
Anthropogenic Pressure and Climate Change Could Severely Hamper the Avian Metacommunity of the Sicilian Wetlands


Abstract
:1. Introduction
2. Materials and Methods
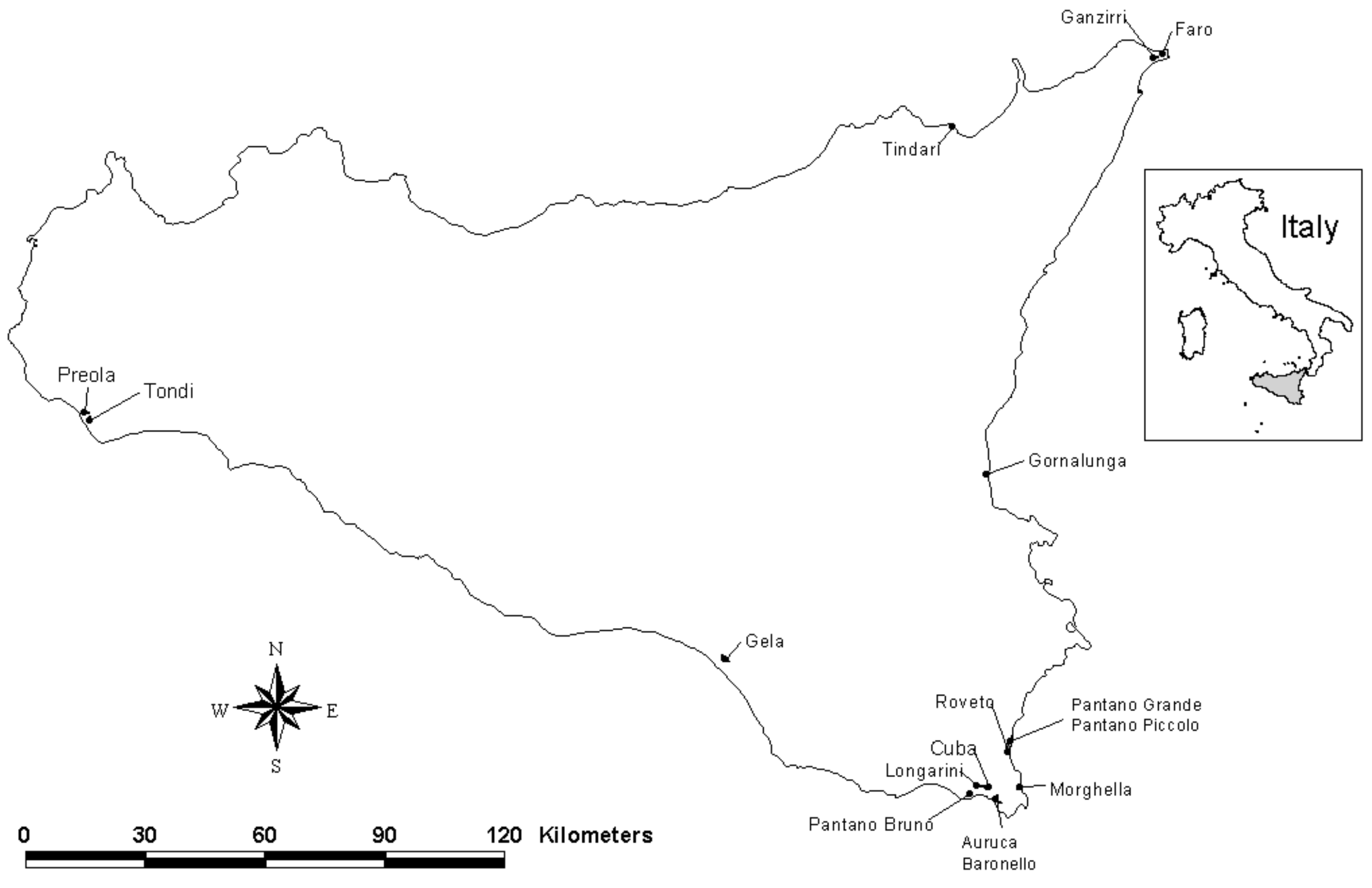
2.1. Field Surveys
2.2. Wetland Attributes
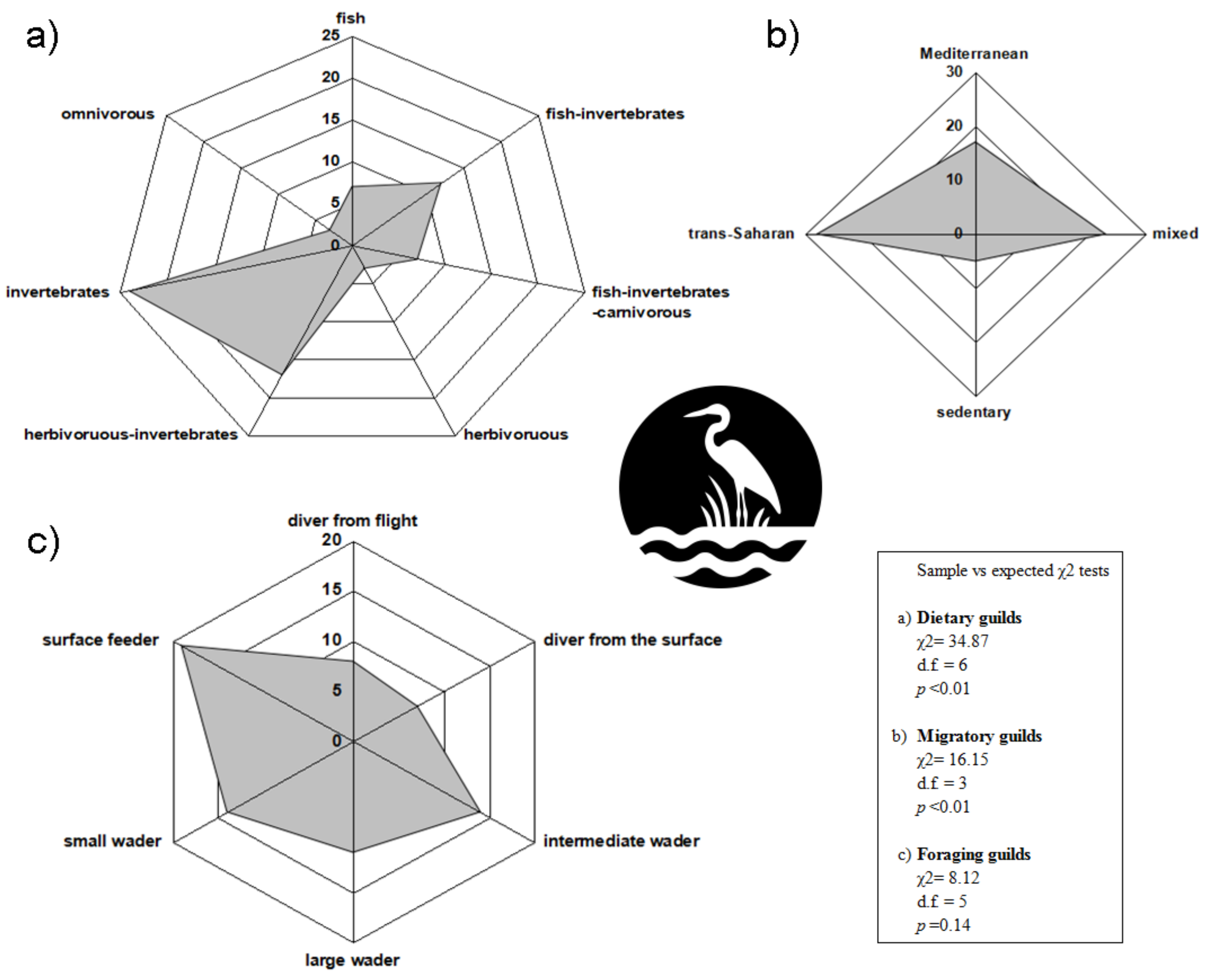
2.3. Species Functional Traits
2.4. Avian Diversity
2.5. Metacommunity Structure
2.5.1. Metacommunity Modularity
2.5.2. Metacommunity Nestedness
2.5.3. Species Assemblage
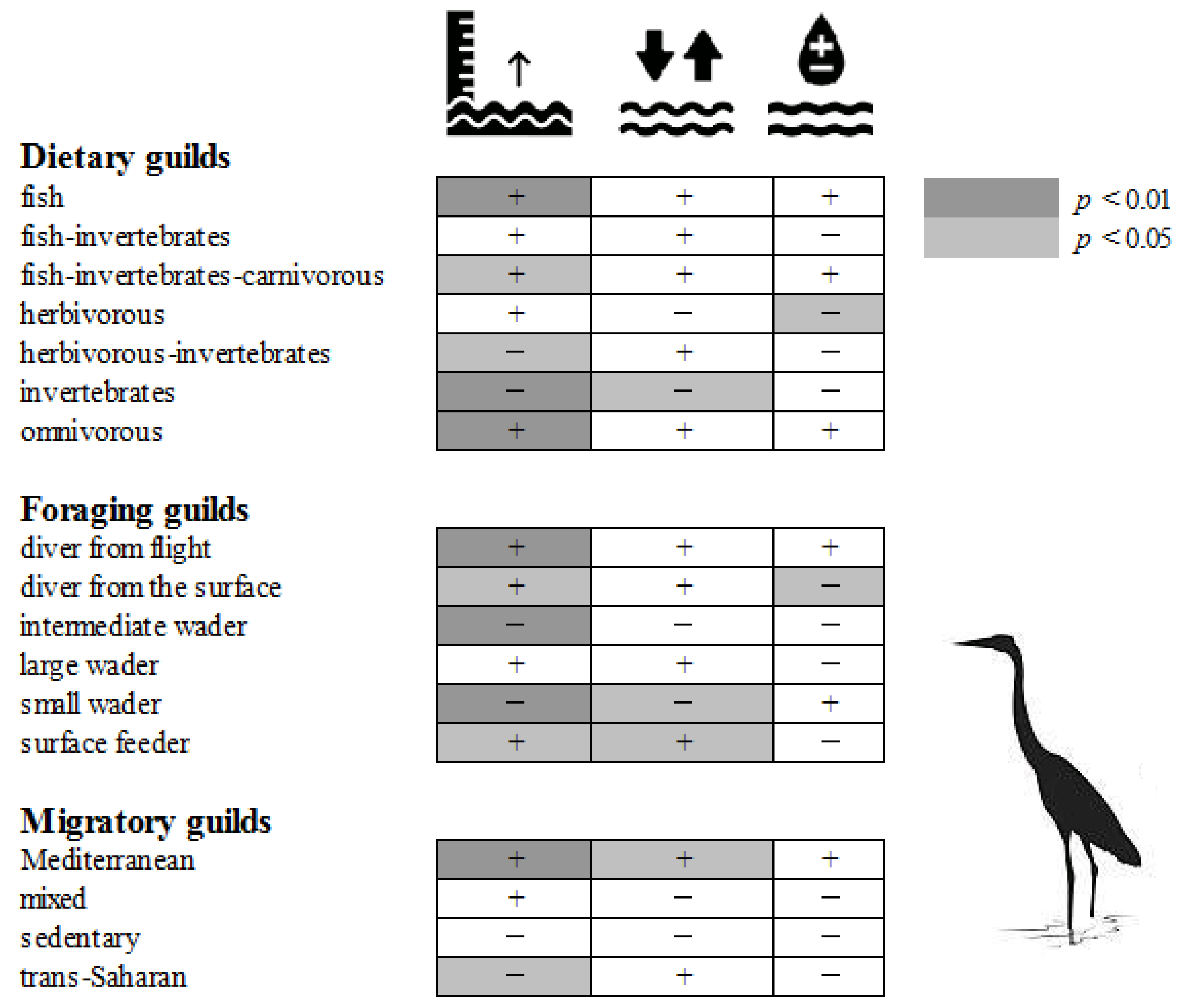
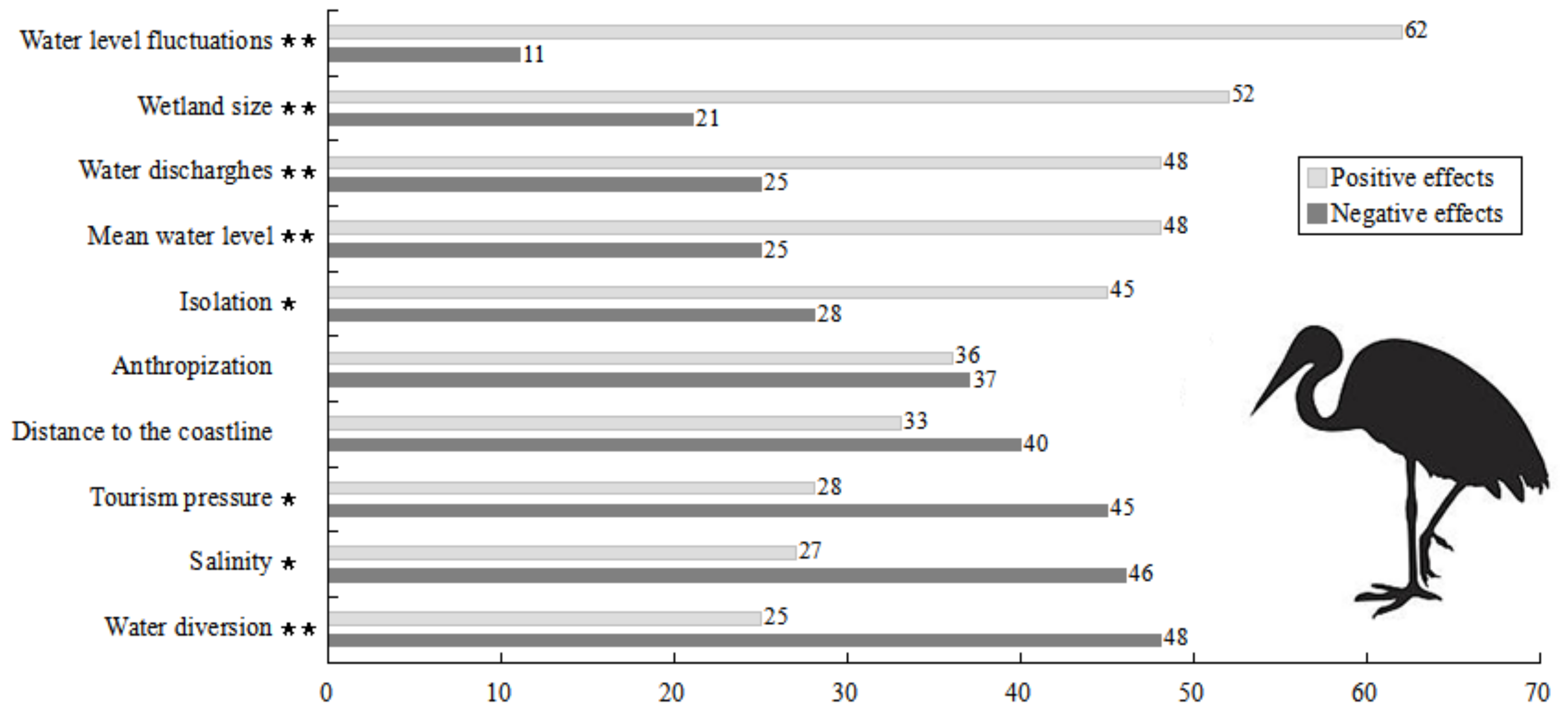
2.6. Effects of Wetland Attributes on the Avian Metacommunity
2.6.1. Effects on Species Richness
2.6.2. Effects on Species Guilds
2.6.3. Effects on Single Species
3. Results
4. Discussion
Implications for the Conservation of the Avian Metacommunity of the Sicilian Wetlands
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elphick, J. The Atlas of Bird Migration: Tracing the Great Journeys of the World’s Birds; Natural History Museum: London, UK, 2007. [Google Scholar]
- Runge, C.A.; Watson, J.E.M.; Butchart, S.H.M.; Hanson, J.O.; Possingham, H.P.; Fuller, R.A. Protected areas and global conservation of migratory birds. Science 2015, 350, 1255–1258. [Google Scholar] [CrossRef] [PubMed]
- Alerstam, T. Bird Migration; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Boere, G.C.; Galbraith, C.A.; Stroud, D.A. (Eds.) Waterbirds around the World; The Stationery Office: Edinburgh, UK, 2006. [Google Scholar]
