
Characterization of 15 Earthworm Mitogenomes from Northeast China and Its Phylogenetic Implication (Oligochaeta: Lumbricidae, Moniligastridae)

Abstract
:1. Introduction
2. Materials and Methods
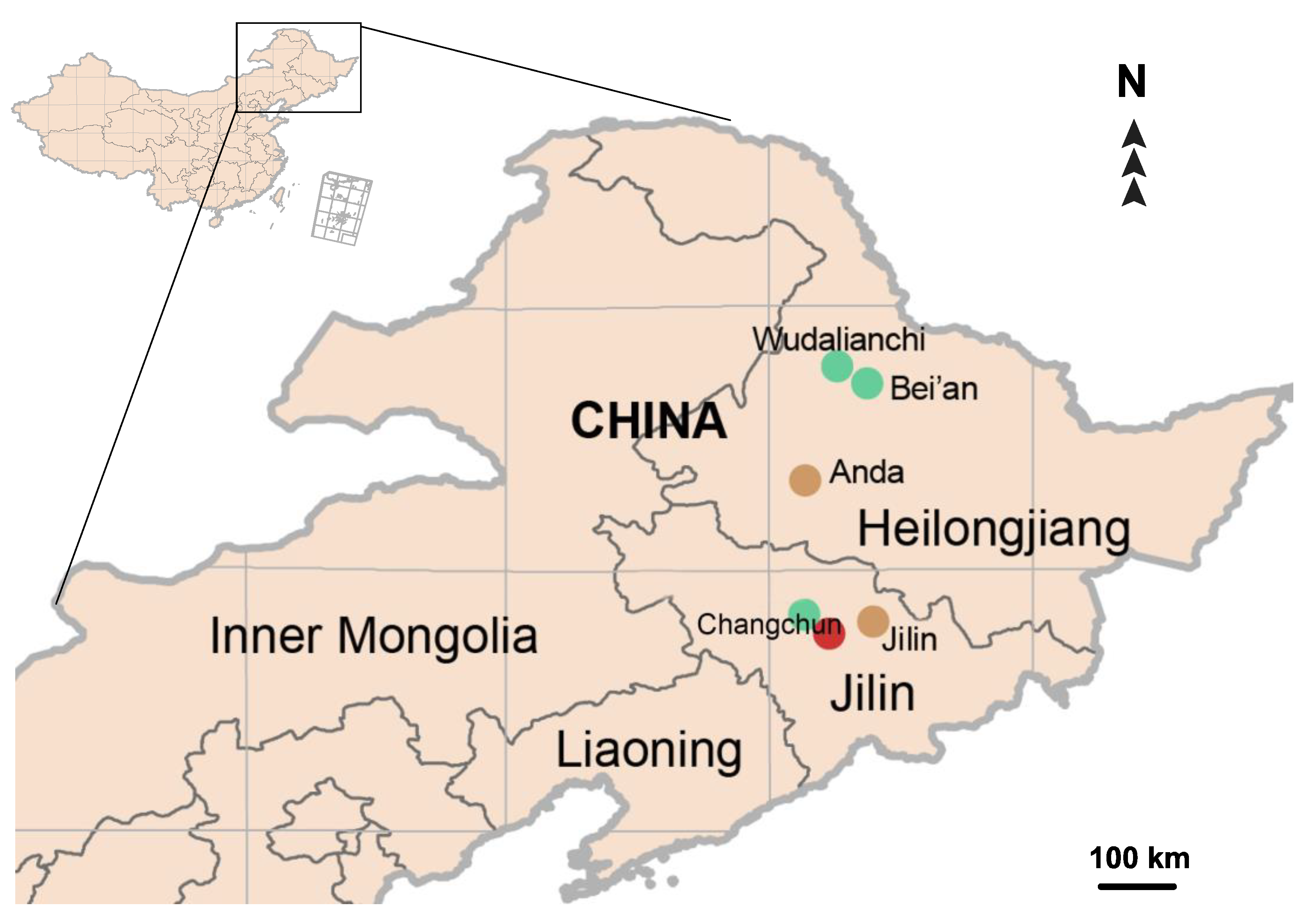
2.1. Sample Preparation and DNA Extraction
2.2. Next Generation Sequencing
2.3. Data Assembly and Mitogenome Annotation
2.4. Statistics of the Earthworm Mitogenomes
2.5. Phylogenetic Analysis
3. Results
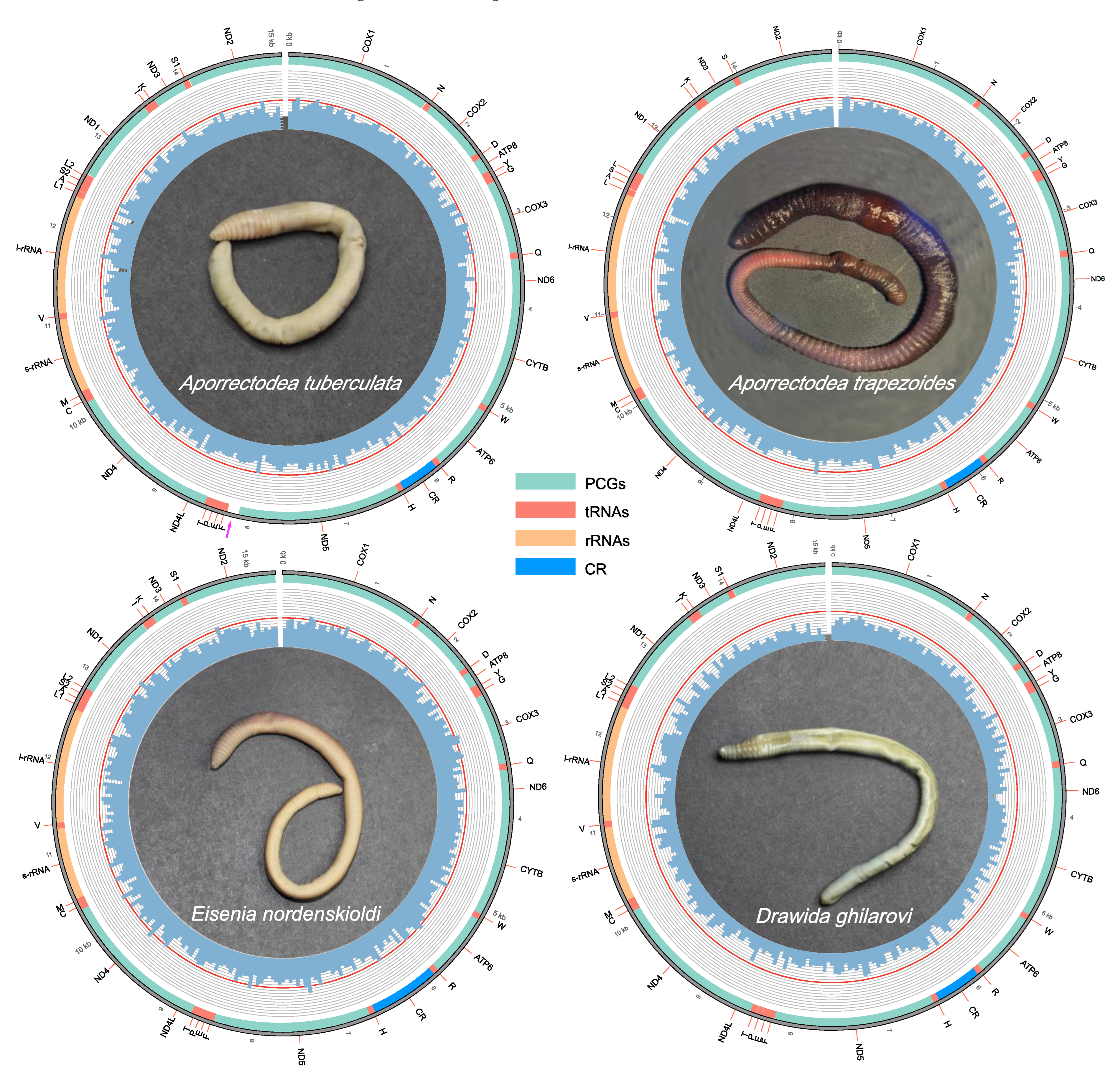
3.1. Mitogenome Organization and Nucleotide Composition of the Four Earthworm Species
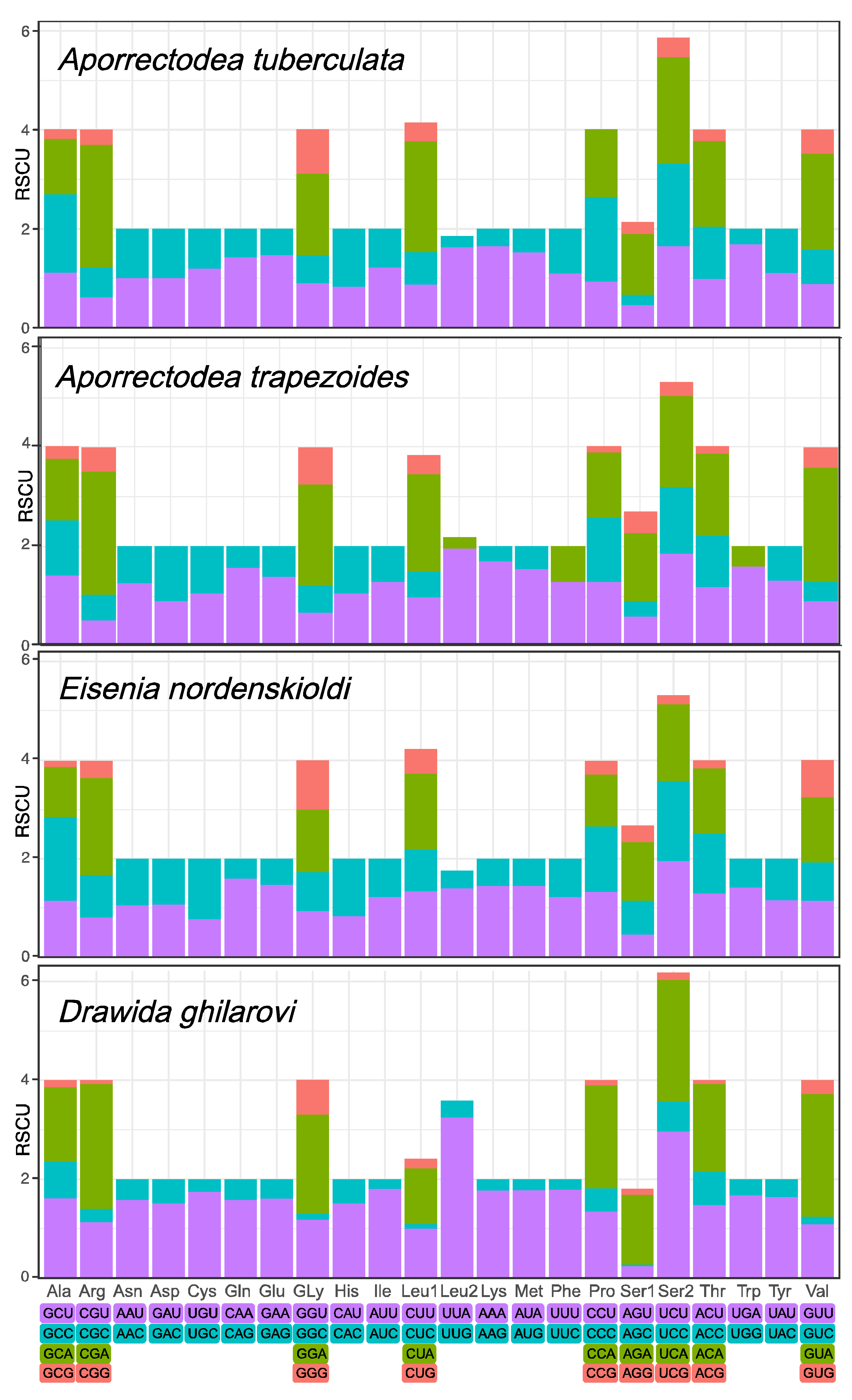
3.2. Protein-Coding Genes and Codon Usage in the Four Earthworm Species
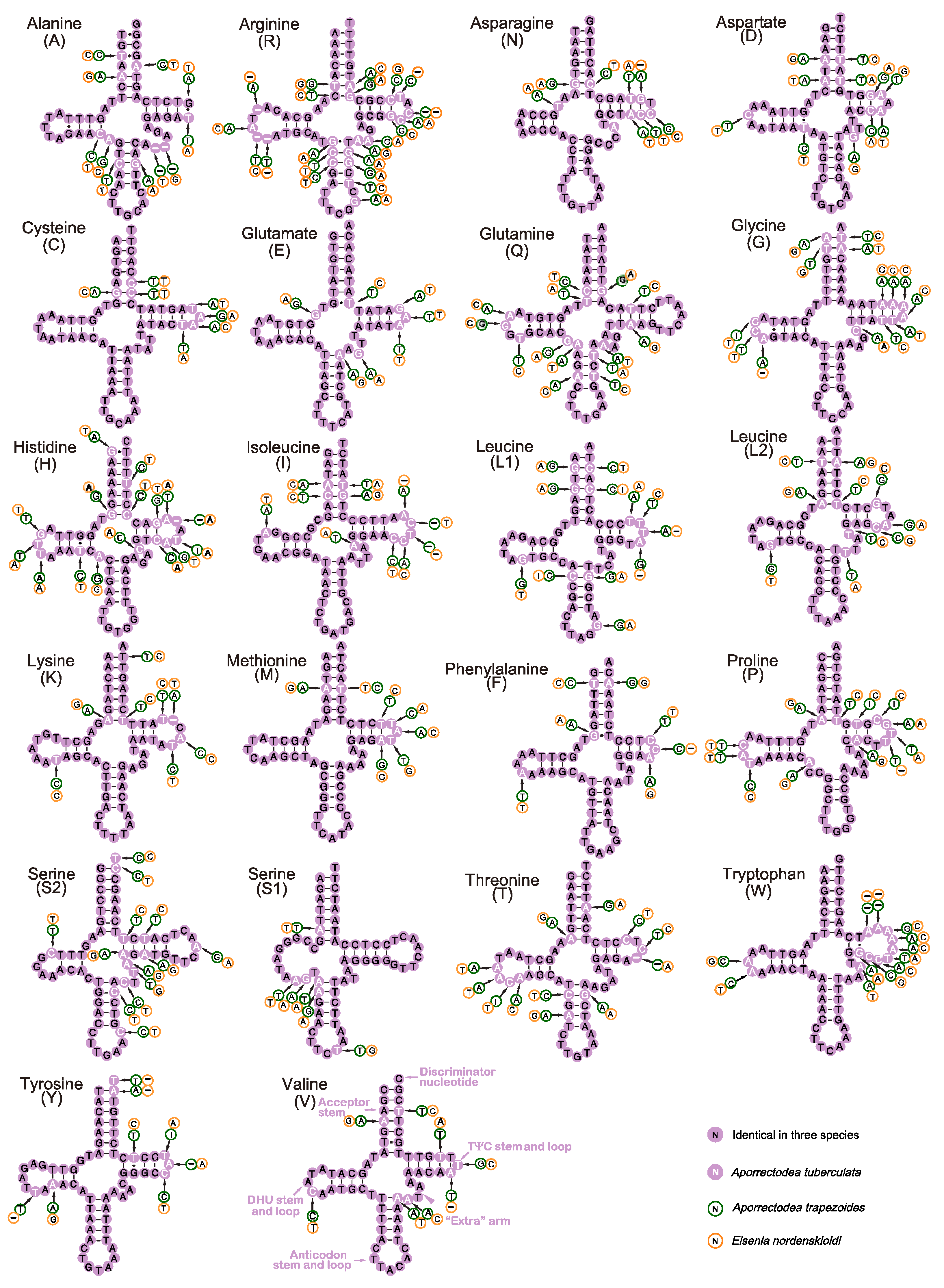
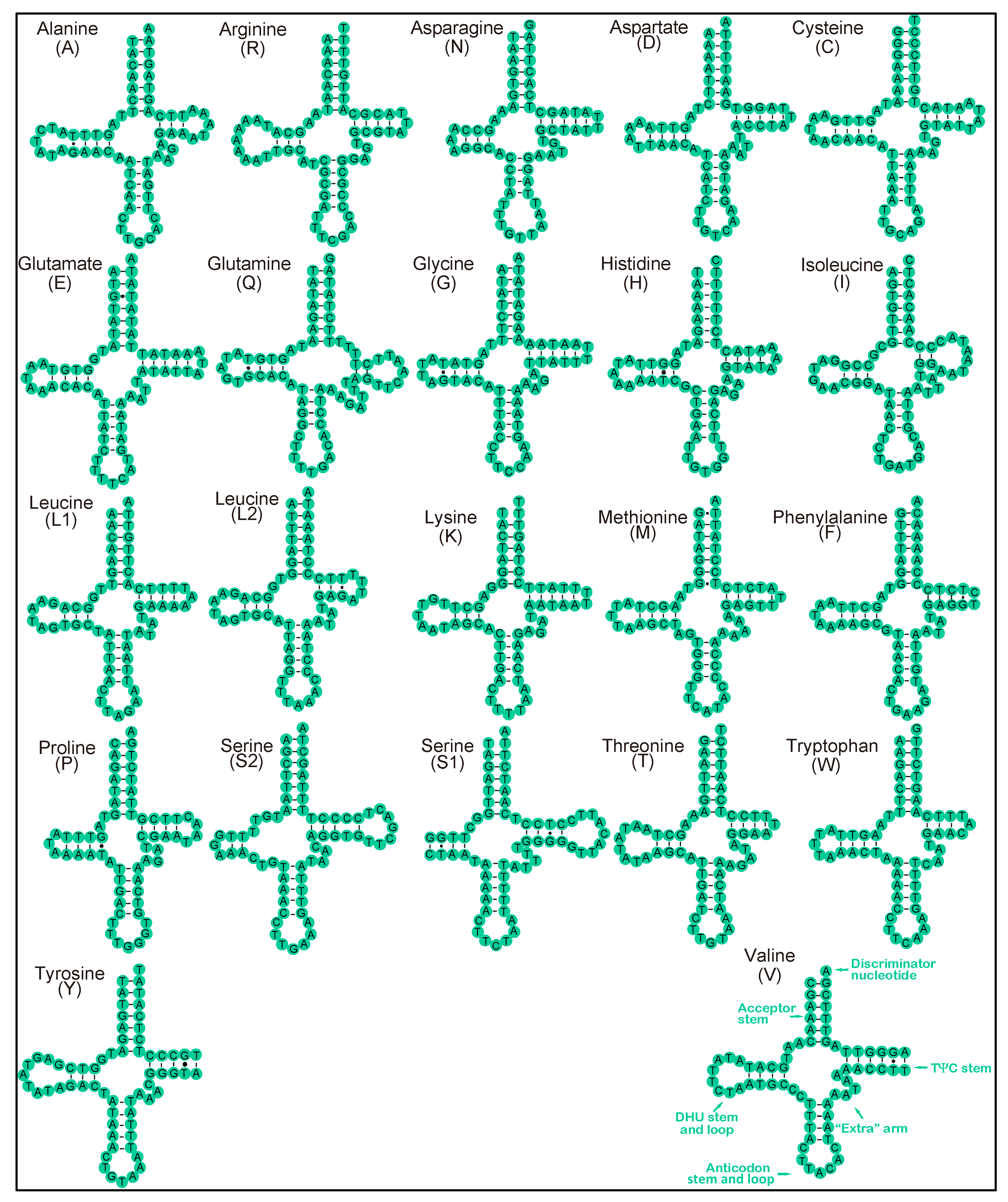
3.3. RNA Genes of the Four Earthworm Species
3.4. Putative Control Region and Non-Coding Region
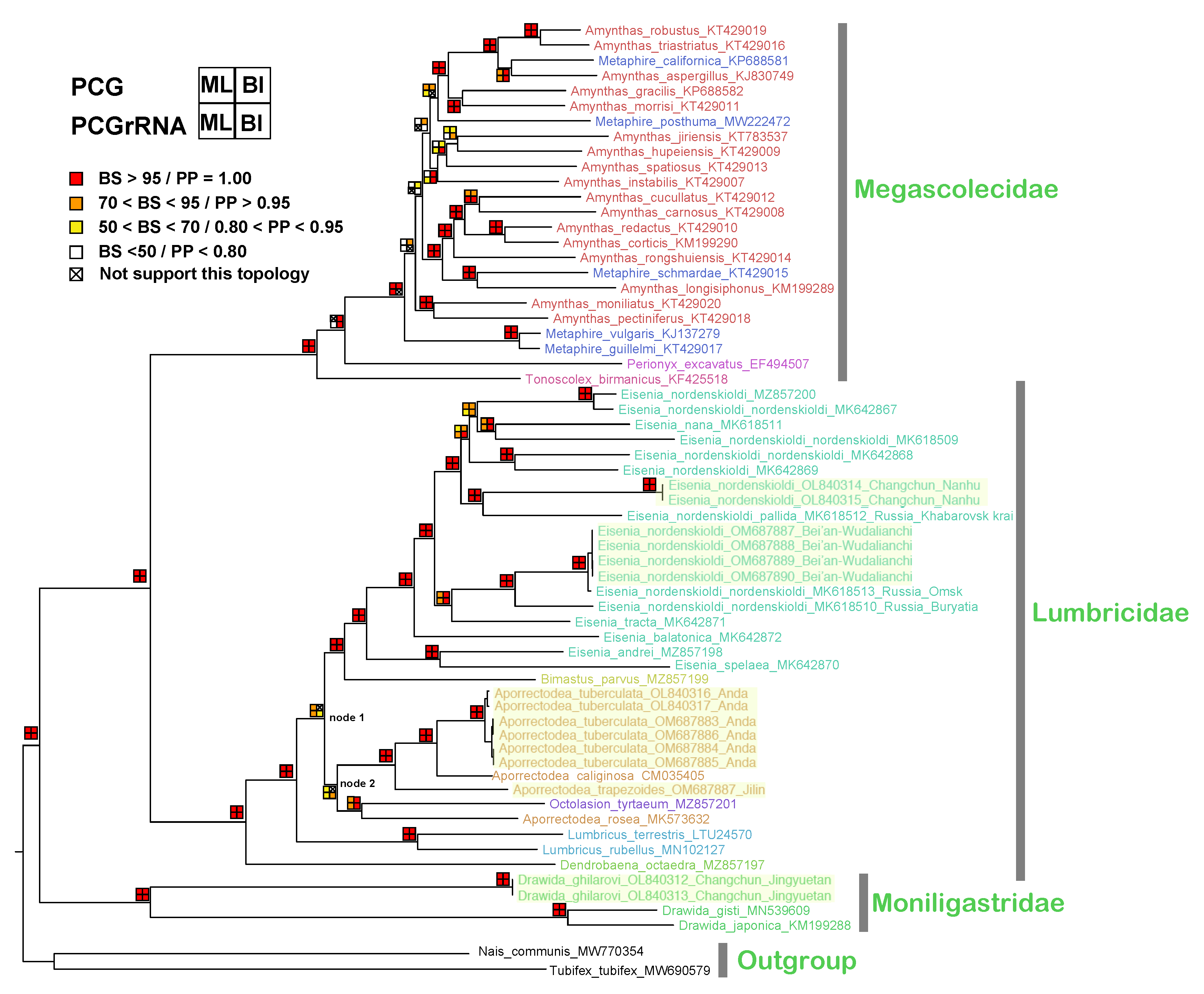
3.5. Phylogenetic Analysis
4. Discussion
4.1. Organization of a Partial Mitochondrial Genome
4.2. Genetic Diversity of Lumbricids in Northeast China
4.3. Phylogenetic Inference
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, C.A. Earthworm Ecology; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar] [CrossRef]
- Ganin, G.N.; Atopkin, D.M. Molecular differentiation of epigeic and anceic forms of Drawida ghilarovi Gates, 1969 (Moniligastridae, Clitellata) in the Russian Far East: Sequence data of two mitochondrial genes. Eur. J. Soil. Biol. 2018, 86, 1–7. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Bulakhova, N.A.; Vinokurov, N.N.; Peltek, S.E. Phylogeography of Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) from the north of Asia. Polar Biol. 2018, 41, 237–247. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Derzhinsky, Y.A.; Poluboyarova, T.V.; Golovanova, E.V.; Peltek, S.E. Phylogeography and genetic lineages of Aporrectodea rosea (Lumbricidae, Annelida). Eur. J. Soil. Biol. 2020, 99, 103191. [Google Scholar] [CrossRef]
- Pérez-Losada, M.; Ricoy, M.; Marshall, J.C.; Domínguez, J. Phylogenetic assessment of the earthworm Aporrectodea caliginosa species complex (Oligochaeta: Lumbricidae) based on mitochondrial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2009, 52, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Ganin, G.; Anisimov, A.; Roslik, G.; Atopkin, D. The Russian Far East Endemic Drawida Ghilarovi (Oligochaeta, Moniligastridae): Polimorphism, Ecology Specifics And Karyotype. Russ. J. Zool. 2014, 93, 1070–1079. [Google Scholar] [CrossRef]
- Atopkin, D.M.; Ganin, G.N. Genetic differentiation of black and grey colored forms of the earthworm Drawida ghilarovi Gates, 1969 (Moniligastridae, Oligochaeta) on Russian Far East. Eur. J. Soil. Biol. 2015, 67, 12–16. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Golovanova, E.V.; Ershov, N.I.; Poluboyarova, T.V.; Berman, D.I.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Phylogeny of the Eisenia nordenskioldi complex based on mitochondrial genomes. Eur. J. Soil. Biol. 2020, 96, 103137. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Golovanova, E.V.; Peltek, S.E. Cryptic diversity within the Nordenskiold’s earthworm, Eisenia nordenskioldi subsp. nordenskioldi (Lumbricidae, Annelida). Eur. J. Soil. Biol. 2013, 58, 13–18. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Peltek, S.E. Phylogeography of the earthworm Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) in northeastern Eurasia. Dokl. Biol. Sci. 2015, 461, 85–88. