
Plastomes Provide Insights into Differences between Morphology and Molecular Phylogeny: Ostericum and Angelica (Apiaceae) as an Example


Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling, DNA Extraction, and Sequencing
2.2. Plastome Assembly and Annotation
2.3. Plastome Comparative Analyses
2.4. Phylogenetic Analyses
3. Results
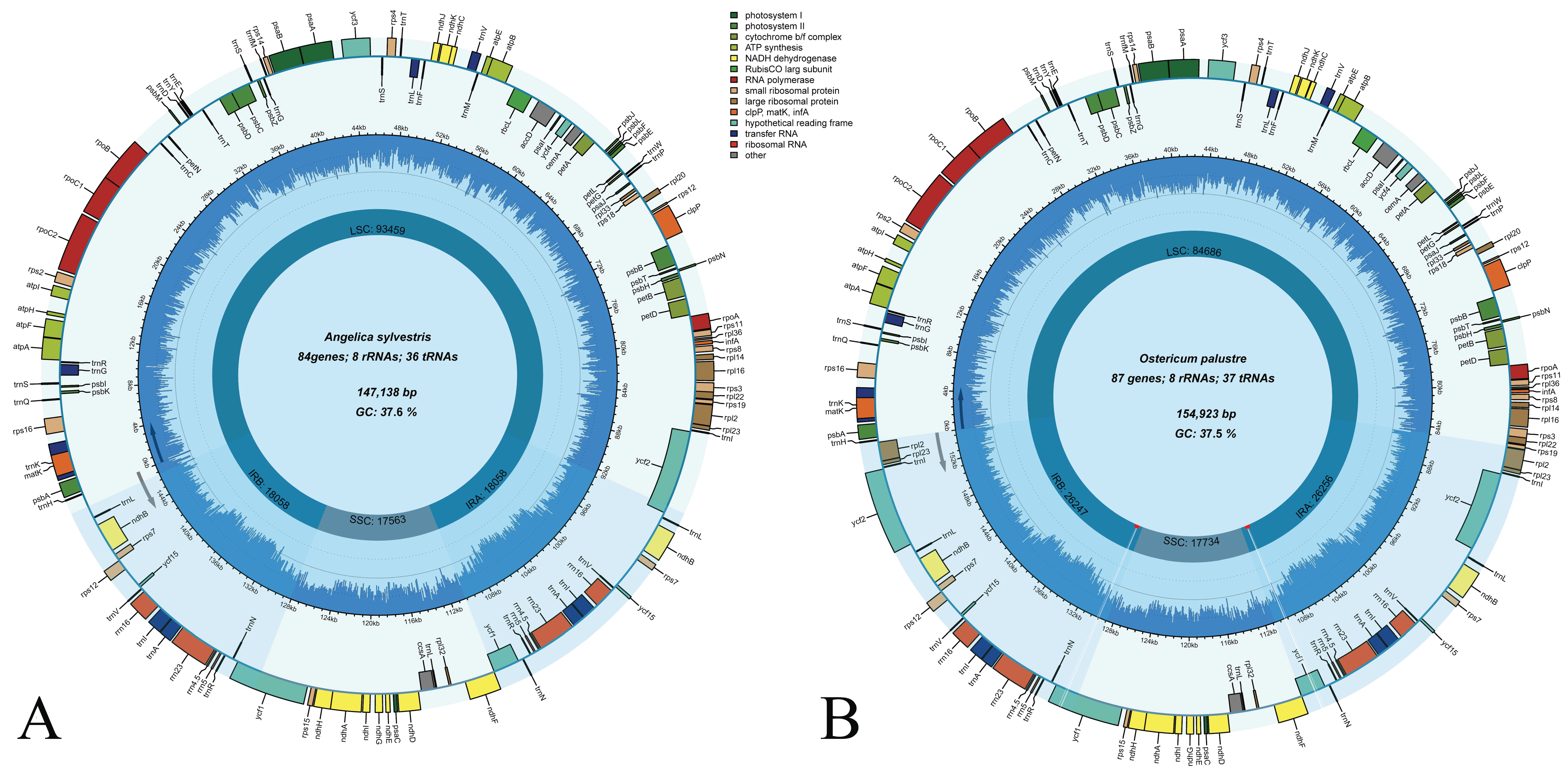
3.1. The Plastome Features of Angelica and Ostericum
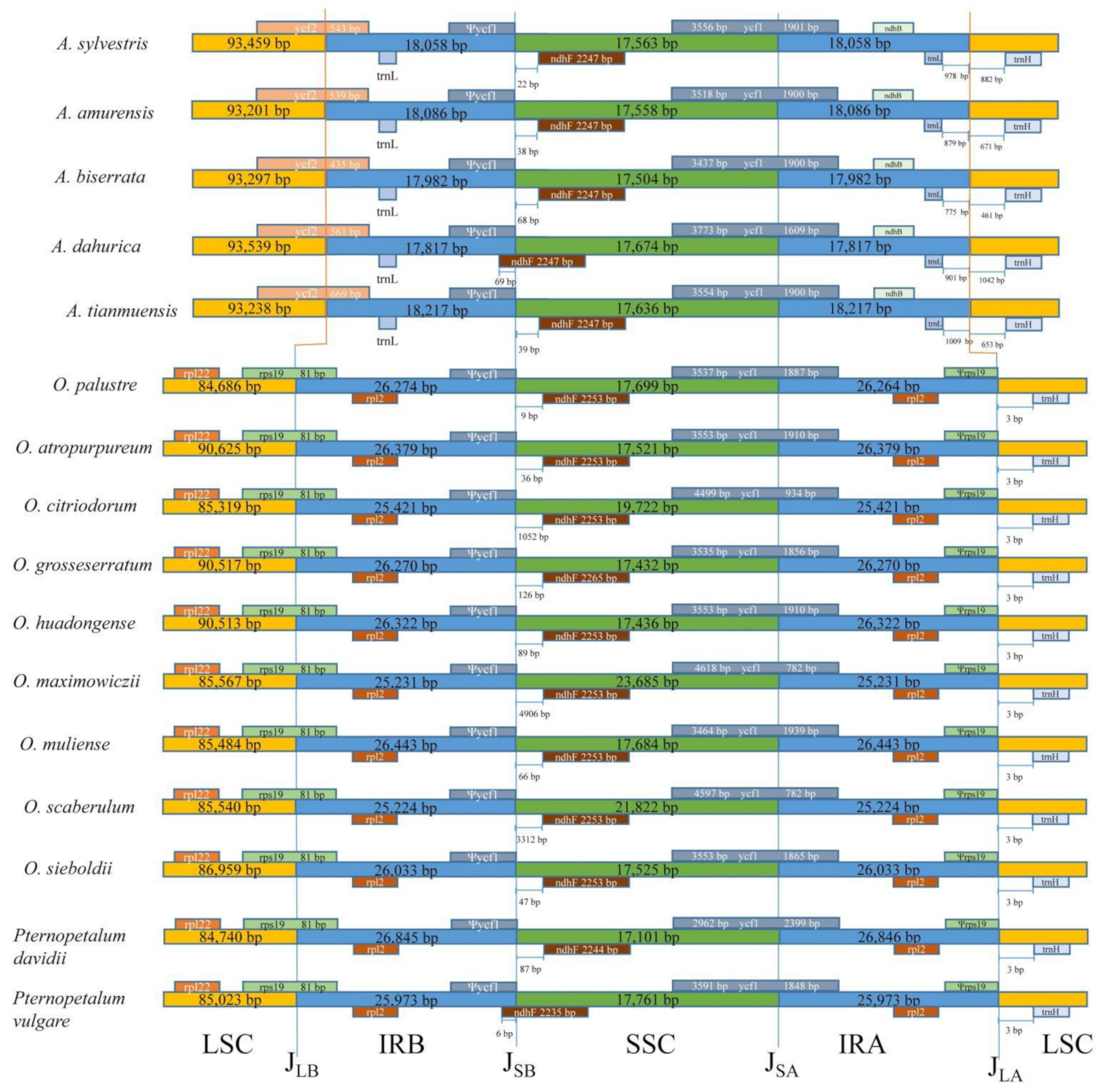
3.2. Analyses of Inverted Repeat Contraction and Expansion
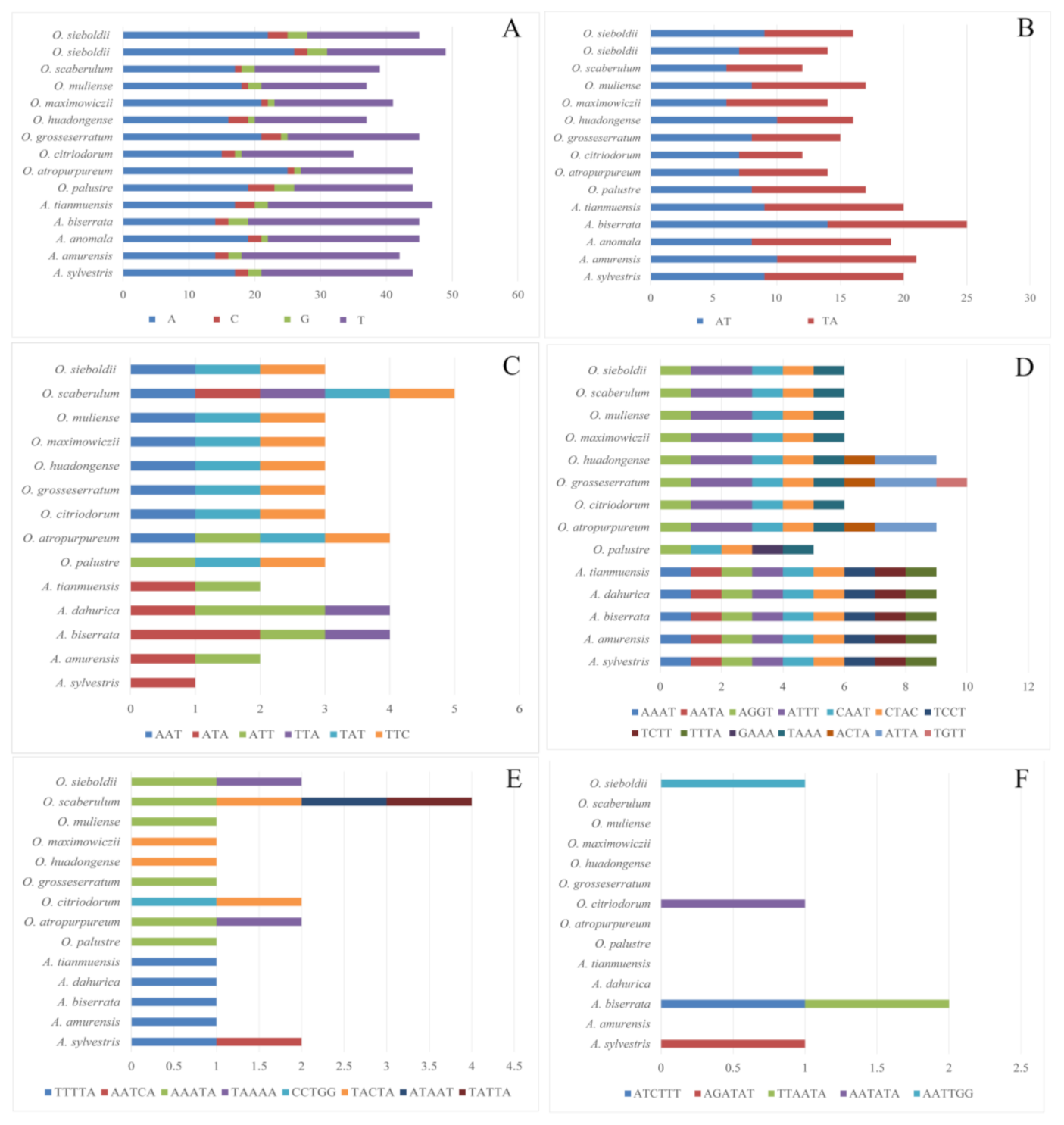
3.3. Single Sequence Repeat (SSR) Analyses
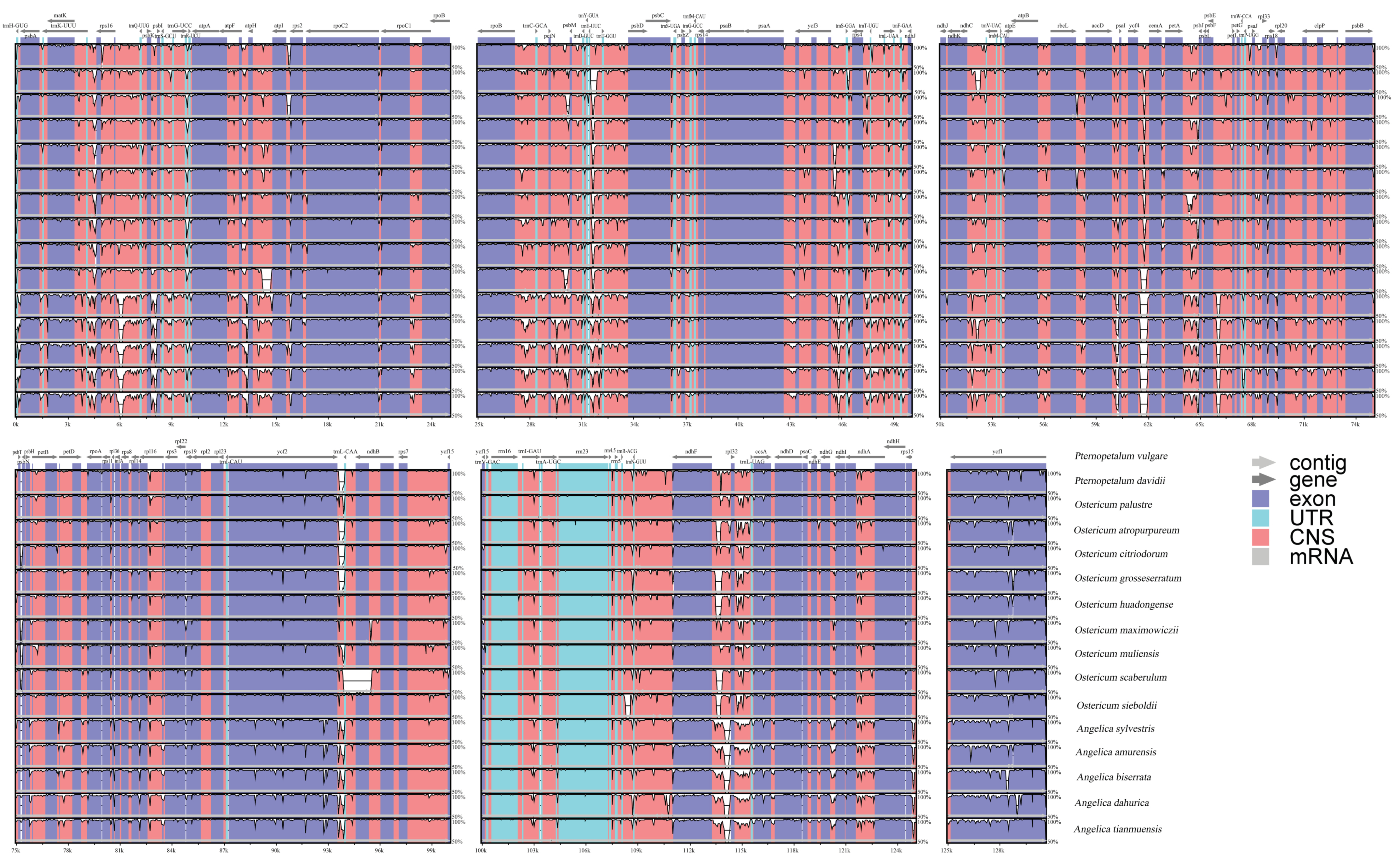
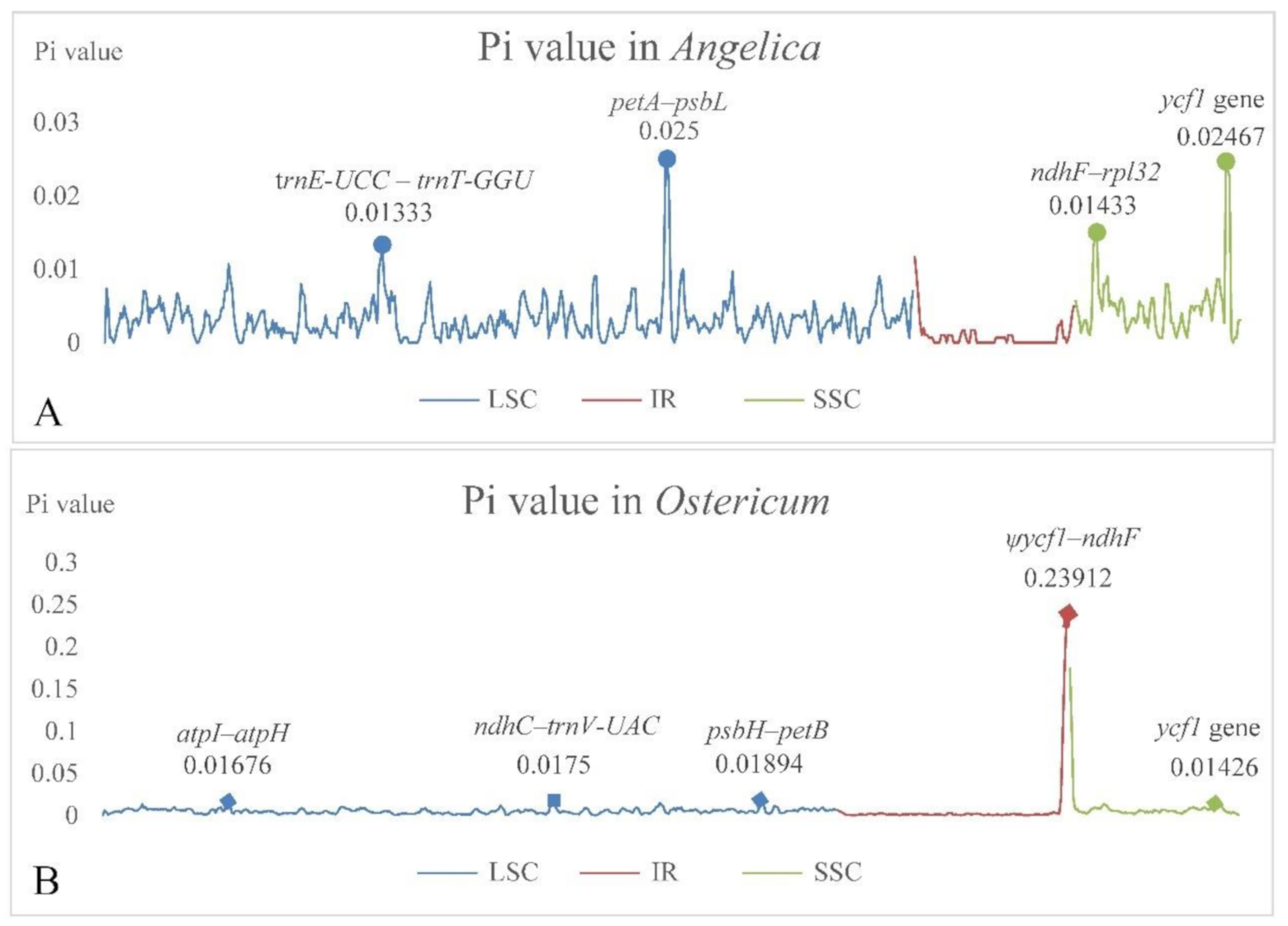
3.4. Sequence Diversity Analyses
3.5. Phylogenetic Analyses
4. Discussion
4.1. Comparative Analyses of Plastomes
4.2. Phylogenetic Position and Intergeneric Relationship of Ostericum
4.3. The Relationship between Angelica and Ostericum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Szöllősi, G.J.; Tannier, E.; Daubin, V.; Boussau, B. The Inference of Gene Trees with Species Trees. Syst. Biol. 2015, 64, e42–e62. [Google Scholar] [CrossRef] [PubMed]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Zhang, Q.; Feild, T.S.; Antonelli, A. Assessing the impact of phylogenetic incongruence on taxonomy, floral evolution, biogeographical history, and phylogenetic diversity. Am. J. Bot. 2015, 102, 566–580. [Google Scholar] [CrossRef]
- Briggs, B.G.; Marchant, A.D.; Perkins, A.J. Phylogeny of the restiid clade (Poales) and implications for the classification of Anarthriaceae, Centrolepidaceae and Australian Restionaceae. Taxon 2014, 63, 24–46. [Google Scholar] [CrossRef]
- Downie, S.R.; Katz-Downie, D.S. A molecular phylogeny of Apiaceae subfamily Apioideae: Evience from nuclear ribosomal DNA internal transcribed spacer sequences. Am. J. Bot. 1996, 83, 234–251. [Google Scholar] [CrossRef]
- Plunkett, G.M.; Downie, S.R. Major lineages within apiaceae subfamily apioideae: A comparison of chloroplast restriction site and dna sequence data. Am. J. Bot. 1999, 86, 1014–1026. [Google Scholar] [CrossRef]
- Downie, S.R.; Spalik, K.; Katz-Downie, D.S.; Reduron, J.-P. Major clades within Apiaceae subfamily Apioideae as inferred by phylogenetic analysis of nrDNA ITS sequences. Plant Divers. Evol. 2010, 128, 111–136. [Google Scholar] [CrossRef]
- Shan, R.H.; Sheh, M.L. Umbelliferae. In Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1992; Volume 55, p. 560. [Google Scholar]
- Sheh, M.L.; Pu, F.T.; Pan, Z.H.; Watson, M.F.; Cannon, J.F.M.; Holmes-Smith, I.; Kljuykov, E.V.; Phillippe, L.R.; Pimenov, M.G. Apiaceae. In Flora of China; Wu, Z.-Y., Raven, P.H., Hong, D.-Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: Saint Louis, MO, USA, 2005; Volume 14, pp. 1–205. [Google Scholar]
- Drude, C.G.O. Umbelliferae. In Die Natürlichen Pflanzenfamilien, Div.; Engler, A., Prantl, K., Eds.; Wilhelm Engelmann: Leipzig, Germany, 1898; Volume 3, p. 187. [Google Scholar]
