


Diversity Patterns of Wetland Angiosperms in the Qinghai-Tibet Plateau, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Species Checklist and Collection of Distribution Data
- (i)
- Criteria for evaluating wetland plants: According to the relationship between plants and water, wetland plants are divided into two categories: hydrophytes (high moisture dependence group) and hygrophytes (low moisture dependence group). Hydrophytes were composed of emergent, floating-leaf, floating and submerged plants, which refer to plants living in water, whereas hygrophytes were composed of wet and marshy plants, which are less dependent on water than hydrophytes. They refer to a class of plants that grow in swamps or humid environments. We use the following habitat description keywords as the judgment criteria for hygrophytes: river beach, ditch edge, stream edge, river edge, lake edge, pond, moss bush, tundra, wet grassland, shady and wet place under the forest, shady and wet place in the valley, shady and wet place on the hillside, floodplain, swamp, paddy field, salt lake edge, near the snow line.
- (ii)
- According to monographs, research papers, online databases, and field survey data, the list and distribution map of wetland angiosperms in the Qinghai-Tibet Plateau were compiled. These data sources are as follows: The Vascular Plants and Their Ecogeographical Distribution of the Qinghai-Tibetan Plateau [34], Flora Reipublicae Popularis Sinicae [35], Flora of China [36], floras of Tibet, Qinghai, Sichuan, Yunnan, Gansu and Xinjiang, China Wetlands Resources (Qinghai Volume, Tibet Volume) [17,18]. We also supplemented a large number of specimen data and online data, such as National Specimen Information Infrastructure (http://www.nsii.org.cn, accessed on 27 January 2022), Global Biodiversity Information Facility (https://www.gbif.org/, accessed on 22 February 2022), Chinese Virtual Herbarium (https://www.cvh.ac.cn/, accessed on 25 February 2022), Plant Photo Bank of China (http://ppbc.iplant.cn, accessed on 1 May 2021). These monographs and specimen data are a large number of investigations and arrangements made by many scientific researchers on the plants and their distribution on the Qinghai-Tibet Plateau in the past 60 years [37]. We used the following database to calibrate the directory to ensure the accuracy of the directory: China Wetland Plant Database (http://zgsdzw.com, accessed on 1 May 2021), China Aquatic Plant Database (http://www.plant.csdb.cn/aquaticplants, accessed on 14 March 2022). Finally, the field survey data from 2018 to 2020 are supplemented.
- (iii)
- In order to ensure the consistency of the naming of all databases, we used “Species 2000 China Node” (http://www.sp2000.org.cn, accessed on 8 March 2022) [38] and R package “plantlist” to calibrate the checklist of wetland plants on the Qinghai-Tibet Plateau [39]. We recorded the life form, altitude range, habitat information, and county-level distribution points of each species in detail. We used the national Qinghai-Tibet Plateau scientific data center Qinghai-Tibet Plateau urban distribution and urbanization index data set (2018, 2019) to determine the scope of the geographical area of the Qinghai-Tibet Plateau [40].
- (iv)
- According to the Chinese Biodiversity Redlist of Higher Plants, we have compiled the checklist of endangered wetland angiosperms in the Qinghai-Tibet Plateau, and mapped the distribution of endangered wetland angiosperms [41].
