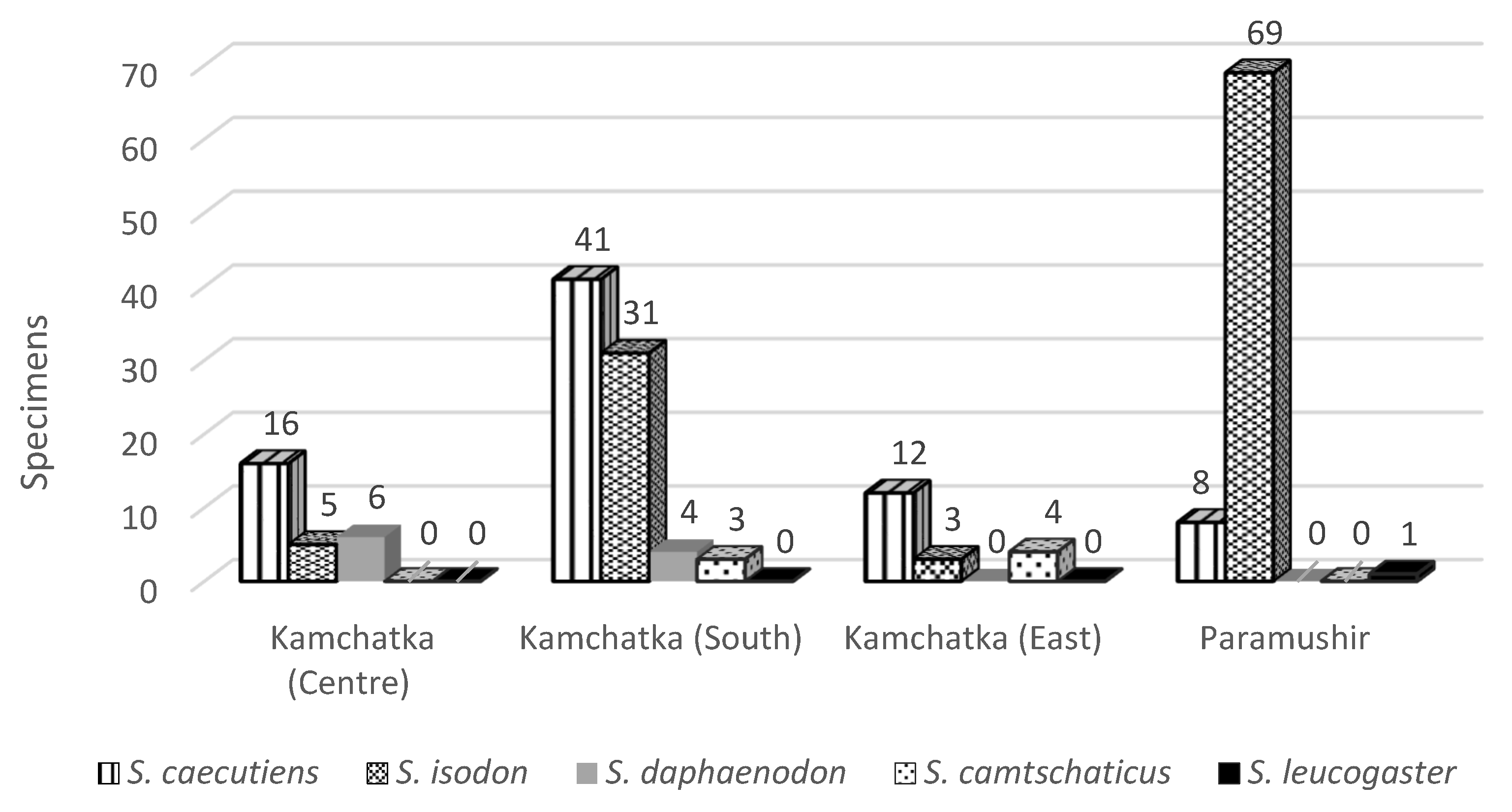
3. Results
3.1. Species Diversity of Shrews Cestodes of Kamchatka Peninsula and Paramushir Island
3.2. Prevalence of Infection of Shrew Cestodes of Kamchatka Peninsula and Paramushir Island
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Mammal Collection of the Institute of Biological Problems of the North FEB RAS (IBPN, Magadan, Russia) with field number labels NED No/Year (N.E. Dokuchaev) and VDG No/Year (V.D. Gulyaev), Mammal Collection of the Institute of Systematics and Ecology of Animals (ISEA SB RAS, Novosibirsk, Russia) with field number labels SVK No/Year (S.V. Karpenko).
- RUSSIA, Kamchatka Peninsula, the vicinities of the settlement Esso:
- Sorex caecutiens (NED42/2002, 52/2002, 53/2002, 54/2002, 57/2002, 60/2002, 65/2002, 66/2002, 71/2002, 76/2002, 77/2002, 79/2002, 81/2002, 87/2002, 88/2002, 93/2002),
- Sorex daphaenodon (NED49/2002, 51/2002, 61/2002, 62/2002, 63/2002, 64/2002).
- Sorex isodon (NED50/2002, 58/2002, 78/2002, 80/2002, 82/2002).