- Zhou, J.; Peng, H.; Downie, S.R.; Liu, Z.W.; Gong, X. A molecular phylogeny of Chinese Apiaceae subfamily Apioideae inferred from nuclear ribosomal DNA internal transcribed spacer sequences. Taxon 2008, 57, 402–416. [Google Scholar]
- Calviño, C.I.; Teruel, F.E.; Downie, S.R. The role of the Southern Hemisphere in the evolutionary history of Apiaceae, a mostly north temperate plant family. J. Biogeogr. 2016, 43, 398–409. [Google Scholar] [CrossRef]
- Wen, J.; Yu, Y.; Xie, D.F.; Peng, C.; Liu, Q.; Zhou, S.D.; He, X.J. A transcriptome-based study on the phylogeny and evolution of the Taxonomically controversial subfamily Apioideae (Apiaceae). Ann. Bot. 2020, 125, 937–953. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Downie, S.R.; Yu, Y.; Zhang, X.; Chen, W.; He, X.; Liu, S. Molecular systematics of Angelica and allied genera (Apiaceae) from the Hengduan Mountains of China based on nrDNA ITS sequences: Phylogenetic affinities and biogeographic implications. J. Plant Res. 2009, 122, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Downie, S.R.; Li, Q.; Yu, Y.; He, X.; Zhou, B. New Insights into the Phylogeny of Angelica and its Allies (Apiaceae) with Emphasis on East Asian Species, Inferred from nrDNA, cpDNA, and Morphological Evidence. Syst. Bot. 2013, 38, 266–281. [Google Scholar] [CrossRef]
- Pimenov, M.G. Updated checklist of Chinese Umbelliferae: Nomenclature, synonymy, typification, distribution. Turczaninowia 2017, 20, 106–239. [Google Scholar] [CrossRef]
- Pimenov, M.G. Updated checklist of Chinese Umbelliferae: Nomenclature, synonymy, typification, distribution. Supplementum. Turczaninowia 2018, 21, 113–123. [Google Scholar] [CrossRef]
- Mathias, M.E.; Constance, L. North American Flora; The New York Botanical Garden Press: New York, NY, USA, 1944; Volume 28B. [Google Scholar]
- Pimenov, M.G.; Kljuykov, E.V. Notes on some Sino-Himalayan species of Angelica and Ostericum (Umbelliferae). Willdenowia 2003, 33, 121–137. [Google Scholar] [CrossRef]
- Maximowiz, C.J. Angelica sect. Ostericum Maxim. Bull. Acad. Imp. Sci. Saint-Pétersbourg, sér. 1874, 3, 19, 249–276. [Google Scholar]
- Kitagawa, M. Ostericum and Angelica from Manchurica and Korea(Ⅰ). J. Jpn. Bot. 1935, 12, 229–246. [Google Scholar]
- Harborne, J.B.; Heywood, V.H.; Chen, X.Y. Separation of Ostericum from Angelica on the basis of leaf and mericarp flavonoids. Biochem. Syst. Ecol. 1986, 14, 81–83. [Google Scholar] [CrossRef]
- Qin, H.Z.; Li, B.Y.; Wu, Z.J.; Pan, Z.H. On the fruit anatomy of Angelica L. (s.l.) of East Asia and North America and its evolution. Acta Bot. Boreali-Occident. Sin. 1995, 15, 48–54. [Google Scholar]
- Sheh, M.L.; Su, P.; Pan, Z.H. The comparative study of pollen morphology of Angelica L. between East Asia and North America. J. Plant Resour. Environ. 1997, 6, 41–47. [Google Scholar]
- Shu, P.; Sheh, M.L. Ultrastructure of pollen exine in Peucedaneae Drude with reference to its systematic significance. Acta Bot. Sin. 2004, 46, 311–318. [Google Scholar]