2.2. Environmental Variables
2.3. Statistical Analysis
3. Results
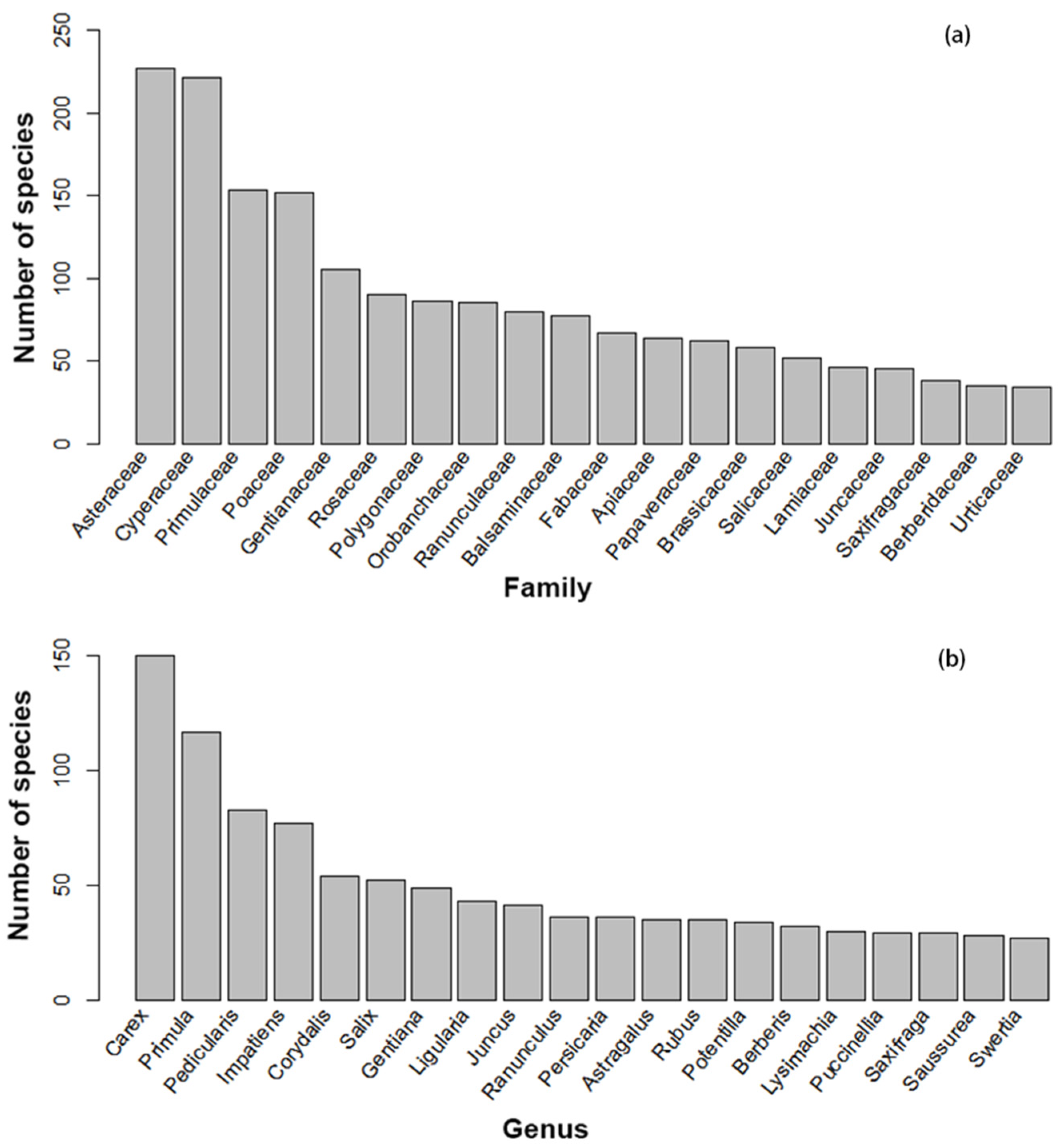
3.1. Families, Genera, Species, and Life Forms of Wetland Angiosperms
3.2. County Level Distribution Pattern of Species Richness and Species Density
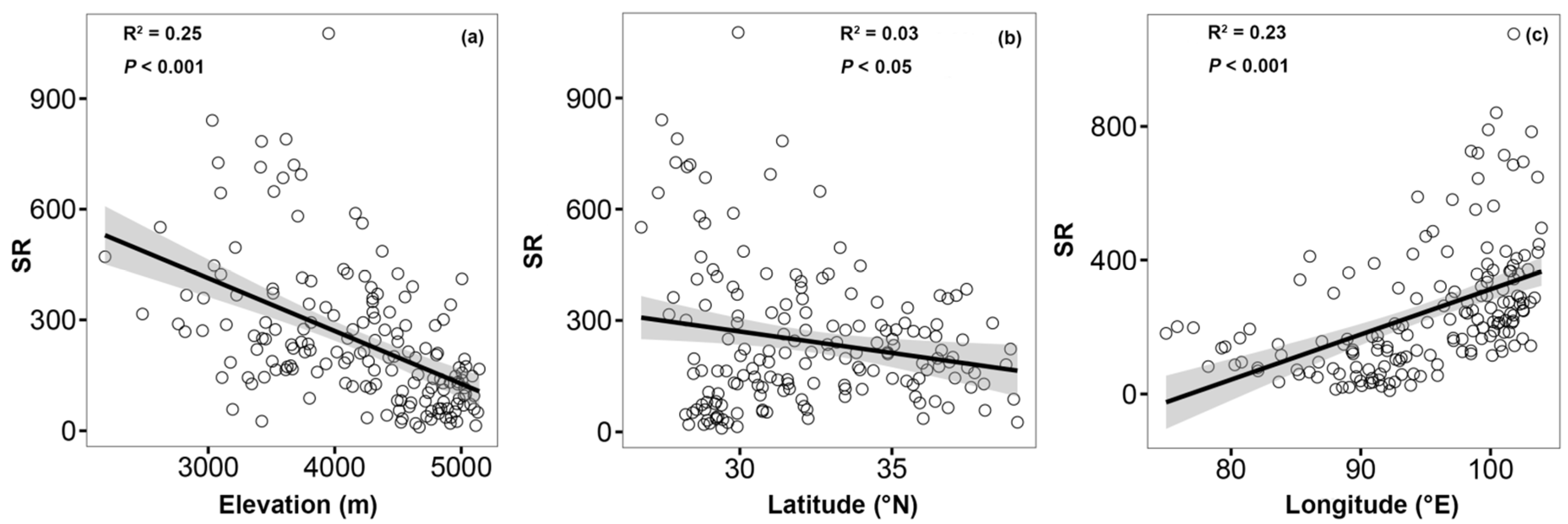
3.3. Elevation, Longitude, and Latitude Patterns of Species Richness
3.4. Relationship between Environmental Variables and Species Richness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.; Song, W. Spatiotemporal Distribution and Influencing Factors of Ecosystem Vulnerability on Qinghai-Tibet Plateau. Int. J. Environ. Res. Public Health 2021, 18, 6508. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Duan, Y.; Hao, G.; Ge, X.; Sun, H. Evolutionary history and underlying adaptation of alpine plants on the Qinghai-Tibet Plateau. J. Syst. Evol. 2014, 52, 241–249. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Liu, Z.; Peng, Y.; Liu, W. Retrieval algorithm for microwave surface emissivities based on multi-source, remote-sensing data: An assessment on the Qinghai-Tibet Plateau. Sci. China Earth Sci. 2013, 56, 93–101. [Google Scholar] [CrossRef]
- Yu, H.; Deane, D.C.; Zhang, Y.; Li, S.; Miao, S.; Xie, G.; Yin, X.; Favre, A. Integrating multiple indices of geobiodiversity reveals a series of regional species-rich areas worthy of conservation in the region of the Qinghai-Tibet Plateau. Biol. Conserv. 2021, 261, 109238. [Google Scholar] [CrossRef]
- Fu, Y.; Gao, X.; Zhu, Y.; Guo, D. Climate change projection over the Tibetan Plateau based on a set of RCM simulations. Adv. Clim. Chang. Res. 2021, 12, 313–321. [Google Scholar] [CrossRef]
- Gao, J.; Li, X.; Brierley, G.; Cheung, A.; Yang, Y. Geomorphic-centered classification of wetlands on the Qinghai-Tibet Plateau, Western China. J. Mt. Sci.-Engl. 2013, 10, 632–642. [Google Scholar] [CrossRef]
- Yu, H.; Xu, J.; Okuto, E.; Luedeling, E.; Wright, J. Seasonal response of grasslands to climate change on the Tibetan Plateau. PLoS ONE 2012, 7, e49230. [Google Scholar] [CrossRef]
- Gao, J.; Li, X.; Cheung, A.; Yang, Y. Degradation of wetlands on the Qinghai-Tibet Plateau: A comparison of the effectiveness of three indicators. J. Mt. Sci. Engl. 2013, 10, 658–667. [Google Scholar] [CrossRef]
- Wan, W.; Gadd, G.M.; Yang, Y.; Yuan, W.; Gu, J.; Ye, L.; Liu, W. Environmental adaptation is stronger for abundant rather than rare microorganisms in wetland soils from the Qinghai-Tibet Plateau. Mol. Ecol. 2021, 30, 2390–2403. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Liu, L.; Liu, F.; Zhang, H. Recent changes in wetlands on the Tibetan Plateau: A review. J. Geogr. Sci. 2015, 25, 879–896. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, T.; Sun, W.; Zhang, W.; Zhang, Q.; Yu, L.; Qin, Z.; Guo, B.; Liu, J.; Zha, X. Degradation of wetlands on the Qinghai-Tibetan Plateau causing a loss in soil organic carbon in 1966–2016. Plant Soil 2021, 467, 253–265. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, G.; Wang, Y. Changes in alpine wetland ecosystems of the Qinghai–Tibetan plateau from 1967 to 2004. Environ. Monit. Assess. 2011, 180, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xue, Z.; Gao, J. Environmental Influence on Vegetation Properties of Frigid Wetlands on the Qinghai-Tibet Plateau, Western China. Wetlands 2016, 36, 807–819. [Google Scholar] [CrossRef]
- Zhao, K.Y. The Characteristics of swamp vegetation on Xizang Plateau. Sci. Geogr. Sin. 1982, 2, 73–82. [Google Scholar]
- Chen, G.-C.; Huang, Z.-W.; Lu, X.-F.; Peng, M. Characteristics of Wetland and Its Conservation in the Qinghai Plateau. J. Glaciol. Geocryol. 2002, 24, 254–259. [Google Scholar]
- Liu, W.L.; Zhu, X.L. Plateau Wetland in Tibet of China; China Forestry Publishing House: Beijing, China, 2013; pp. 1–424. [Google Scholar]
- State Forestry Administration. China Wetlands Resources_Qinghai Volume; China Forestry Publishing House: Beijing, China, 2015; pp. 1–306.
- State Forestry Administration. China Wetlands Resources_Tibet Volume; China Forestry Publishing House: Beijing, China, 2015; pp. 1–171.
- Pandey, B.; Khatiwada, J.R.; Zhang, L.; Pan, K.; Dakhil, M.A.; Xiong, Q.; Yadav, R.K.P.; Siwakoti, M.; Tariq, A.; Olatunji, O.A.; et al. Energy–water and seasonal variations in climate underlie the spatial distribution patterns of gymnosperm species richness in China. Ecol. Evol. 2020, 10, 9474–9485. [Google Scholar] [CrossRef]
- Currie, D.J.; Paquin, V. Large-scale biogeographical patterns of species richness of trees. Nature 1987, 329, 326–327. [Google Scholar] [CrossRef]
- O’Brien, E. Water-energy dynamics, climate, and prediction of woody plant species richness: An interim general model. J. Biogeogr. 1998, 25, 379–398. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, Water, and Broad-Scale Geographic Patterns of Species Richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Shrestha, N.; Su, X.; Xu, X.; Wang, Z. The drivers of high Rhododendron diversity in south-west China: Does seasonality matter? J. Biogeogr. 2018, 45, 438–447. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Fjeldså, J.; Bowie, R.C.K.; Rahbek, C. The Role of Mountain Ranges in the Diversification of Birds. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Currie, D.J. Energy and Large-Scale Patterns of Animal- and Plant-Species Richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Francis, A.P.; Currie, D.J. A globally consistent richness-climate relationship for angiosperms. Am. Nat. 2003, 161, 523–536. [Google Scholar] [CrossRef]
- Cui, S.; Ouyang, J.; Lu, Y.; Liu, W.; Li, W.; Liu, G.; Zhou, W. The Diversity and Community Assembly Process of Wetland Plants from Lakeshores on the Qinghai-Tibetan Plateau. Diversity 2021, 13, 685. [Google Scholar] [CrossRef]