- RUSSIA, Kamchatka Peninsula, the vicinities of the settlement Yelizovo:
- Sorex caecutiens (VDG7/2002, 8/2002, 10/2002, 12/2002, 13/2002, 16/2002, 17/2002, 18/2002, 19/2002, 20/2002, 23/2002, 24/2002, 25/2002).
- Sorex daphaenodon (VDG2/2002).
- Sorex isodon (VDG1/2002, 3/2002, 4/2002, 5/2002, 6/2002, 9/2002, 11/2002, 14/2002, 15/2002, 21/2002, 22/2002).
- RUSSIA, Kamchatka Peninsula, the vicinities of the settlement Pinachevo:
- Sorex caecutiens (NED180/2007, 183/2007, 184/2007, 199/2007, 202/2007, 203/2007, 209/2007, 219/2007, 221/2007, 222/2007, 223/2007, 224/2007, 232/2007, 234/2007, 235/2007, 236/2007, 237/2007, 242/2007, 247/2007, 248/2007, 249/2007, 261/2007, 263/2007).
- Sorex camtschaticus (NED182/2007, 233/2007, 264/2007).
- Sorex daphaenodon (NED181/2007, 201/2007, 220/2007, 260/2007).
- Sorex isodon (NED185/2007, 186/2007, 197/2007, 198/2007, 205/2007, 206/2007, 226/2007, 227/2007, 238/2007, 239/2007, 240/2007, 252/2007, 253/2007, 254/2007, 255/2007).
- RUSSIA, Kamchatka Peninsula, caldera Uzon:
- Sorex caecutiens (NED27/2009, 28/2009, 29/2009, 30/2009, 33/2009, 34/2009, 35/2009, 37/2009, 38/2009, 39/2009, 40/2009, 42/2009).
- Sorex camtschaticus (NED24/2009, 25/2009, 26/2009, 41/2009).
- Sorex isodon (NED31/2009, 32/2009, 36/2009).
- RUSSIA, Kamchatka Peninsula, Azhabachye Lake:
- Sorex caecutiens (SVK 1/1982, 2/1982, 4/1982, 7/1982, 8/1982, 10/1982, 14/1982, 17/1982, 33/1982, 36/1982, 59/1982, 64/1982, 81/1982, 118/1982, 120/1982, 125/1982, 177/1982, 185/1982, 187/1982, 208/1982, 212/1982, 214/1982, 216/1982, 230/1982, 240/1982, 250/1982, 259/1982, 270/1982, 272/1982, 278/1982, 281/1982, 288/1982, 304/1982, 356/1982, 405/1982, 408/1982, 415/1982, 418/1982, 430/1982, 454/1982).
- Sorex isodon (SVK 3/1982, 5/1982, 6/1982, 15/1982, 22/1982, 56/1982, 97/1982, 101/1982, 158/1982, 211/1982, 232/1982, 237/1982, 248/1982, 255/1982, 284/1982, 305/1982, 341/1982, 342/1982, 345/1982, 347/1982, 397/1982, 400/1982, 445/1982)
- Sorex daphaenodon (SVK 252/1982, 302/1982, 393/1982, 395/1982).
- Sorex camtschaticus (SVK 25/1982, 27/1982, 425/1982, 450/1982, 451/1982).
- Sorex sp. (SVK 11/1982, 12/1982, 13/1982, 24/1982, 18/1982, 19/1982, 20/1982, 21/1982, 23/1982, 24/1982, 65/1982, 69/1982, 70/1982, 75/1982, 105/1982, 119/1982, 126/1982, 127/1982, 135/1982, 141/1982, 155/1982, 165/1982, 171/1982, 179/1982, 234/1982, 235/1982, 247/1982, 258/1982, 289/1982, 290/1982, 325/1982, 350/1982, 351/1982, 353/1982, 370/1982, 374/1982, 394/1982, 412/1982, 413/1982, 432/1982, 435/1982, 436/1982).
- RUSSIA, Paramushir Island:
- Sorex caecutiens (NED 132/2007, 138/2007, 139/2007, 140/2007, 141/2007, 149/2007, 150/2007, 152/2007).
- Sorex isodon (NED7/2007, 8/2007, 9/2007, 10/2007, 11/2007, 12/2007, 13/2007, 14/2007, 15/2007, 17/2007, 18/2007, 19/2007, 31/2007, 32/2007, 33/2007, 34/2007, 35/2007, 36/2007, 37/2007, 38/2007, 40/2007, 41/2007, 53/2007, 54/2007, 55/2007, 56/2007, 57/2007, 58/2007, 59/2007, 60/2007, 67/2007, 68/2007, 69/2007, 72/2007, 73/2007, 74/2007, 75/2007, 76/2007, 77/2007, 78/2007, 85/2007, 86/2007, 87/2007, 102/2007, 103/2007, 104/2007, 105/2007, 106/2007, 107/2007, 108/2007, 109/2007, 110/2007, 111/2007, 112/2007, 126/2007, 127/2007, 128/2007, 129/2007, 133/2007, 134/2007, 135/2007, 136/2007, 137/2007, 151/2007, 153/2007, 154/2007).
- Sorex leucogaster (NED148/2007).