- Hiroe, M.; Constance, L. Umbelliferae of Japan; University of California Press: Berkeley, CA, USA, 1958; p. 39. [Google Scholar]
- Downie, S.R.; Ramanath, S.; Katz-Downie, D.S.; Llanas, E. Molecular systematics of Apiaceae subfamily Apioideae: Phylogenetic analyses of nuclear ribosomal DNA internal transcribed spacer and plastid rpoC1 intron sequences. Am. J. Bot. 1998, 85, 563–591. [Google Scholar] [CrossRef]
- Downie, S.R.; Katz-Downie, D.S.; Watson, M.F. A phylogeny of the flowing plant family Apiaceae based on chloroplast DNA rpl16 and rpoC1 intron sequences: Towards a suprageneric classification of subfamily Apioideae. Am. J. Bot. 2000, 87, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S. Molecular systematics of Old World Apioideae (Apiaceae): Relationships among some members of tribe Peucedaneae sensu lato, the placement of several island-endemic species, and resolution within the apioid superclade. Can. J. Bot. 2000, 78, 506–528. [Google Scholar]
- Spalik, K.; Reduron, J.P.; Downie, S.R. The phylogenetic position of Peucedanum sensu lato and allied genera and their placement in tribe Selineae (Apiaceae, subfamily Apioideae). Plant Syst. Evol. 2004, 243, 189–210. [Google Scholar] [CrossRef]
- Schneider, J.V.; Paule, J.; Jungcurt, T.; Cardoso, D.; Amorim, A.M.; Berberich, T.; Zizka, G. Resolving Recalcitrant Clades in the Pantropical Ochnaceae: Insights from Comparative Phylogenomics of Plastome and Nuclear Genomic Data Derived from Targeted Sequencing. Front. Plant Sci. 2021, 12, 638650. [Google Scholar] [CrossRef]
- Asaf, S.; Khan, A.L.; Khan, A.R.; Waqas, M.; Kang, S.M.; Khan, M.A.; Lee, S.M.; Lee, I.J. Complete Chloroplast Genome of Nicotiana otophora and its Comparison with Related Species. Front. Plant Sci. 2016, 7, 843. [Google Scholar] [CrossRef]
- Xie, D.F.; Yu, H.X.; Price, M.; Xie, C.; Deng, Y.Q.; Chen, J.P.; Yu, Y.; Zhou, S.D.; He, X.J. Phylogeny of Chinese Allium Species in Section Daghestanica and Adaptive Evolution of Allium (Amaryllidaceae, Allioideae) Species Revealed by the Chloroplast Complete Genome. Front. Plant Sci. 2019, 10, 460. [Google Scholar] [CrossRef]
- Magallon, S.; Hilu, K.W.; Quandt, D. Land plant evolutionary timeline: Gene effects are secondary to fossil constraints in relaxed clock estimation of age and substitution rates. Am. J. Bot. 2013, 100, 556–573. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Zheng, H.Y.; Price, M.; Zhou, S.D.; He, X.J. Phylogeny and Comparative Analysis of Chinese Chamaesium Species Revealed by the Complete Plastid Genome. Plants 2020, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, D.F.; Guo, X.L.; Zheng, Z.Y.; He, X.J.; Zhou, S.D. Comparative Analysis of the Complete Plastid Genome of Five Bupleurum Species and New Insights into DNA Barcoding and Phylogenetic Relationship. Plants 2020, 9, 543. [Google Scholar] [CrossRef]
- Ren, T.; Li, Z.-X.; Xie, D.-F.; Gui, L.-J.; Peng, C.; Wen, J.; He, X.-J. Plastomes of eight Ligusticum species: Characterization, genome evolution, and phylogenetic relationships. BMC Plant Biol. 2020, 20, 519. [Google Scholar] [CrossRef]