- Jones, J.I.; Li, W.; Maberly, S.C. Area, altitude and aquatic plant diversity. Ecography 2003, 26, 411–420. [Google Scholar] [CrossRef]
- Zhou, Y.D.; Xiao, K.Y.; Chen, S.C.; Liu, X.; Wang, Q.F.; Yan, X. Altitudinal diversity of aquatic plants in the Qinghai-Tibet Plateau. Freshw. Biol. 2022, 67, 709–719. [Google Scholar] [CrossRef]
- Griffiths, D.; Mcgonigle, C.; Quinn, R. Climate and species richness patterns of freshwater fish in North America and Europe. J. Biogeogr. 2014, 41, 452–463. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, Q.; Yang, Y.; Zhang, X.; Zhang, J.; Yuan, M.; Chen, H.; Peng, C. High uncertainties detected in the wetlands distribution of the Qinghai–Tibet Plateau based on multisource data. Landsc. Ecol. Eng. 2020, 16, 47–61. [Google Scholar] [CrossRef]
- Russell, K.N.; Beauchamp, V.B. Plant Species Diversity in Restored and Created Delmarva Bay Wetlands. Wetlands 2017, 37, 1119–1133. [Google Scholar] [CrossRef]
- Wu, Y. The Vascular Plants and Their Eco-Geographical Distribution of the Qinghai-Tibetan Plateau; Science Press: Beijing, China, 2008. [Google Scholar]
- Editorial Committee of Flora Reipublicae Popularis Sinicae. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1959–2004. [Google Scholar]
- Wu, Z.; Raven, P.H.; Hong, D. Flora of China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1994–2013. [Google Scholar]
- Yan, Y.; Yang, X.; Tang, Z. Patterns of species diversity and phylogenetic structure of vascular plants on the Qinghai-Tibetan Plateau. Ecol. Evol. 2013, 3, 4584–4595. [Google Scholar] [CrossRef] [PubMed]
- The Biodiversity Committee of Chinese Academy of Sciences. Catalogue of Life China: 2022 Annual Checklist; Academy of Sciences: Beijing, China, 2022. [Google Scholar]
- Zhang, J.L. Plantlist: Looking up Families of Higher Plants. 2015. Available online: http://R-Forge.R-project.org/projects/plantlist/ (accessed on 11 May 2022).
- He, C.; Liu, Z.; Wang, Y. Dataset of Urban Land and Urbanization Index on the Tibetan Plateau (2018, 2019); National Tibetan Plateau Data Center: Beijing, China, 2020. [Google Scholar]
- Ministry of Environmental Protection (MEP) of the People’s Republic of China and the Chinese Academy of Sciences. The Chinese Biodiversity Redlist-Higher Plants Volume. 2015. Available online: http://www.mee.gov.cn/gkml/hbb/bgg/201309/W020130917614244055331.pdf (accessed on 30 June 2022).
- Mao, D.; Wang, Z.; Du, B.; Li, L.; Tian, Y.; Jia, M.; Zeng, Y.; Song, K.; Jiang, M.; Wang, Y. National wetland mapping in China: A new product resulting from object-based and hierarchical classification of Landsat 8 OLI images. ISPRS J. Photogramm. 2020, 164, 11–25. [Google Scholar] [CrossRef]