Appendix B
- The helminthological collection in the Institute of Systematics and Ecology of Animals (ISEA SB RAS, Novosibirsk, Russia), NED Number of slide/field number labels/Year (N.E. Dokuchaev), VDG Number of slide/field number labels/Year (V.D. Gulyaev), SVK Number of slide/field number labels/Year (S.V. Karpenko)
- RUSSIA, Kamchatka Peninsula, the vicinities of the settlement Esso:
- Sorex caecutiens (NED 2/42/2002, 1–3/53/2002, 1/54/2002, 1/57/2002, 1/66/2002, 1/71/2002, 1/76/2002, 1/81/2002, 1–2/88/2002, 1/93/2002),
- Sorex daphaenodon (NED 1/51/2002, 1/61/2002, 1/62/2002, 1/63/2002, 1/64/2002).
- Sorex isodon (NED 1/58/2002, 2/80/2002, 1/82/2002).
- RUSSIA, Kamchatka Peninsula, the vicinities of the settlement Yelizovo:
- Sorex caecutiens (VDG 1/8/2002, 1–2/10/2002, 1/13/2002, 1/16/2002, 1/17/2002, 1/19/2002, 1/20/2002, 1/23/2002, 1/24/2002, 1–2/25/2002).
- Sorex daphaenodon (VDG 1/2/2002).
- Sorex isodon (VDG 1/1/2002, 1–2/3/2002, 1/6/2002, 1–49/2002, 1/11/2002, 1/14/2002, 1/15/2002, 1/22/2002).
- RUSSIA, Kamchatka Peninsula, the vicinities of the settlement Pinachevo:
- Sorex caecutiens (NED 1/180/2007, 1/183/2007, 1/184/2007, 1/199/2007, 1/202/2007, 1/203/2007, 1/219/2007, 1/221/2007, 1/222/2007, 1/223/2007, 1/224/2007, 1/232/2007, 1/234/2007, 1–7/235/2007, 1/236/2007, 1/237/2007, 1/242/2007, 1/247/2007, 1/249/2007, 1/261/2007, 1/263/2007).
- Sorex camtschaticus (NED 1–5/264/2007).
- Sorex daphaenodon (NED 1/181/2007, 1/201/2007, 1/260/2007).
- Sorex isodon (NED 1–8/185/2007, 1–2/186/2007, 1–2/198/2007, 1/206/2007, 1–3/226/2007, 1/227/2007, 1–4238/2007, 1/240/2007, 1/253/2007, 1/254/2007, 1/255/2007).
- RUSSIA, Kamchatka Peninsula, caldera Uzon:
- Sorex caecutiens (NED1/27/2009, 1–3/39/2009, 1/42/2009)
- Sorex camtschaticus (NED1/24/2009).
- Sorex isodon (NED1/31/2009, 1/36/2009).
- RUSSIA, Kamchatka Peninsula, Azhabachye Lake:
- Sorex caecutiens (SVK 7/1982, 10/1982, 17/1982, 33/1982, 36/1982, 59/1982, 81/1982, 118/1982, 120/1982, 125/1982, 208/1982, 212/1982, 214/1982, 216/1982, 230/1982, 240/1982, 259/1982, 270/1982, 272/1982, 278/1982, 281/1982, 288/1982, 304/1982, 356/1982, 405/1982, 408/1982, 430/1982, 454/1982).
- Sorex isodon (SVK 22/1982, 56/1982, 97/1982, 101/1982, 158/1982, 211/1982, 232/1982, 237/1982, 248/1982, 305/1982, 341/1982, 342/1982, 397/1982)
- Sorex camtschaticus (SVK 25/1982, 27/1982)
- Sorex daphaenodon (SVK 302/1982, 395/1982)
- Sorex sp. (SVK 11/1982, 12/1982, 24/1982, 70/1982, 75/1982, 105/1982, 119/1982, 141/1982, 234/1982, 235/1982, 247/1982, 258/1982, 289/1982, 290/1982, 325/1982, 350/1982, 351/1982, 353/1982, 370/1982, 394/1982, 412/1982, 413/1982, 432/1982, 435/1982, 436/1982)
- RUSSIA, Paramushir Island:
- Sorex caecutiens (NED 1/132/2007, 1–4/138/2007, 1/139/2007, 1/140/2007, 1/141/2007, 1/149/2007, 1–3/150/2007, 1–2/152/2007).
- Sorex isodon (NED1–6/7/2007, 1–3/8/2007, 1–3/9/2007, 1–2/10/2007, 1–3/11/2007, 1/12/2007, 1–4/13/2007, 1–3/14/2007, 1–3/15/2007, 1/17/2007, 1/18/2007, 1/19/2007, 1/31/2007, 1/32/2007, 1/33/2007, 1/34/2007, 1/35/2007, 1/36/2007, 1/37/2007, 1/38/2007, 1/40/2007, 1/41/2007, 1/53/2007, 1/54/2007, 1/55/2007, 1/56/2007, 1–14/57/2007, 1/58/2007, 2/59/2007, 1/60/2007, 1/67/2007, 1/68/2007, 1/69/2007, 1/72/2007, 1/73/2007, 1–2/74/2007, 1–4/75/2007, 1/76/2007, 1/77/2007, 1/78/2007, 1/85/2007, 1/86/2007, 1/87/2007, 1/102/2007, 1–2/103/2007, 1/104/2007, 1/105/2007, 1/106/2007, 1/107/2007, 1/108/2007, 1–3/109/2007, 1/110/2007, 1/111/2007, 1/112/2007, 1–2/126/2007, 1/127/2007, 1–2/128/2007, 1/129/2007, 1/133/2007, 1/134/2007, 1/135/2007, 1–4/136/2007, 1/137/2007, 1/151/2007, 1/153/2007, 1–3/154/2007).