- Barnard, J.; Bennun, L.; Iyer, S.; Keil, F.; Ponziani, C. Wings over Wetlands (WOW)—Flyway Conservation at Work across Africa and Eurasia. Enhancing the Conservation of the Critical Network of Sites Required by Migratory Waterbirds on the African–Eurasian Flyways; UNEP-GEF: Geneva, Switzerland, 2010. [Google Scholar]
- Gardner, R.C.; Finlayson, C.M. Global Wetland Outlook: State of the World’s Wetlands and Their Services to People; Ramsar Convention: Gland, Switzerland, 2018. [Google Scholar]
- Gardner, R.C.; Barchiesi, S.; Beltrame, C.; Finlayson, C.M.; Galewski, T.; Harrison, I.J.; Paganini, M.; Perennou, C.; Rosenqvist, A.; Walpole, M.; et al. State of the World’s Wetlands and Their Services to People: A Compilation of Recent Analyses; Social Science Electronic Publishing: Gland, Switzerland, 2015. [Google Scholar]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; John Wiley & Sons: New York, NY, USA, 2015. [Google Scholar]
- Dodman, T.; Boere, G.C. The Flyway Approach to the Conservation and Wise Use of Waterbirds and Wetlands: The Flyway Training Kit; Wetlands International and BirdLife International: Ede, The Netherlands, 2010. [Google Scholar]
- Berthold, P. Bird Migration: A General Survey; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Spina, F.; Volponi, S. Atlante della Migrazione Degli Uccelli in Italia; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale: Rome, Italy, 2008. [Google Scholar]
- Bijlsma, R.G. Bottleneck Areas for Migratory Birds in the Mediterranean Region: An Assessment of the Problems and Recommendations for Action; International Council for Bird Preservation: Cambridge, UK, 1990. [Google Scholar]