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Bazarova, N.E.; Bulakhova, N.A.; Porco, D.; Peltek, S.E. Cryptic genetic lineages in Eisenia nordenskioldi pallida (Oligochaeta, Lumbricidae). Eur. J. Soil. Biol. 2016, 75, 151–156. [Google Scholar] [CrossRef]
- Shekhovtsov, S.; Berman, D.; Golovanova, E.; Peltek, S. Genetic diversity of the earthworm Eisenia nordenskioldi (Lumbricidae, Annelida). Vavilov J. Genet. Breed. 2017, 21, 589–595. [Google Scholar] [CrossRef]
- Vsevolodova-Perel, T.S. The Earthworms of the Fauna of Russia; Nauka: Moscow, Russia, 1997. [Google Scholar]
- Ganin, G. Structural and Functional Organization of Mezopedobiont Communities of the Southern Russian Far East; Russian Academy of Sciences Far Eastern Banch, Institute of Water and Ecological Problems: Vladivostok, Russia, 2011; p. 380. [Google Scholar]
- Ganin, G.; Anisimov, A.; Roslik, G.; Atopkin, D. Earthworms Drawida ghilarovi Gates, 1969 (Oligochaeta, Moniligastridae): 1. Polimorphism, geographic range, ecology specifics. Amurian Zool. J. 2013, 4, 401–404. [Google Scholar] [CrossRef]
- Boore, J.L.; Brown, W.M. Complete sequence of the mitochondrial DNA of the annelid worm Lumbricus terrestris. Genetics 1995, 141, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.R.; Hong, Y.; Win, T.M.; Kim, I. Complete mitochondrial genome of the Burmese giant earthworm, Tonoscolex birmanicus (Clitellata: Megascolecidae). Mitochondrial DNA 2015, 26, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiang, J.; Dong, Y.; Qiu, J. Complete mitochondrial genome of four pheretimoid earthworms (Clitellata: Oligochaeta) and their phylogenetic reconstruction. Gene 2015, 574, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiang, J.; Dong, Y.; Qiu, J. Complete mitochondrial genome of an Amynthas earthworm, Amynthas aspergillus (Oligochaeta: Megascolecidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 1876–1877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiang, J.; Dong, Y.; Qiu, J. Complete mitochondrial genome of a Pheretimoid earthworm Metaphire vulgaris (Oligochaeta: Megascolecidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2014, 27, 297–298. [Google Scholar] [CrossRef]
- Zhang, L.; Sechi, P.; Yuan, M.; Jiang, J.; Dong, Y.; Qiu, J. Fifteen new earthworm mitogenomes shed new light on phylogeny within the Pheretima complex. Sci. Rep. 2016, 6, 20096. [Google Scholar] [CrossRef]
- Conrado, A.C.; Arruda, H.; Stanton, D.W.G.; James, S.W.; Peter, K.; Brown, G.; Silva, E.; Dupont, L.; Taheri, S.; Morgan, A.J.; et al. The complete mitochondrial DNA sequence of the pantropical earthworm Pontoscolex corethrurus (Rhinodrilidae, Clitellata): Mitogenome characterization and phylogenetic positioning. ZooKeys 2017, 688, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, H.; Zhang, Y.; Ruan, H. The complete mitochondrial genome of Lumbricus rubellus (Oligochaeta, Lumbricidae) and its phylogenetic analysis. Mitochondrial DNA B Resour. 2019, 4, 2677–2678. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Peltek, S.E. The complete mitochondrial genome of Aporrectodea rosea (Annelida: Lumbricidae). Mitochondrial DNA B Resour. 2019, 4, 1752–1753. [Google Scholar] [CrossRef]