- Sorex leucogaster (NED 1/148/2007).
References
- Kurentsov, A.I. Zoogeography of Kamchatka. In Kamchatka Fauna; Kamchatka Complex Expedition of the USSR Academy of Sciences. Moscow–Leningrad; Publishing House of AS USSR: Saint Petersburg, Russia, 1963; pp. 3–60. (In Russian) [Google Scholar]
- Dokuchaev, N.E. Shrews of Northeast Asia and Neighboring Territories. Ph.D. Thesis, Biology and Soil Institute of the FESC, Vladivostok, Russia, 1998; 64p. (In Russian). [Google Scholar]
- Pietsch, T.W.; Bogatov, V.V.; Amaoka, K.; Zhuravlev, Y.N.; Barkalov, V.Y.; Gage, S.; Takahashi, H.; Lelej, A.S.; Storozhenko, S.Y.; Minakawa, N.; et al. Biodiversity and biogeography of the islands of the Kuril Archipelago. J. Biogeogr. 2003, 30, 1297–1310. [Google Scholar] [CrossRef]
- Kostenko, V.A.; Nesterenko, V.A.; Trukhin, A.M. Mammals of the Kuril Archipelago; Dalnauka: Vladivostok, Russia, 2004; 186p. (In Russian) [Google Scholar]
- Karpenko, S.V. Helminths of the shrews of the Kamchatka Peninsula. In Proceedings of the II Scientific Conference of Novosibirsk Branch of Parasitological Society RAS, Novosibirsk, Russia, 15 April 1997; pp. 50–52. (In Russian). [Google Scholar]
- Dokuchaev, N.E.; Melnikova, Y.A.; Gulyaev, V.D. Cestodes of shrews in Kamchatka and adjacent areas. In Proceedings of the IV Scientific Conference, Petropavlovsk-Kamchatsky, Russia, 18–19 November 2003; pp. 39–42. (In Russian). [Google Scholar]
- Melnikova, Y.A.; Gulyaev, V.D.; Dokuchaev, N.E. The structure of community of cestodes of shrews of West Beringia and the Far East. In Proceedings of the International Conference and 3rd Congress of the Russian Society of Parasitologists—Russian Academy of Sciences, Petrozavodsk, Russia, 6–12 October 2003; pp. 18–20. (In Russian). [Google Scholar]
- Gulyaev, V.D.; Kornienko, S.A.; Dokuchaev, N.E. Cestodes of the shrews of the Paramushir island. In Proceedings of the International Symposium Dedicated to the 130-th Anniversary of Acad. K.I. Skrjabin, Moscow, Russia, 9–11 December 2008; pp. 91–93. (In Russian). [Google Scholar]
- Gulyaev, V.D.; Lykova, K.A.; Melnikova, Y.A.; Bibik, E.V. Validity of species Spasskylepis ovaluteri (Cestoda, Cyclophyllidae, Hymenolepididae)—Parasite of shrews from Palearctic. Zoolog. Zh. 2004, 83, 387–394. (In Russian) [Google Scholar]
- Gulyaev, V.D.; Kornienko, S.A.; Dokuchaev, N.E. The cestodes of Staphylocystoides Yamaguti, 1959 genus (Cestoda, Hymenolepididae) in shrews of Beringia. Bull. North-East Sci. Cent. FE Branch RAS 2007, 4, 75–84. (In Russian) [Google Scholar]
- Binkienė, R.; Kontrimavichus, V.; Hoberg, E.P. Overview of the cestode fauna of European shrews of the genus Sorex with comments on the fauna in Neomys and Crocidura and an exploration of historical processes in post-glacial Europe. J. Helminthol. 2011, 48, 207–228. [Google Scholar] [CrossRef] [Green Version]
- Binkienė, R.; Kornienko, S.A.; Tkach, V.V. Soricinia genovi n. sp. from Neomys fodiens in Bulgaria, with redescription of Soricinia globosa (Baer, 1931) (Cyclophyllidea: Hymenolepididae). Parasitol. Res. 2015, 114, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Kornienko, S.A. Species diversity of cestodes of genus Neoskrjabinolepis (Cestoda, Hymenolepididae)—Parasites of shrews (Soricidae) of Palaearctic. In The Problems of Cestodology; Pugachev, O.N., Ed.; Elmor: St. Petersburg, Russia, 2012; Volume 4, pp. 85–101. (In Russian) [Google Scholar]
- Kornienko, S.A.; Gulyaev, V.D. Species resources of cestodes of genus Neoskrjabinolepis of Palaearctic shrews. In Proceedings of the IV Congress of the Russian Society of Parasitologists—RAS, St. Petersburg, Russia, 20–25 October 2008; pp. 72–75. (In Russian). [Google Scholar]