- Heath, M.F.; Evans, M.I. (Eds.) Important Bird Areas in Europe. Priority Sites for Conservation. 2 Vols; BirdLife International: Cambridge, UK, 2000. [Google Scholar]
- Ferrarini, A.; Gustin, M.; Celada, C. Twenty-Three Years of Land-Use Changes Induced Considerable Threats to the Main Wetlands of Sardinia and Sicily (Italy) along the Mediterranean Bird Flyways. Diversity 2021, 13, 240. [Google Scholar] [CrossRef]
- Ferrarini, A.; Celada, C.; Gustin, M. Preserving the Mediterranean bird flyways: Assessment and prioritization of 38 main wetlands under human and climate threats in Sardinia and Sicily (Italy). Sci. Total Environ. 2020, 751, 141556. [Google Scholar] [CrossRef] [PubMed]
- De Pietro, R.; Ientile, R.; Puccia, S.; Sabella, G. Birds of Gelsari and Lentini marshes, special protection area for the protection and maintenance of aquatic avifauna in central Mediterranean. Ocean Coast. Manag. 2019, 169, 96–103. [Google Scholar] [CrossRef]
- Griffith, D.A. Spatial Autocorrelation: A Primer; Association of American Geographers: Washington, DC, USA, 1987. [Google Scholar]
- Hutto, R.L.; Pletschet, S.M.; Hendricks, P. A fixed-radius point count method for nonbreeding and breeding season use. Auk 1986, 103, 593–602. [Google Scholar] [CrossRef]