- Xie, D.F.; Yu, Y.; Deng, Y.Q.; Li, J.; Liu, H.Y.; Zhou, S.D.; He, X.J. Comparative Analysis of the Chloroplast Genomes of the Chinese Endemic Genus Urophysa and Their Contribution to Chloroplast Phylogeny and Adaptive Evolution. Int. J. Mol. Sci. 2018, 19, 1847. [Google Scholar] [CrossRef]
- Liao, C.; Chen, X.; Chen, Y.; Gao, Y. The complete chloroplast genome of Angelica sylvestris, the type species of the genus Angelica (Apiaceae). Mitochondrial DNA B Resour. 2019, 4, 3596–3597. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Chen, X.; Tan, J.; Gao, Q. The complete chloroplast genome of Ostericum palustre (Apiaceae). Mitochondrial DNA Part B 2020, 5, 1357–1358. [Google Scholar] [CrossRef]
- Wang, M.; Wang, X.; Sun, J.; Wang, Y.; Ge, Y.; Dong, W.; Yuan, Q.; Huang, L. Phylogenomic and evolutionary dynamics of inverted repeats across Angelica plastomes. BMC Plant Biol. 2021, 21, 26. [Google Scholar] [CrossRef]
- Doyle, J.J. A rapid dna isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. Bot. Soc. Am. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef]
- Qu, X.J.; Moore, M.J.; Li, D.Z.; Yi, T.S. PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Poczai, P.; Hyvonen, J.; Tang, J.; Amiryousefi, A. Chloroplot: An Online Program for the Versatile Plotting of Organelle Genomes. Front. Genet. 2020, 11, 576124. [Google Scholar] [CrossRef] [PubMed]
- Thiel, T.; Michalek, W.; Varshney, R.K.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Bock, R.; Knoop, V. Genomics of Chloroplasts and Mitochondria: Plastid Genomes of Seed Plants; Springer: Berlin/Heidelberg, Germany, 2012; Volume 35, p. 23. [Google Scholar]
- Wicke, S.; Schneeweiss, G.M.; DePamphilis, C.W.; Muller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Jansen, R.K. A Comparative Analysis of Whole Plastid Genomes from the Apiales: Expansion and Contraction of the Inverted Repeat, Mitochondrial to Plastid Transfer of DNA, and Identification of Highly Divergent Noncoding Regions. Syst. Bot. 2015, 40, 336–351. [Google Scholar] [CrossRef]
- Wen, J.; Xie, D.F.; Price, M.; Ren, T.; Deng, Y.Q.; Gui, L.J.; Guo, X.L.; He, X.J. Backbone phylogeny and evolution of Apioideae (Apiaceae): New insights from phylogenomic analyses of plastome data. Mol. Phylogenet. Evol. 2021, 161, 107183. [Google Scholar] [CrossRef]
- Gou, W.; Jia, S.B.; Price, M.; Guo, X.L.; Zhou, S.D.; He, X.J. Complete Plastid Genome Sequencing of Eight Species from Hansenia, Haplosphaera and Sinodielsia (Apiaceae): Comparative Analyses and Phylogenetic Implications. Plants 2020, 9, 1523. [Google Scholar] [CrossRef]
- Liu, C.K.; Lei, J.Q.; Jiang, Q.P.; Zhou, S.D.; He, X.J. The complete plastomes of seven Peucedanum plants: Comparative and phylogenetic analyses for the Peucedanum genus. BMC Plant Biol. 2022, 22, 101. [Google Scholar] [CrossRef]
- Martin, W.; Deusch, O.; Stawski, N.; Grunheit, N.; Goremykin, V. Chloroplast genome phylogenetics: Why we need independent approaches to plant molecular evolution. Trends Plant Sci. 2005, 10, 203–209. [Google Scholar] [CrossRef]