- Liu, H.; Xu, N.; Zhang, Q.; Wang, G.; Xu, H.; Ruan, H. Characterization of the complete mitochondrial genome of Drawida gisti (Metagynophora, Moniligastridae) and comparison with other Metagynophora species. Genomics 2020, 112, 3056–3064. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Zhao, H.; Yu, F.; Zhang, A.; Li, H. The First Mitogenomes of the Subfamily Odontiinae (Lepidoptera, Crambidae) and Phylogenetic Analysis of Pyraloidea. Insects 2021, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-García, M.; Pinilla-Beltrán, D.; Murillo-García, O.E.; Pinto, C.M.; Brito, J.; Shostell, J.M. Comparative mitogenome phylogeography of two anteater genera (Tamandua and Myrmecophaga; Myrmecophagidae, Xenarthra): Evidence of discrepant evolutionary traits. Zool. Res. 2021, 42, 525–547. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Hasegawa, H.; Cooley, J.R.; Simon, C.; Yoshimura, J.; Cai, W.; Sota, T.; Li, H. Mitochondrial Genomics Reveals Shared Phylogeographic Patterns and Demographic History among Three Periodical Cicada Species Groups. Mol. Biol. Evol. 2019, 36, 1187–1200. [Google Scholar] [CrossRef]
- Hou, Q.; Tu, F.; Liu, Y.; Liu, S. Characterization of the mitogenome of Uropsilus gracilis and species delimitation. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 1836–1837. [Google Scholar] [CrossRef]
- Sun, X. Divergence across the mitogenomes of Branchinella kugenumaensis (Anostraca: Thamnocephalidae) with implications for species delimitation. Mitochondrial DNA B Resour. 2021, 6, 631–633. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, Y.; Wang, Z.; Chen, H.; Qin, Y. Two Complete Mitogenomes of Chalcididae (Hymenoptera: Chalcidoidea): Genome Description and Phylogenetic Implications. Insects 2021, 12, 1049. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, H.; Li, Y. A report of the complete mitochondrial genome of Bisetocreagris titanium (Arachnida: Pseudoscorpiones: Neobisiidae) from Yunnan Province, China. Mitochondrial DNA Part B Resour. 2021, 6, 3212–3213. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Ganin, G.N.; Atopkin, D.M.; Wu, D.H. Earthworm Drawida (Moniligastridae) Molecular phylogeny and diversity in Far East Russia and Northeast China. Eur. Zool. J. 2020, 87, 180–191. [Google Scholar] [CrossRef]
- Saccone, C.; De Giorgi, C.; Gissi, C.; Pesole, G.; Reyes, A. Evolutionary genomics in Metazoa: The mitochondrial DNA as a model system. Gene 1999, 238, 195–209. [Google Scholar] [CrossRef]
- Ramesh, A.; Small, S.T.; Kloos, Z.A.; Kazura, J.W.; Nutman, T.B.; Serre, D.; Zimmerman, P.A. The complete mitochondrial genome sequence of the filarial nematode Wuchereria bancrofti from three geographic isolates provides evidence of complex demographic history. Mol. Biochem. Parasitol. 2012, 183, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Kern, E.M.A.; Kim, T.; Park, J.-K. The Mitochondrial Genome in Nematode Phylogenetics. Front. Ecol. Evol. 2020, 8, 250. [Google Scholar] [CrossRef]
- Yu, L.; Li, Y.-W.; Ryder, O.A.; Zhang, Y.-P. Analysis of complete mitochondrial genome sequences increases phylogenetic resolution of bears (Ursidae), a mammalian family that experienced rapid speciation. BMC Evol. Biol. 2007, 7, 198. [Google Scholar] [CrossRef] [PubMed]