- Cramp, S.E.; Simmons, K.E.L.; Brooks, D.J.; Perrins, C.M. Handbook of the Birds of Europe, the Middle East and North Africa: The Birds of the Western Palearctic; Oxford University Press: Oxford, UK, 1977–1994; Volume 1–9. [Google Scholar]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Leibold, M.A.; Mikkelson, G.M. Coherence, species turnover, and boundary clumping: Elements of meta-community structure. Oikos 2002, 97, 237–250. [Google Scholar] [CrossRef]
- Newman, M.E.J.; Girvan, M. Finding and evaluating community structure in networks. Phys. Rev. 2004, 69, 026113. [Google Scholar] [CrossRef]
- Erdős, P.; Rényi, A. On random graphs. Publ. Math. Debr. 1959, 6, 290–297. [Google Scholar]
- Bascompte, J.; Jordano, P.; Melian, C.J.; Olesen, J.M. The nested assembly of plant–animal mutualistic networks. Proc. Natl. Acad. Sci. USA 2003, 100, 9383–9387. [Google Scholar] [CrossRef]
- Rodríguez-Gironés, M.A.; Santamaría, L. A new algorithm to calculate the nestedness temperature of presence-absence matrices. J. Biogeogr. 2006, 33, 924–935. [Google Scholar] [CrossRef]
- Atmar, W.; Patterson, B.D. The measure of order and disorder in the distribution of species in fragmented habitat. Oecologia 1993, 96, 373–382. [Google Scholar] [CrossRef]
- Stone, L.; Roberts, A. The checkerboard score and species distributions. Oecologia 1990, 85, 74–79. [Google Scholar] [CrossRef]
- Diamond, J.M. Assembly of species communities. In Ecology and Evolution of Communities; Cody, M., Diamond, J.M., Eds.; Harvard University Press: Cambridge, UK, 1975; pp. 342–444. [Google Scholar]
- Gotelli, N.J. Null model analysis of species co-occurrence patterns. Ecology 2000, 81, 2606–2621. [Google Scholar] [CrossRef]