- Li, R.; Kraft, N.J.B.; Yu, H.; Li, H. Seed plant phylogenetic diversity and species richness in conservation planning within a global biodiversity hotspot in eastern Asia. Conserv. Biol. 2015, 29, 1552–1562. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, X.; Qian, H.; Huang, X.; Liu, P.; Landis, J.B.; Fu, Q.; Sun, L.; Wang, H.; Sun, H.; et al. Elevational patterns of phylogenetic structure of angiosperms in a biodiversity hotspot in eastern Himalaya. Divers. Distrib. 2022, 1–15. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Xiang, S.; Guo, R.; Wu, N.; Sun, S. Current status and future prospects of Zoige Marsh in Eastern Qinghai-Tibet Plateau. Ecol. Eng. 2009, 35, 553–562. [Google Scholar] [CrossRef]
- Wang, D. The Geography of Aquatic Vascular Plants of Qinghai-Xizang(Tibet) Plateau. Ph.D. Thesis, Wuhan University, Wuhan, China, 2003. [Google Scholar]
- Zhang, A.; Hu, X.; Yao, S.; Yu, M.; Ying, Z. Alien, Naturalized and Invasive Plants in China. Plants 2021, 10, 2241. [Google Scholar] [CrossRef]
- Yu, H.B.; Zhang, Y.L.; Liu, L.S.; Chen, Z.; Qi, W. Floristic characteristics and diversity patterns of seed plants endemic to the Tibetan Plateau. Biodiv. Sci. 2018, 26, 130–137. [Google Scholar] [CrossRef]
- Liu, J. Uniformity of karyotypes in Ligularia (Asteraceae: Senecioneae), a highly diversified genus of the eastern Qinghai-Tibet Plateau highlands and adjacent areas. Bot. J. Linn. Soc. 2004, 144, 329–342. [Google Scholar] [CrossRef]
- Wen, J.; Zhang, J.; Nie, Z.; Zhong, Y.; Sun, H. Evolutionary diversifications of plants on the Qinghai-Tibetan Plateau. Front. Genet. 2014, 5, 4. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2014, 29, 592–599. [Google Scholar] [CrossRef]
- Ji, W.L. Study on Ecophysiological Responses of Sedges (Carex: Cyperaceae) to Heterogeneous Environment. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2008. [Google Scholar]
- Wu, Z.; Xu, X.; Zhang, J.; Wiegleb, G.; Hou, H. Influence of environmental factors on the genetic variation of the aquatic macrophyte Ranunculus subrigidus on the Qinghai-Tibetan Plateau. BMC Evol. Biol. 2019, 19, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kull, T.; Selgis, U.; Pecina, M.V.; Metsare, M.; Ilves, A.; Tali, K.; Sepp, K.; Kull, K.; Shefferson, R.P. Factors influencing IUCN threat levels to orchids across Europe on the basis of national red lists. Ecol. Evol. 2016, 6, 6245–6265. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Z.; Jin, X.; Gao, J.; Chen, Y.; Liu, Q.; Zhang, D. Assessing conservation efforts against threats to wild orchids in China. Biol. Conserv. 2020, 243, 108484. [Google Scholar] [CrossRef]
- Wraith, J.; Pickering, C. Quantifying anthropogenic threats to orchids using the IUCN Red List. Ambio 2018, 47, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Wang, Z.; Ye, L. Integrative Taxonomy Supports Two New Species of Rhodiola (Crassulaceae) in Xizang, China. Diversity 2022, 14, 289. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, D.; Chen, S.; Xia, T.; Gao, Q.; Duan, Y.; Zhang, F. Determination of Salidroside in Medicinal Plants Belonging to the Rhodiola L. Genus Originating from the Qinghai–Tibet Plateau. Chromatographia 2008, 68, 299–302. [Google Scholar] [CrossRef]