- Miya, M.; Nishida, M. The mitogenomic contributions to molecular phylogenetics and evolution of fishes: A 15-year retrospect. Ichthyol. Res. 2015, 62, 29–71. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Xu, W.; Fang, Y.; Ruan, H. Characterization of Five New Earthworm Mitogenomes (Oligochaeta: Lumbricidae): Mitochondrial Phylogeny of Lumbricidae. Diversity 2021, 13, 580. [Google Scholar] [CrossRef]
- Hong, Y.; Kim, M.J.; Wang, A.R.; Kim, I. Complete mitochondrial genome of the earthworm, Amynthas jiriensis (Clitellata: Megascolecidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2017, 28, 163–164. [Google Scholar] [CrossRef]
- Yu, X.; Yang, H.; Liu, J.; Qi, Y.; Sun, L.; Tian, X. A strategy for a high enrichment of insect mitochondrial DNA for mitogenomic analysis. Gene 2022, 808, 145986. [Google Scholar] [CrossRef]
- Kim, D.W.; Lee, K.S.; Jee, S.H.; Seo, S.B.; Park, S.C.; Choo, J.K. Complete sequence analysis of the mitochondrial genome in the earthworm, Perionyx excavatus. Integr. Biosci. 2005, 9, A705. [Google Scholar]
- Lee, J.; Jung, J. First record of the complete mitochondrial genome of Tubifex tubifex (Müller) 1774 (Annelida; Clitellata; Oligochaeta) and phylogenetic analysis. Mitochondrial DNA B Resour. 2022, 7, 1208–1210. [Google Scholar] [CrossRef]
- Lee, J.; Jung, J. The complete mitochondrial genome of Nais communis Piguet, 1906 (Annelida; Clitellata; Naididae). Mitochondrial DNA B Resour. 2022, 7, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Yu, W.; Yang, J.; Song, Y.; dePamphilis, C.W.; Yi, T.; Li, D. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved annotation of protein-coding genes boundaries in metazoan mitochondrial genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 53942. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Figtree 1.4.0. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 July 2022).