- Kornienko, S.A.; Gulyaev, V.D. New cestodes of the genus Neoskrjabinolepis (Cestoda, Hymenolepididae) from shrews of the Far East. Zoolog. Zh. 2011, 90, 1296–1303. (In Russian) [Google Scholar]
- Kornienko, S.A.; Gulyaev, V.D.; Melnikova, Y.A. On the morphology and systematics of cestodes of the genus Neoskrjabinolepis Spassky, 1947 (Cyclophyllidea, Hymenolepididae). Zoolog. Zh. 2006, 85, 134–145. (In Russian) [Google Scholar]
- Kornienko, S.A.; Gulyaev, V.D.; Melnikova, Y.A. New species of cestodes of the genus Neoskrjabinolepis Spassky, 1947 (Cyclophyllidea, Hymenolepididae) from shrews of Russia. Zoolog. Zh. 2007, 86, 259–269. (In Russian) [Google Scholar]
- Kornienko, S.A.; Gulyaev, V.D.; Melnikova, Y.A.; Georgiev, B.B. Neoskrjabinolepis nuda n. sp. from shrews in Sakhalin Island, Russia, with a taxonomic review of the genus Neoskrjabinolepis Spasskii, 1947 (Cyclophyllidea, Hymenolepidide). Syst. Parasytol. 2008, 70, 147–158. [Google Scholar] [CrossRef]
- Kornienko, S.; Binkienė, R.; Tkach, V.V. Revision of the genus Soricinia Spassky & Spasskaja, 1954 (Cestoda: Cyclophyllidea: Hymenolepididae) with redescriptions of three species, an amended generic diagnosis and an identification key to species. Syst. Parasitol. 2016, 93, 451–465. [Google Scholar]
- Kornienko, S.A.; Binkienė, R.; Dokuchaev, N.E.; Tkach, V.V. Molecular phylogeny and systematics of cestodes with rudimentary rostellum (Cestoda: Hymenolepididae) from Holarctic Sorex shews (Eulipotyphla: Soricidae). Zool. J. Lin. Soc. 2019, 187, 965–986. [Google Scholar] [CrossRef]
- Mariaux, J.; Tkach, V.V.; Vasileva, G.P.; Waeschenbach, A.; Beveridge, I.; Dimitrova, Y.D.; Haukisalmi, V.; Greiman, S.E.; Littlewood, D.T.J.; Makarikov, A.A.; et al. Cyclophyllidea van Beneden in Braun, 1900. In Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth; Caira, J.N., Jensen, K., Eds.; University of Kansas, Natural History Museum: Lawrence, KS, USA, 2017; pp. 77–148. [Google Scholar]
- Neov, B.; Vasileva, G.P.; Radoslavov, G.; Hristov, P.; Littlewood, D.T.J.; Georgiev, B.B. Phylogeny of hymenolepidid cestodes (Cestoda: Cyclophyllidea) from mammalian hosts based on partial 28S rDNA, with focus on parasites from shrews. Parasitol. Res. 2019, 118, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Neov, B.; Vasileva, G.P.; Radoslavov, G.; Hristov, P.; Littlewood, D.T.J.; Georgiev, B.B. Phylogeny of hymenolepidids (Cestoda: Cyclophyllidea) from mammals: Sequences of 18S rDNA and COI genes confirm major clades revealed by the 28S rDNA analyses. J. Helminthol. 2021, 95, E23. [Google Scholar] [CrossRef] [PubMed]
- Ivashkin, V.M.; Kontrimavichus, V.L.; Nazarova, N.S. Methods of Collection and Study of Helminths from Terrestrial Mammals; Nauka: Moscow, Russia, 1971. (In Russian) [Google Scholar]
- Fedorov, K.P. Patterns of Spatial Distribution of Parasitic Worms; Nauka: Novosibirsk, Russia, 1986; 256p. (In Russian) [Google Scholar]
- Gulyaev, V.D. Species diversity and taxonomic structure of hymenolepidid cestodes of shrews in Holarctic. In Proceedings of the International Conference and 3rd Congress of the Russian Society of Parasitologists—Russian Academy of Sciences, Petrozavodsk, Russia, 6–12 October 2003; pp. 137–139. (In Russian). [Google Scholar]
- Kornienko, S.A. Tapeworms of Northern Asian shrews. In Proceedings of the IV All-Russian Conference with International Participation, Ulan-Ude, Russia, 15–18 June 2021; pp. 234–235. (In Russian). [Google Scholar]
- Kornienko, S.A.; Dokuchaev, N.E. Cestodes of shrews of the Indigirka River basin. Bull. North-East Sci. Cent. FE Branch RAS 2015, 1, 42–48. (In Russian) [Google Scholar]