- Raubeson, L.A.; Peery, R.; Chumley, T.W.; Dziubek, C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Comparative chloroplast genomics: Analyses including new sequences from the angiosperms Nuphar advena and Ranunculus macranthus. BMC Genom. 2007, 8, 174. [Google Scholar] [CrossRef]
- Yi, X.; Gao, L.; Wang, B.; Su, Y.J.; Wang, T. The complete chloroplast genome sequence of Cephalotaxus oliveri (Cephalotaxaceae): Evolutionary comparison of cephalotaxus chloroplast DNAs and insights into the loss of inverted repeat copies in gymnosperms. Genome Biol. Evol. 2013, 5, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liu, J.; Zhou, M.; Pan, Y. Chloroplast genome characterization of Bupleurum dracaenoides, a critically endangered woody species endemic to China, with insights of Apioideae phylogeny. Gene Rep. 2020, 20, 100784. [Google Scholar] [CrossRef]
- Li, J.; Price, M.; Su, D.M.; Zhang, Z.; Yu, Y.; Xie, D.F.; Zhou, S.D.; He, X.J.; Gao, X.F. Phylogeny and Comparative Analysis for the Plastid Genomes of Five Tulipa (Liliaceae). Biomed Res. Int. 2021, 2021, 6648429. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.F.; Tan, J.B.; Yu, Y.; Gui, L.J.; Su, D.M.; Zhou, S.D.; He, X.J. Insights into phylogeny, age and evolution of Allium (Amaryllidaceae) based on the whole plastome sequences. Ann. Bot. 2020, 125, 1039–1055. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low Taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef]
- Hu, Y.; Woeste, K.E.; Zhao, P. Completion of the Chloroplast Genomes of Five Chinese Juglans and Their Contribution to Chloroplast Phylogeny. Front. Plant Sci. 2016, 7, 1955. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, M.; Pencakowski, B.; Stafiniak, M.; Jakubowski, K.; Rahimmalek, M.; Gharibi, S.; Matkowski, A.; Slusarczyk, S. Metabolomics and DNA-Based Authentication of Two Traditional Asian Medicinal and Aromatic Species of Salvia subg. Perovskia. Cells 2021, 10, 112. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Mcdevitt, R.; Vendramin, G.G.; Rafalski, J.A. Polymorphic simple sequence repeat regions in chloroplast genomes: Applications to the population genetics of pines. Proc. Natl. Acad. Sci. USA 1995, 92, 7759–7763. [Google Scholar] [CrossRef]
- Gil, J.; Um, Y.; Kim, S.; Kim, O.T.; Koo, S.C.; Reddy, C.S.; Kim, S.C.; Hong, C.P.; Park, S.G.; Kim, H.B.; et al. Development of Genome-Wide SSR Markers from Angelica gigas Nakai Using Next Generation Sequencing. Genes 2017, 8, 238. [Google Scholar] [CrossRef]
- Zalapa, J.E.; Cuevas, H.; Zhu, H.; Steffan, S.; Senalik, D.; Zeldin, E.; McCown, B.; Harbut, R.; Simon, P. Using next-generation sequencing approaches to isolate simple sequence repeat (SSR) loci in the plant sciences. Am. J. Bot. 2012, 99, 193–208. [Google Scholar] [CrossRef]