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Lavrov, D.V.; Brown, W.M.; Boore, J.L. A novel type of RNA editing occurs in the mitochondrial tRNAs of the centipede Lithobius forficatus. Proc. Natl. Acad. Sci. USA 2000, 97, 13738–13742. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The Third Revolution in Sequencing Technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef]
- Hu, F.; Piao, Y.; Wang, H.; Pan, Y.; Fu, Y.; Song, W. The Investigation of the Usage of Herbicide in Heilongjiang Province. J. Agric. 2015, 5, 25–31. [Google Scholar]
- Pop, A.A.; Csuzdi, C.; Wink, M.; Pop, V.V. An Attempt to Reconstruct the Molecular Phylogeny of the Genus Allolobophora (sensu lato, Pop, 1941) Using 16S rDNA and COI Sequences (Oligochaeta, Lumbricidae); University Press: Cluj-Napoca, Romania, 2006. [Google Scholar]
- Shekhovtsov, S.; Ermolov, S.; Poluboyarova, T.; Kim-Kashmenskaya, M.; Derzhinsky, Y.; Peltek, S. Morphological differences between genetic lineages of the peregrine earthworm: Aporrectodea caliginosa (Savigny, 1826). Acta Zool. Acad. Sci. Hung. 2021, 67, 235–246. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Breinholt, J.W.; Stojanovic, M.; James, S.W.; Pérez-Losada, M. Underground evolution: New roots for the old tree of lumbricid earthworms. Mol. Phylogenet. Evol. 2015, 83, 7–19. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Taxa | Accession Number | Body Color | Habitat | References |
|---|---|---|---|---|---|
| Lumbricidae | Aporrectodeacaliginosa | CM035405 | - | - | Unpublished |
| Aporrectodea tuberculata | OL840316–7, OM687883–6 | Light grey | Forest | This study | |
| Aporrectodea trapezoides | OM687882 | Dark brown | Farmland | This study | |
| Aporrectodea rosea | MK573632 | - | - | [24] | |
| Bimastus parvus | MZ857199 | - | - | [39] | |
| Dendrobaena octaedra | MZ857197 | - | - | [39] | |
| Eisenia andrei | MZ857198 | - | - | [39] | |
| Eisenia balatonica | MK642872 | - | - | [8] | |
| Eisenia nana | MK618511 | - | - | [8] | |
| Eisenia nordenskioldi | OL840314–5, OM687887–90 | Light reddish | Forest | This study | |
| Eisenia nordenskioldi | MK642869 | - | - | [8] | |
| Eisenia nordenskioldi | MZ857200 | - | - | [39] | |
| Eisenia nordenskioldi nordenskioldi | MK618509 | - | - | [8] | |
| Eisenia nordenskioldi nordenskioldi | MK618510 | - | - | [8] | |
| Eisenia nordenskioldi nordenskioldi | MK618513 | - | - | [8] | |
| Eisenia nordenskioldi nordenskioldi | MK642867 | - | - | [8] | |
| Eisenia nordenskioldi nordenskioldi | MK642868 | - | - | [8] | |
| Eisenia nordenskioldi pallida | MK618512 | - | - | [8] | |
| Eisenia spelaea | MK642870 | - | - | [8] | |
| Eisenia tracta | MK642871 | - | - | [8] | |
| Lumbricus rubellus | MN102127 | Pink | Forest | [23] | |
| Lumbricus terrestris | LTU24570 | - | - | [16] | |
| Octolasion tyrtaeum | MZ857201 | - | - | [39] | |
| Megascolecidae | Amynthas aspergillus | KJ830749 | - | - | [19] |
| Amynthas carnosus | KT429008 | - | - | [21] | |
| Amynthas corticis | KM199290 | - | - | [18] | |
| Amynthas cucullatus | KT429012 | - | - | [21] | |
| Amynthas gracilis | KP688582 | - | - | [18] | |
| Amynthas hupeiensis | KT429009 | - | - | [21] | |
| Amynthas jiriensis | KT783537 | - | - | [40] | |
| Amynthas longisiphonus | KM199289 | - | - | [18] | |
| Amynthas moniliatus | KT429020 | - | - | [21] | |
| Amynthas morrisi | KT429011 | - | - | [21] | |
| Amynthas pectiniferus | KT429018 | - | - | [21] | |
| Amynthas robustus | KT429019 | - | - | [21] | |
| Amynthas redactus | KT429010 | - | - | [21] | |
| Amynthas instabilis | KT429007 | - | - | [21] | |
| Amynthas rongshuiensis | KT429014 | - | - | [21] | |
| Amynthas spatiosus | KT429013 | - | - | [21] | |
| Amynthas triastriatus | KT429016 | - | - | [21] | |
| Metaphire schmardae | KT429015 | - | - | [21] | |
| Metaphire californica | KP688581 | - | - | [18] | |
| Metaphire guillelmi | KT429017 | - | - | [21] | |
| Metaphire posthuma | MW222472 | - | - | [41] | |
| Metaphire vulgaris | KJ137279 | - | - | [20] | |
| Perionyx excavatus | EF494507 | - | - | [42] | |
| Tonoscolex birmanicus | KF425518 | - | - | [17] | |
| Moniligastridae | Drawida ghilarovi | OL840312–3 | Gray | Forest | This study |
| Drawida gisti | MN539609 | - | - | [25] | |
| Drawida japonica | KM199288 | - | - | [21] | |
| Naididae Outgroup | Tubifex tubifex | MW690579 | - | - | [43] |
| Nais communis | MW770354 | - | - | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Fan, S.; Aspe, N.M.; Feng, L.; Zhang, Y. Characterization of 15 Earthworm Mitogenomes from Northeast China and Its Phylogenetic Implication (Oligochaeta: Lumbricidae, Moniligastridae). Diversity 2022, 14, 714. https://doi.org/10.3390/d14090714
Zhao H, Fan S, Aspe NM, Feng L, Zhang Y. Characterization of 15 Earthworm Mitogenomes from Northeast China and Its Phylogenetic Implication (Oligochaeta: Lumbricidae, Moniligastridae). Diversity. 2022; 14(9):714. https://doi.org/10.3390/d14090714
Chicago/Turabian StyleZhao, Huifeng, Shuanghu Fan, Nonillon M. Aspe, Lichao Feng, and Yufeng Zhang. 2022. "Characterization of 15 Earthworm Mitogenomes from Northeast China and Its Phylogenetic Implication (Oligochaeta: Lumbricidae, Moniligastridae)" Diversity 14, no. 9: 714. https://doi.org/10.3390/d14090714