- Yu, H.; Miao, S.; Xie, G.; Guo, X.; Chen, Z.; Favre, A. Contrasting Floristic Diversity of the Hengduan Mountains, the Himalayas and the Qinghai-Tibet Plateau Sensu Stricto in China. Front. Ecol. Evol. 2020, 8, 136. [Google Scholar] [CrossRef]
- Rolon, A.S.; Maltchik, L. Environmental Factors as Predictors of Aquatic Macrophyte Richness and Composition in Wetlands of Southern Brazil. Hydrobiologia 2006, 556, 221–231. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal Gradients of Biodiversity: Pattern, Process, Scale, and Synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Heikkinen, R.K. Predicting Patterns of Vascular Plant Species Richness with Composite Variables: A Meso-Scale Study in Finnish Lapland. Vegetatio 1996, 126, 151–165. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J. Abiotic and Biotic Determinants of Plant Diversity in Aquatic Communities Invaded by Water Hyacinth [Eichhornia crassipes (Mart.) Solms]. Front. Plant Sci. 2020, 11, 1306. [Google Scholar] [CrossRef] [PubMed]
- Alahuhta, J.; Antikainen, H.; Hjort, J.; Helm, A.; Heino, J. Current climate overrides historical effects on species richness and range size of freshwater plants in Europe and North America. J. Ecol. 2020, 108, 1262–1275. [Google Scholar] [CrossRef]
- Li, X.R. Characteristics of Temperature and Precipitation Change on the Tibet Plateau under the Background of Global Warming. Adv. Geosci. 2019, 9, 1042–1049. [Google Scholar] [CrossRef]
- Mao, L.; Chen, S.; Zhang, J.; Hou, Y.; Zhou, G.; Zhang, X. Vascular plant diversity on the roof of the world: Spatial patterns and environmental determinants. J. Syst. Evol. 2013, 51, 371–381. [Google Scholar] [CrossRef]
- Pandey, B.; Nepal, N.; Tripathi, S.; Pan, K.; Dakhil, M.A.; Timilsina, A.; Justine, M.F.; Koirala, S.; Nepali, K.B. Distribution Pattern of Gymnosperms’ Richness in Nepal: Effect of Environmental Constrains along Elevational Gradients. Plants 2020, 9, 625. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, W. Spatial–Temporal Characteristics of Precipitation and Its Relationship with Land Use/Cover Change on the Qinghai-Tibet Plateau, China. Land 2021, 10, 269. [Google Scholar] [CrossRef]
- Li, F.; Lu, H.; Wang, G.; Yao, Z.; Li, Q.; Qiu, J. Zoning of precipitation regimes on the Qinghai–Tibet Plateau and its surrounding areas responded by the vegetation distribution. Sci. Total Environ. 2022, 838, 155844. [Google Scholar] [CrossRef]
- Wang, R.; He, M.; Niu, Z. Responses of Alpine Wetlands to Climate Changes on the Qinghai-Tibetan Plateau Based on Remote Sensing. Chin. Geogr. Sci. 2020, 30, 189–201. [Google Scholar] [CrossRef]