- Xie, F.M.; Xie, D.F.; Xie, C.; Yu, Y.; Zhou, S.D.; He, X.J. Adaptation Evolution and Phylogenetic Analyses of Species in Chinese Allium Section Pallasia and Related Species Based on Complete Chloroplast Genome Sequences. Biomed Res. Int. 2020, 2020, 8542797. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, D.F.; Chen, J.P.; Zhou, S.D.; Yu, Y.; He, X.J. Comparative Analysis of the Complete Chloroplast Genomes in Allium Subgenus Cyathophora (Amaryllidaceae): Phylogenetic Relationship and Adaptive Evolution. Biomed. Res. Int. 2020, 2020, 1732586. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-Y.; Downie, S.R.; Yu, Y.; He, X.-J. Historical biogeography of the Angelica group (Apiaceae tribe Selineae) inferred from analyses of nrDNA and cpDNA sequences. J. Syst. Evol. 2012, 50, 206–217. [Google Scholar] [CrossRef]
- Gou, W.; Guo, X.L.; Zhou, S.D.; He, X.J. Phylogeny and Taxonomy of Meeboldia, Sinodielsia and their relatives (Apiaceae: Apioideae) inferred from nrDNA ITS, plastid DNA intron (rpl16 and rps16) sequences and morphological characters. Phytotaxa 2021, 482, 121–142. [Google Scholar] [CrossRef]
- Yuan, C.Q.; Shan, R.H. On the genera Angelica L. and Ostericum Hoffm. (Umbelliferae) in China. Bull. Nanjing Bot. Gard. 1985, 1984–1985, 1–5. [Google Scholar]
- Pan, Z.H.; Waston, M.F. Ostericum Hoffm. In Flora of China; Wu, Z.Y., Raven, R.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: Saint Louis, MO, USA, 2005; Volume 14, p. 3. [Google Scholar]
- Stewart, J.E.; Timmer, L.W.; Lawrence, C.B.; Pryor, B.M.; Peever, T.L. Discord between morphological and phylogenetic species boundaries: Incomplete lineage sorting and recombination results in fuzzy species boundaries in an asexual fungal pathogen. BMC Evol. Biol. 2014, 14, 38. [Google Scholar] [CrossRef] [PubMed]
- Davalos, L.M.; Cirranello, A.L.; Geisler, J.H.; Simmons, N.B. Understanding phylogenetic incongruence: Lessons from phyllostomid bats. Biol. Rev. Camb. Philos. Soc. 2012, 87, 991–1024. [Google Scholar] [CrossRef]
- Feng, S.; Bai, M.; Rivas-Gonzalez, I.; Li, C.; Liu, S.; Tong, Y.; Yang, H.; Chen, G.; Xie, D.; Sears, K.E.; et al. Incomplete lineage sorting and phenotypic evolution in marsupials. Cell 2022, 185, 1646–1660. [Google Scholar] [CrossRef]
- Yang, M.-Q.; Van Velzen, R.; Bakker, F.T.; Sattarian, A.; Li, D.-Z.; Yi, T.S. Molecular phylogenetics and character evolution of Cannabaceae. Taxon 2013, 62, 473–485. [Google Scholar] [CrossRef]
- Bouman, R.W.; Keßler, P.J.A.; Telford, I.R.H.; Bruhl, J.J.; Strijk, J.S.; Saunders, R.M.K.; Welzen, P.C. Molecular phylogenetics of Phyllanthus sensu lato (Phyllanthaceae): Towards coherent monophyletic taxa. Taxon 2020, 70, 72–98. [Google Scholar] [CrossRef]
- Feng, C.M.; Manchester, S.R.; Xiang, Q.Y. Phylogeny and biogeography of Alangiaceae (Cornales) inferred from DNA sequences, morphology, and fossils. Mol. Phylogenet. Evol. 2009, 51, 201–214. [Google Scholar] [CrossRef]
- Chase, M.W. Monocot Relationships: An Overview. Am. J. Bot. 2004, 91, 1645–1655. [Google Scholar] [CrossRef]