- Kornienko, S.A.; Dokuchaev, N.E.; Odnokurtsev, V.A. Shrew’ cestodes of the Central and Southern Yakutia. Zoolog. Zh. 2018, 97, 1110–1120. (In Russian) [Google Scholar]
- Kornienko, S.A.; Vasileva, G.P.; Georgiev, B.B. New cestode species of the genus Neoskrjabinolepis Spassky, 1947 (Cyclophyllidea: Hymenolepididae) from the Common Shrew (Sorex araneus L.) in Europe. Syst. Parasitol. 2022, 99, 13–21. [Google Scholar] [CrossRef]
- Kornienko, S.A.; Dokuchaev, N.E. Two new cestode species of Neoskrjabinolepis Spasskii, 1947 (Cyclophyllidea: Hymenolepididae) from the tundra shrew Sorex tundrensis Merriam (Mammalia: Soricidae) in Alaska and Chukotka. Syst. Parasitol. 2012, 83, 179–188. [Google Scholar] [CrossRef]
- Dokuchaev, N.E.; Gulyaev, V.D. Quaternary history of Sorex shrews in North-Eastern Asia according to helminthological data. In Proceedings of the III All-Russian Scientific Conference on the Biology of Insectivorous Mammals, Novosibirsk, Russia, 15–20 September 2007; pp. 38–40. (In Russian). [Google Scholar]
- Greiman, S.E.; Tkach, V.V.; Cook, J.A. Description and molecular differentiation of a new Staphylocystoides (Cyclophyllidea: Hymenolepididae) from the dusky shrew Sorex monticolus in Southeast Alaska. J. Parasitol. 2013, 99, 1045–1049. [Google Scholar] [CrossRef]
- Tkach, V.V.; Kinsella, J.M.; Greiman, S.E. Two new species of Staphylocystoides yamaguti, 1959 (Cyclophyllidea: Hymenolepididae) from the masked shrew Sorex cinereus in North America. J. Parasitol. 2018, 104, 157–167. [Google Scholar] [CrossRef]
- Kornienko, S.A. Cestodes of the shrews of the Palearctic. In Proceedings of the XV Conference of Ukrainian Scientific Society of Parasitologists, Chernivtsi, Ukraine, 15–18 October 2013; p. 57. [Google Scholar]
- Kornienko, S.A.; Ishigenova, L.A. Distribution of the cestode Urocystis prolifer Villot, 1880 (Cyclophyllidea: Hymenolepididae) in the Palaearctic and new data on its postembryonic development. J. Helminthol. 2021, 95, e35, 1–11. [Google Scholar] [CrossRef]
- Melnikova, Y.A.; Gulyaev, V.D.; Dokuchaev, N.E. Cestoda of insectivorous mammals of Russian Far East. In Proceedings of the Lazovsky State Nature Reserve Named after L.G. Kaplanov, Vladivostok, Russia, 19–20 April 2005; Volume 3, pp. 82–90. (In Russian). [Google Scholar]
- Gulyaev, V.D.; Kornienko, S.A. On morphological peculiarities of cysticercoids Monocercus (Cestoda: Cyclophyllidea: Dilepididae). Parasitologiia 1998, 32, 141–145. (In Russian) [Google Scholar]
- Goüy de Bellocq, J.; Morand, S.; Feliu, C. Patterns of parasites species richness of Western Palaearctic region micro-mammals: Island effects. Ecography 2002, 25, 173–183. [Google Scholar] [CrossRef]
- Anikanova, V.S.; Boyko, N.S.; Ieshko, E.P. Helminths of shrews of the Kandalaksha Bay Islands. Teriol. Res. 2004, 5, 49–63. (In Russian) [Google Scholar]
- Anikanova, V.S.; Boyko, N.S.; Ieshko, E.P. Helminths fauna of Kandalaksha Reserve. Parasitologiia 2005, 39, 559–568. (In Russian) [Google Scholar]
- Magnanou, E.; Morand, S. Insularity and micromammal-macroparasite relationships. In Micromammals and Macroparasites; From Evolutionary Ecology to Management; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer: Tokyo, Japan, 2006; pp. 295–315. [Google Scholar]
- Kornienko, S.A.; Makarikov, A.A.; Dokuchaev, N.E. Cestodes of small mammals on the Bolshoi Shantar Island. Bull. North-East Sci. Cent. FE Branch RAS 2014, 2, 64–69. (In Russian) [Google Scholar]
- Bugmyrin, S.V.; Korosov, A.V.; Bespyatova, L.A.; Ieshko, E.P. Helminth fauna of the bank vole Myodes glareolus (Schreber, 1780) in the Kizhi archipelago. Parasitologiia 2015, 49, 61–71. (In Russian) [Google Scholar]