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating behavior to habitat: Solutions to the fourth-corner problem. Ecology 1997, 78, 547–562. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Ulrich, W.; Maestre, F.T. Randomization tests for quantifying species importance to ecosystem function. Methods Ecol. Evol. 2011, 2, 634–642. [Google Scholar] [CrossRef]
- Thébault, E. Identifying compartments in presence–absence matrices and bipartite networks: Insights into modularity measures. J. Biogeogr. 2013, 40, 759–768. [Google Scholar] [CrossRef]
- Krishna, A.; Guimaraes, P.R., Jr.; Jordano, P.; Bascompte, J. A neutral-niche theory of nestedness in mutualistic networks. Oikos 2008, 117, 1609–1618. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.-L. Nestedness of insular avifaunas: Simple summary statistics masking complex species patterns. Ornis Fenn. 1991, 68, 178–192. [Google Scholar]
- Wright, D.H.; Reeves, J.H. On the meaning and measurement of nestedness of species assemblages. Oecologia 1992, 92, 416–428. [Google Scholar] [CrossRef]
- Patterson, B.D.; Atmar, W. Nested subsets and the structure of insular mammalian faunas and archipelagos. Biol. J. Linn. Soc. Lond. 1986, 28, 6582. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P. Ecological Assembly Rules: Perspectives, Advances, Retreats; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Kessler, M.; Abrahamczyk, S.; Bos, M.; Buchori, D.; Putra, D.D.; Gradstein, S.R.; Höhn, P.; Kluge, J.; Orend, F.; Pitopang, R.; et al. Alpha and beta diversity of plants and animals along a tropical land-use gradient. Ecol. Appl. 2009, 19, 2142–2156. [Google Scholar] [CrossRef]