- Ren, T.; Xie, D.; Peng, C.; Gui, L.; Price, M.; Zhou, S.; He, X. Molecular evolution and phylogenetic relationships of Ligusticum (Apiaceae) inferred from the whole plastome sequences. BMC Ecol. Evol. 2022, 22, 55. [Google Scholar] [CrossRef]
- Akbarian, A.; Rahimmalek, M.; Sabzalian, M.R.; Hodaei, M. Sequencing and phylogenetic analysis of phenylalanine ammonia lyase (pal) and chalcone synthase (chs) genes in some Iranian endemic species of Apiaceae. Gene Rep. 2021, 23, 101147. [Google Scholar] [CrossRef]
- Pan, Z.H.; Zhuang, T.D.; Yao, X.M.; Sheng, N. A study on karyotypes and geographical distribution of Angelica and related genera (Umbelliferae) in China. Acta Phytotax. Sin. 1994, 32, 419–424. [Google Scholar]
- Pan, Z.H.; Liu, X.T.; Li, X.H.; Yao, X.M.; Zhuang, T.D.; Lin, X. A Study on Karyotypes and Geographical Distribution of Ostericum (Umbelliferae) in China. Acta Phytotax. Sin. 1997, 35, 511–520. [Google Scholar]
- Degnan, J.H.; Rosenberg, N.A. Gene tree discordance, phylogenetic inference and the multispecies coalescent. Trends Ecol. Evol. 2009, 24, 332–340. [Google Scholar] [CrossRef]
- Cranston, K.A.; Hurwitz, B.; Ware, D.; Stein, L.; Wing, R.A. Species trees from highly incongruent gene trees in rice. Syst. Biol. 2009, 58, 489–500. [Google Scholar] [CrossRef]
- Sackton, T.B.; Clark, N. Convergent evolution in the genomics era: New insights and directions. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190102. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Size (bp) | LSC Length (bp) | SSC Length (bp) | IR Length (bp) | Number of Different Genes/Total Number of Genes | Number of Different Protein-Coding Genes (Duplicated in IR) | Number of Different tRNA Genes (Duplicated in IR) | Number of Different rRNA Genes (Duplicated in IR) | Number of Genes Duplicated in IR | GC Content (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| A. sylvestris | 147,138 | 93,459 | 17,563 | 18,058 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.5 |
| A. amurensis | 146,931 | 93,201 | 17,558 | 18,086 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.5 |
| A. biserrata | 146,765 | 93,297 | 17,504 | 17,982 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.5 |
| A. dahurica | 146,847 | 93,539 | 17,674 | 17,817 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.5 |
| A. tianmuensis | 147,308 | 93,238 | 17,636 | 18,217 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.5 |
| O. palustre | 154,923 | 84,686 | 17,699 | 26,274 (26,264) | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.5 |
| O. atropurpureum | 160,904 | 90,625 | 17,521 | 26,379 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.7 |
| O. citriodorum | 155,883 | 85,319 | 19,722 | 25,421 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.6 |
| O. grosseserratum | 160,489 | 90,517 | 17,432 | 26,270 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.7 |
| O. huadongense | 160,489 | 90,517 | 17,432 | 26,270 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.7 |
| O. maximowiczii | 159,714 | 85,567 | 23,685 | 25231 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.7 |