- Vestergaard, O.; Sand-Jensen, K. Aquatic macrophyte richness in Danish lakes in relation to alkalinity, transparency, and lake area. Can. J. Fish. Aquat. Sci. 2000, 57, 2022–2031. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- Muellner Riehl, A.N.; Schnitzler, J.; Kissling, W.D.; Mosbrugger, V.; Rijsdijk, K.F.; Seijmonsbergen, A.C.; Versteegh, H.; Favre, A. Origins of global mountain plant biodiversity: Testing the ‘mountain-geobiodiversity hypothesis’. J. Biogeogr. 2019, 46, 2826–2838. [Google Scholar] [CrossRef] [Green Version]
- Chatanga, P.; Seleteng-Kose, L. Montane Palustrine Wetlands of Lesotho: Vegetation, Ecosystem Services, Current Status, Threats and Conservation. Wetlands 2021, 41, 67. [Google Scholar] [CrossRef]
- Bolpagni, R.; Piotti, A. The importance of being natural in a human-altered riverscape: Role of wetland type in supporting habitat heterogeneity and the functional diversity of vegetation. Aquatic Conserv. Mar. Freshw. Ecosyst. 2016, 26, 1168–1183. [Google Scholar] [CrossRef]
- Fleishman, E.; Noss, R.; Noon, B. Utility and limitations of species richness metrics for conservation planning. Ecol. Indic. 2006, 6, 543–553. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Baptista, L.R.D.M.; Lorscheitter, M.L.; Scherer, C. Floristic composition of a Subtropical bog, Eastern Plateau from southern Brazil. Check List 2012, 8, 224–236. [Google Scholar] [CrossRef]
- Tendar, P.; Sridith, K. Vegetation structure of wetlands in Eastern Himalayan Highlands of Gasa, Bhutan. Scienceasia 2021, 47, 78. [Google Scholar] [CrossRef]
- Pott, A.; Oliveira, A.K.M.; Damasceno-Junior, G.A.; Silva, J.S.V. Plant diversity of the Pantanal wetland. Braz. J. Biol. 2011, 71, 265–273. [Google Scholar] [CrossRef]
- Sieben, E.J.J.; Glen, R.P.; van Deventer, H.; Dayaram, A. The contribution of wetland flora to regional floristic diversity across a wide range of climatic conditions in southern Africa. Biodivers. Conserv. 2021, 30, 575–596. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | p-Value | R2 |
|---|---|---|
| Energy | ||
| AMT | p < 0.001 | 0.29 |
| TS | p < 0.001 | 0.15 |
| Water | ||
| AP | p < 0.001 | 0.39 |
| PDM | p < 0.001 | 0.24 |
| PS | p < 0.001 | 0.20 |
| Habitat | ||
| Ratio_WA | p < 0.05 | 0.02 |
| EVC | p < 0.001 | 0.14 |
| WA | p < 0.05 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhou, Y.; Liu, F.; Liu, X.; Wang, Q. Diversity Patterns of Wetland Angiosperms in the Qinghai-Tibet Plateau, China. Diversity 2022, 14, 777. https://doi.org/10.3390/d14100777
Li Y, Zhou Y, Liu F, Liu X, Wang Q. Diversity Patterns of Wetland Angiosperms in the Qinghai-Tibet Plateau, China. Diversity. 2022; 14(10):777. https://doi.org/10.3390/d14100777
Chicago/Turabian StyleLi, Yigang, Yadong Zhou, Fan Liu, Xing Liu, and Qingfeng Wang. 2022. "Diversity Patterns of Wetland Angiosperms in the Qinghai-Tibet Plateau, China" Diversity 14, no. 10: 777. https://doi.org/10.3390/d14100777