- Hope, A.G.; Greiman, S.E.; Tkach, V.V.; Hoberg, E.P.; Cook, J.A. Shrews and Their Parasites: Small Species Indicate Big Changes; NOAA: Washington, DC, USA, 2016; pp. 1–11. [Google Scholar]
- Combe, F.J.; Sikes, D.S.; Tkach, V.V.; Hope, A.G. Origins and diversity of the Bering Sea Island fauna: Shifting linkages across the northern continents. Biodiv. Conserv. 2021, 30, 1205–1232. [Google Scholar] [CrossRef]
- Binkienė, R. Helminth fauna of shrews (Sorex spp.) in Lithuania. Acta Zool. Lithu. 2006, 16, 241–245. [Google Scholar] [CrossRef]
- Anikanova, V.S.; Bugmyrin, S.B.; Ieshko, E.P. Methods for Collection and Study of Helminths in Small Mammals; Karelian Research Centre of RAS: Petrozavodsk, Russia, 2007; 145p. (In Russian) [Google Scholar]
- Olsen, O.W. Hymenolepis pribilofensis n. sp. (Cestoda: Hymenolepididae) from the Pribilof shrew (Sorex pribilofensis Merriam) from the Pribilof Islands, Alaska. Can. J. Zool. 1969, 47, 449–454. [Google Scholar] [CrossRef]
- Dokuchaev, N.E. The biogeography and taxonomical diversity of the shrews in North-East of Asia. Dokl. Akad. Nauk. 1999, 364, 420–422. (In Russian) [Google Scholar]
- Melnikova, Y.A.; Gulyaev, V.D.; Dokuchaev, N.E. The role of faunistic exchanges in the formation of parasitic-hostal complexes of shrews of Northeast Asia. In Proceedings of the Siberian Zoological Conference, Novosibirsk, Russia, 15–22 September 2004; p. 393. (In Russian). [Google Scholar]
- Dokuchaev, N.E.; Gulyaev, V.D. Shrew (Sorex, Insectivora) as paratenic hosts of cestodes of the genus Mesocestoides. Teriological Res. 2004, 5, 135–138. (In Russian) [Google Scholar]



{kind=link}
{kind=link}
| Cestode Species | Kamchatka | Paramushir | |||
|---|---|---|---|---|---|
| Esso | Azhabachye | Uzon | Yelizovo/Pinachevo | ||
| Family Dilepididae Fuhrmann, 1907 | |||||
| Dilepis undula (larva) (Schrank, 1788) Weinland, 1858 | + | - | - | + | + |
| Monocercus baicalensis (Eltyshev, 1971) Gulyaev and Kornienko, 1998 | + | + | + | + | + |
| Monocercus sp. | + | + | - | + | + |
| Family Hymenolepididae Perrier, 1897 | |||||
| Ecrinolepis collaris (Karpenko, 1984) Gulyaev, 1991 | - | + | + | + | - |
| Lineolepis parva Rausch and Kuns, 1950 | + | + | + | + | + |
| L. pribilofensis Olsen, 1969 | + | - | - | + | + |
| L. skrjabini Spassky and Morosov, 1959 | - | + | - | + | - |
| Lineolepis sp. | - | - | - | + | - |
| Neoskrjabinolepis corticirrosa Kornienko, Gulyaev and Melnikova, 2007 | - | + | + | + | - |
| N. kedrovensis Kornienko, Gulyaev and Melnikova, 2007 | + | + | - | + | + |
| N. longicirrosa Kornienko, Gulyaev and Melnikova, 2006 | + | + | + | + | + |
| N. nadtochijae Kornienko, Gulyaev and Melnikova, 2006 | + | + | - | + | + |
| Pseudobothrialepis mathevossianae Schaldybin, 1957 | + | + | - | + | + |
| Soricinia bargusinica Eltyschev, 1975 | - | + | - | + | - |
| S. infirma (Zarnowsky, 1955) Czaplinski and Vaucher, 1994 | - | + | - | + | - |
| S. quarta (Karpenko, 1983) Karpenko, 1999 | + | + | - | + | + |
| Spasskylepis ovaluteri Schaldybin, 1964 | + | + | + | + | + |
| Staphylocystis sibirica (Morosov, 1957) Spassky and Andrejko, 1970 | + | + | - | + | + |
| Staphylocystis sp. | + | - | - | + | - |
| Staphylocystoides spasskii (Karpenko, 1984) | + | - | - | - | + |
| Urocystis prolifer Villot, 1880 | - | - | - | + | - |
| Family Mesocestoididae Perrier, 1897 | |||||
| Mesocestoides lineatus (larva) (Goeze, 1782) | + | - | + | + | + |
| The number of species | 15 | 15 | 7 | 21 | 14 |
| Cestode Species | Kamchatka | Paramushir | |||||
|---|---|---|---|---|---|---|---|
| S. caecutiens | S. isodon | S. camtschaticus | S. daphaenodon | S. caecutiens | S. isodon | S. leucogaster | |
| D. undula | + | + | - | + | - | + | - |
| E. collaris | + | + | - | + | - | - | - |
| L. parva | + | + | + | + | - | + | - |