- Poysa, H. Resource utilization pattern and guild structure in a waterfowl community. Oikos 1983, 40, 295–307. [Google Scholar] [CrossRef]
- Ntiamoa-Baidu, Y.; Piersma, T.; Wiersma, P.; Poot, M.; Battley, P.; Gordon, C. Water depth selection, daily feeding routines and diets of waterbirds in coastal lagoons in Ghana. Ibis 1998, 140, 89–103. [Google Scholar] [CrossRef]
- Kirby, J.; Davidson, N.; Giles, N.; Owen, M.; Spray, C. Waterbirds and Wetland Recreation Handbook. A Review of Issues and Management Practice; The Wildfowl and Wetlands Trust Publisher: Gloucester, UK, 2004. [Google Scholar]
- Rubega, M.A.; Robinson, J.A. Water salinization and shorebirds: Emerging issues. Int. Wader Stud. 1997, 9, 45–54. [Google Scholar]
- Holm, T.E.; Clausen, P. Effects of water level management on autumn staging waterbird and macrophyte diversity in three Danish coastal lagoons. Biodivers. Conserv. 2006, 15, 4399–4423. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wetland | Wetland size (ha) | Isolation (m) | Distance to the Coastline (m) | Mean Water Level (cm) | Water-Level Fluctuation (cm) | Salinity | Water Diversion | Water Discharges | Tourism Pressure | Anthropization |
|---|---|---|---|---|---|---|---|---|---|---|
| Auruca | 31.4 | 461 | 1009 | 0.0 | 0.0 | 1 | 3 | 2 | 0 | 3 |
| Baronello | 20.5 | 461 | 779 | 24.3 | 10.7 | 0 | 1 | 2 | 1 | 2 |
| Cuba | 61.5 | 208 | 1499 | 2.9 | 3.0 | 0 | 2 | 0 | 0 | 1 |
| Faro | 26.9 | 818 | 409 | 1849.3 | 1.4 | 2 | 3 | 2 | 3 | 3 |