| O. muliense | 156,054 | 85,484 | 17,684 | 26,443 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.4 |
| O. scaberulum | 157,810 | 85,540 | 21,822 | 25,224 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.6 |
| O. sieboldii | 156,550 | 86,959 | 17,525 | 26,033 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.6 |
| P. davidii | 155,533 | 84,741 | 17,101 | 26,845 (26,846) | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.5 |
| P. vulgare | 154,730 | 85,023 | 17,761 | 25,973 | 114/134 | 80 (7) | 30 (7) | 4 (4) | 20 | 37.5 |
| Category | Gene Group | Gene Name |
|---|---|---|
| Self-replication | Transfer RNA genes | trnI-CAU (*), trnI-GAU *, trnL-UAA, trnL-CAA *, trnL-UAG, trnR-UCU, trnR-ACG *, trnA-UGC *, trnW-CCA, trnM-CAU, trnV-UAC, trnV-GAC *, trnF-GAA, trnT-UGU, trnT-GGU, trnP-UGG, trnfM-CAU, trnG-UCC, trnG-GCC, trnS-GGA, trnS-UGA, trnS-GCU, trnD-GUC, trnC-GCA, trnN-GUU *, trnE-UUC, trnY-GUA, trnQ-UUG, trnK-UUU, trnH-GUG |
| Ribosomal RNA genes | rrn16 *, rrn23 *, rrn4.5 *, rrn5 * | |
| RNA polymerase | rpoA, rpoB, rpoC1, rpoC2 | |
| Small subunit of ribosome | rps2, rps3, rps4, rps7 *, rps8, rps11, rps12 *, rps14, rps15, rps16, rps18, rps19 (*) (rps19, ψrps19) | |
| Large subunit of ribosomal proteins (LSU) | rpl2 (*), rpl14, rpl16, rpl20, rpl22, rpl23 (*), rpl32, rpl33, rpl36 | |
| Genes for photosynthesis | Subunits of NADH-dehydrogenase | ndhA, ndhB *, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK |
| Subunits of photosystem I | psaA, psaB, psaC, psaI, psaJ | |
| Subunits of photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | |
| Subunits of cytochrome | petA, petB, petD, petG, petL, petN | |
| Subunits of ATP synthase | atpA, atpB, atpE, atpF, atpH, atpI | |
| Large subunit of rubisco | rbcL | |
| Other genes | Translational initiation factor | infA |
| Protease | clpP | |
| Maturase | matK | |
| C-type cytochrome synthesis gene | ccsA | |
| Subunit of acetyl-CoA | accD | |
| Envelope membrane protein | cemA | |
| Conserved open reading frames (ycf) | ycf1 * (ycf1, ψycf1), ycf2 (*), ycf3, ycf4, ψycf15 * | |
| Total | Angelica: 129, Ostericum: 134 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.-P.; Liu, C.-K.; Xie, D.-F.; Zhou, S.-D.; He, X.-J. Plastomes Provide Insights into Differences between Morphology and Molecular Phylogeny: Ostericum and Angelica (Apiaceae) as an Example. Diversity 2022, 14, 776. https://doi.org/10.3390/d14090776
Jiang Q-P, Liu C-K, Xie D-F, Zhou S-D, He X-J. Plastomes Provide Insights into Differences between Morphology and Molecular Phylogeny: Ostericum and Angelica (Apiaceae) as an Example. Diversity. 2022; 14(9):776. https://doi.org/10.3390/d14090776
Chicago/Turabian StyleJiang, Qiu-Ping, Chang-Kun Liu, Deng-Feng Xie, Song-Dong Zhou, and Xing-Jin He. 2022. "Plastomes Provide Insights into Differences between Morphology and Molecular Phylogeny: Ostericum and Angelica (Apiaceae) as an Example" Diversity 14, no. 9: 776. https://doi.org/10.3390/d14090776