| L. pribilofensis | + | - | + | - | + | + | + |
| L. skrjabini | + | + | + | + | - | - | - |
| Lineolepis sp. | + | - | - | - | - | - | - |
| M. lineatus | - | - | + | - | + | - | - |
| M. baicalensis | + | + | - | + | + | + | - |
| Monocercus sp. | - | - | - | + | - | + | - |
| N. corticirrosa | + | - | - | - | - | - | - |
| N. kedrovensis | + | - | - | + | + | + | - |
| N. longicirrosa | + | + | - | + | + | + | - |
| N. nadtochijae | + | + | - | - | + | + | - |
| P. mathevossianae | + | + | - | - | - | + | - |
| S. bargusinica | + | - | - | - | - | - | - |
| S. infirma | + | - | - | - | - | - | - |
| S. quarta | + | + | - | - | + | + | - |
| S. ovaluteri | + | + | - | - | + | + | - |
| S. sibirica | + | - | - | - | + | + | - |
| Staphylocystis sp. | + | - | - | + | - | - | - |
| S. spasskii | + | - | - | - | + | + | - |
| U. prolifer | + | + | - | - | - | - | - |
| The number of species | 20 | 11 | 4 | 9 | 10 | 13 | 1 |
| Shrew Species | Kamchatka | Paramushir | |||
|---|---|---|---|---|---|
| Centre (Esso) | South (Yelizovo/Pinachevo) | East (Uzon) | Total | ||
| S. caecutiens | 62.5 ± 12.1 | 87.8 ± 5.1 | 25 ± 12.5 | 72.5 ± 5.4 | 8 (8) * |
| S. camtschaticus * | - | 3 (1) | 4 (1) | 7 (2) | - |
| S. daphaenodon * | 6 (5) | 4 (4) | - | 10 (9) | - |
| S. isodon | 5 (3) | 77.4 ± 7.5 | 3 (3) * | 76.9 ± 6.7 | 94.2 ± 2.8 |
| S. leucogaster * | - | - | - | - | 1 (1) * |
| Cestode Species | Kamchatka | Paramushir | ||||
|---|---|---|---|---|---|---|
| S. caecutiens | S. isodon | Total | S. isodon | |||
| Uzon | Esso | Yelizovo/Pinachevo | Yelizovo/Pinachevo | |||
| D. undula | - | - | 2.9 ± 2.8 | 16 ± 7.3 | 5.4 ± 2.1 | 6.4 ± 2.8 |
| E. collaris | 25 ± 12.5 | 50 ± 12.5 * | 37.1 ± 8.2 | 8 ± 5.4 | 28.6 ± 4.3 | - |
| L. parva | - | 6.3 ± 6.1 | 5.7 ± 3.9 | 28 ± 8.9 | 12.5 ± 3.1 | 37.2 ± 5.5 |
| L. pribilofensis | - | 6.3 ± 6.1 * | - | - | 2.7 ± 1.5 | 43.6 ± 5.6 ** |
| L. skrjabini | - | 6.3 ± 6.1 * | 22.9 ± 7.1 | 23 ± 9.4 | 18.8 ± 3.7 | - |
| M. baicalensis | - | 6.3 ± 6.1 * | 5.7 ± 3.9 | 32 ± 9.3 | 14.3 ± 3.3 | 15.4 ± 4.1 ** |
| Monocercus sp. | - | - | - | - | 0.9 ± 0.9 * | 32.1 ± 5.3 |
| N. corticirrosa | 25 ± 12.5 | 12.5 ± 8.3 | 14.3 ± 5.9 | - | 8.9 ± 2.7 | - |
| N. kedrovensis | - | 12.5 ± 8.3 | 2.9 ± 2.8 | - | 3.6 ± 1.8 | 2.6 ± 1.8 ** |
| N. longicirrosa | - | 6.3 ± 6.1 * | 31.4 ± 7.8 | 24 ± 8.5 | 19.6 ± 3.8 | 47.4 ± 5.7 ** |
| N. nadtochijae | - | - | 14.3 ± 5.9 | 4 ± 3.9 | 5.4 ± 2.1 | 30.8 ± 5.2 ** |
| P. mathevossianae | - | 12.5 ± 8.3 * | - | - | 2.7 ± 1.5 | 2.6 ± 1.8 |
| S. bargusinica | - | 6.3 ± 6.1 | 2.9 ± 2.8 | - | 1.8 ± 1.3 | - |
| S. infirma | - | 18.8 ± 9.8 | 11.4 ± 5.4 | - | 6.3 ± 2.3 | - |
| S. quarta | - | 6.3 ± 6.1 * | 20 ± 6.8 | 12 ± 6.5 | 10.7 ± 2.9 | 17.9 ± 4.3 ** |
| S. ovaluteri | 16.7 ± 10.8 * | 18.8 ± 9.8 * | 14.3 ± 5.9 | - | 10.7 ± 2.9 | 46.1 ± 5.6 ** |
| S. sibirica | - | - | 2.9 ± 2.8 | - | 0.9 ± 0.9 | 2.6 ± 1.8 ** |
| Staphylocystis sp. | - | 12.5 ± 8.3 | 2.9 ± 2.8 | - | 6.3 ± 2.3 | - |
| S. spasskii | - | 6.3 ± 6.1 | - | - | 0.9 ± 0.9 | 14.1 ± 3.9 ** |
| U. prolifer | - | - | 5.7 ± 3.9 | 4 ± 3.9 | 2.7 ± 1.5 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kornienko, S.A.; Dokuchaev, N.E. Cestode Diversity of Shrews on the Kamchatka Peninsula and Paramushir Island. Diversity 2023, 15, 99. https://doi.org/10.3390/d15010099
Kornienko SA, Dokuchaev NE. Cestode Diversity of Shrews on the Kamchatka Peninsula and Paramushir Island. Diversity. 2023; 15(1):99. https://doi.org/10.3390/d15010099
Chicago/Turabian StyleKornienko, Svetlana A., and Nikolai E. Dokuchaev. 2023. "Cestode Diversity of Shrews on the Kamchatka Peninsula and Paramushir Island" Diversity 15, no. 1: 99. https://doi.org/10.3390/d15010099