| Ganzirri | 34.1 | 816 | 990 | 497.6 | 2.6 | 2 | 3 | 3 | 3 | 3 |
| Gela | 107.8 | 65,943 | 2326 | 99.5 | 42.2 | 0 | 3 | 3 | 1 | 3 |
| Gornalunga | 13.9 | 63,208 | 0 | 33.9 | 11.9 | 2 | 0 | 2 | 2 | 3 |
| Longarini | 109.9 | 208 | 1674 | 1.2 | 1.5 | 3 | 3 | 3 | 3 | 3 |
| Morghella | 49.6 | 4798 | 19 | 0.6 | 1.2 | 1 | 3 | 2 | 1 | 3 |
| Pantano Bruno | 16.5 | 1419 | 1162 | 11.2 | 6.8 | 0 | 2 | 2 | 1 | 3 |
| Pantano Grande | 28.3 | 0 | 274 | 4.9 | 3.0 | 2 | 0 | 0 | 3 | 0 |
| Pantano Piccolo | 15.9 | 0 | 19 | 26.3 | 5.0 | 1 | 0 | 0 | 2 | 0 |
| Preola | 30.8 | 488 | 1318 | 145.6 | 6.4 | 0 | 0 | 0 | 1 | 2 |
| Roveto | 124.1 | 628 | 0 | 14.1 | 10.2 | 3 | 1 | 2 | 1 | 1 |
| Tindari | 7.4 | 50,349 | 198 | 200.1 | 86.4 | 3 | 0 | 0 | 3 | 0 |
| Tondi | 6.1 | 488 | 1916 | 506.1 | 17.4 | 0 | 0 | 0 | 0 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrarini, A.; Celada, C.; Gustin, M. Anthropogenic Pressure and Climate Change Could Severely Hamper the Avian Metacommunity of the Sicilian Wetlands. Diversity 2022, 14, 696. https://doi.org/10.3390/d14090696
Ferrarini A, Celada C, Gustin M. Anthropogenic Pressure and Climate Change Could Severely Hamper the Avian Metacommunity of the Sicilian Wetlands. Diversity. 2022; 14(9):696. https://doi.org/10.3390/d14090696
Chicago/Turabian StyleFerrarini, Alessandro, Claudio Celada, and Marco Gustin. 2022. "Anthropogenic Pressure and Climate Change Could Severely Hamper the Avian Metacommunity of the Sicilian Wetlands" Diversity 14, no. 9: 696. https://doi.org/10.3390/d14090696