
The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Definition of Variables Used in This Study
2.3. Data Collection
2.4. Data Analysis
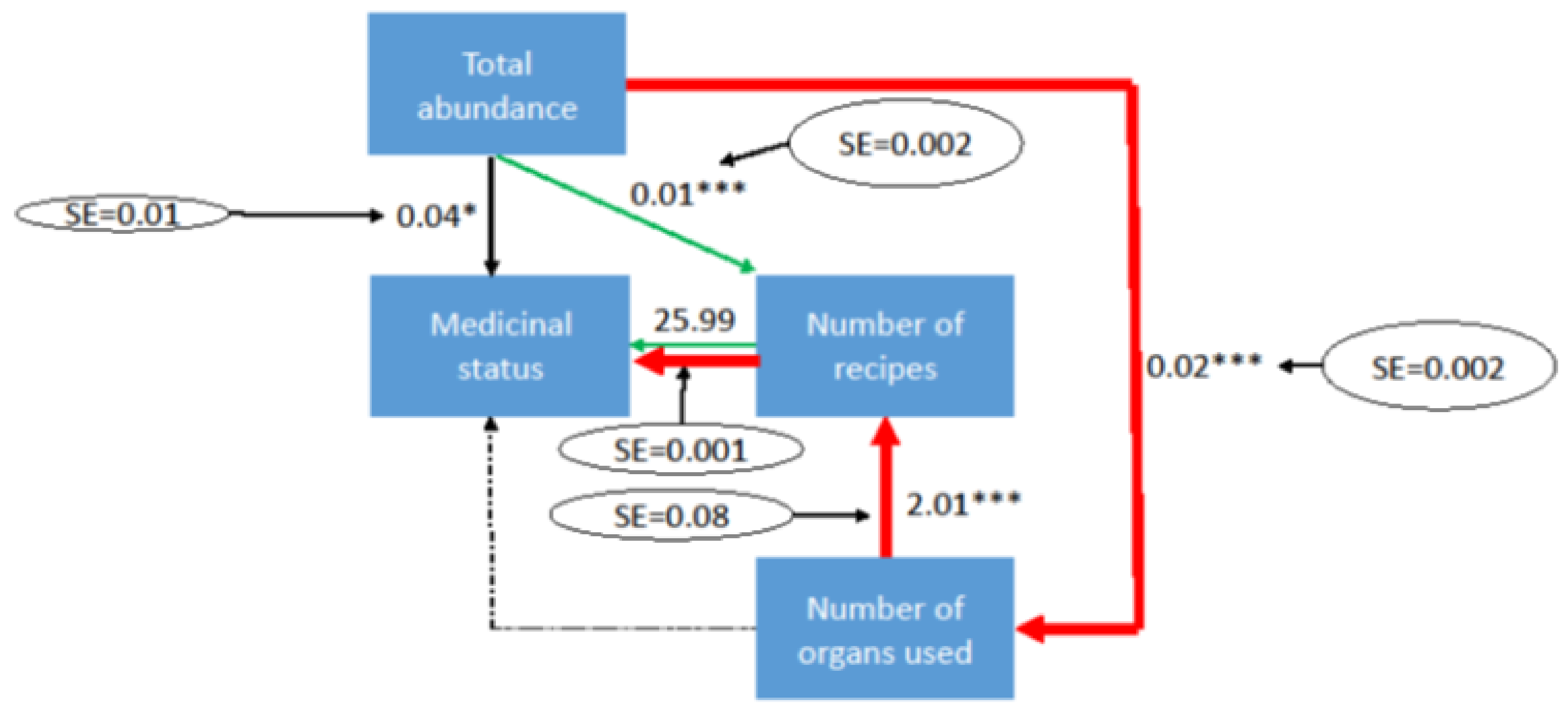
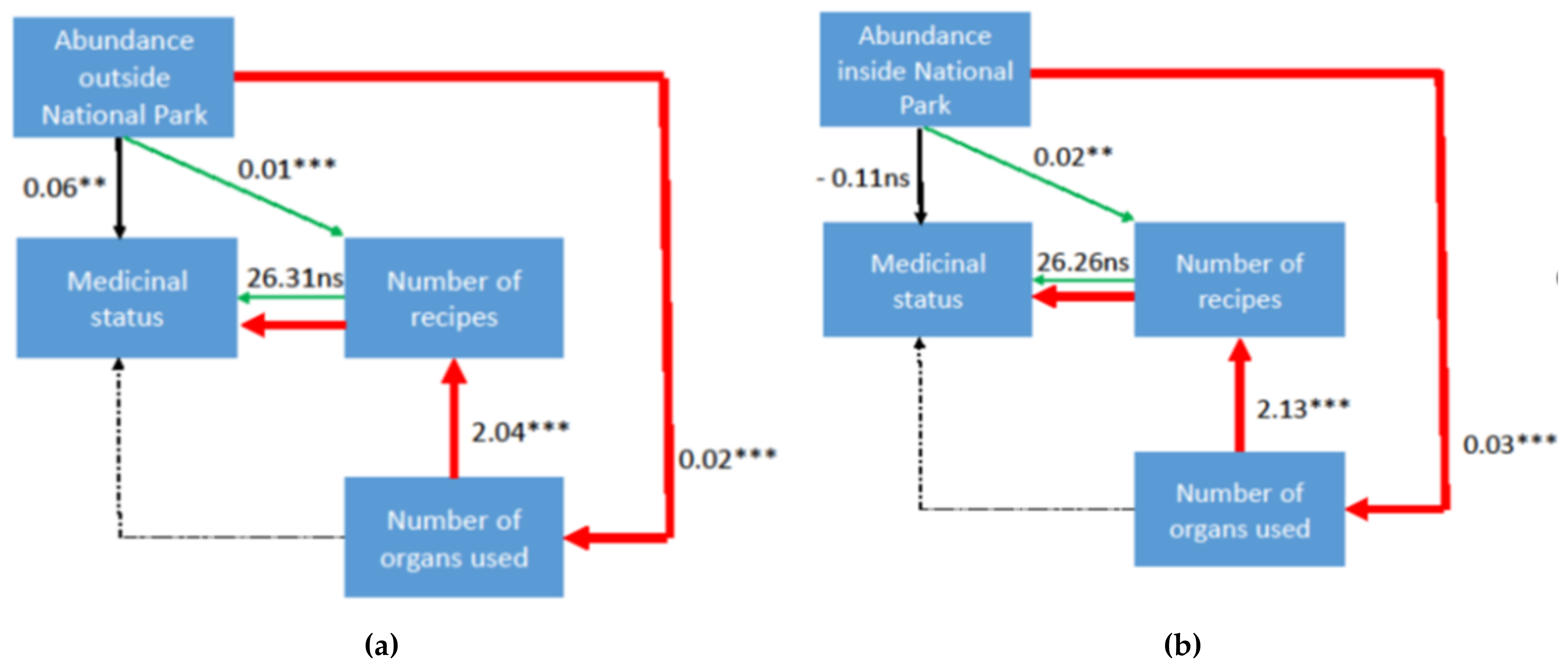
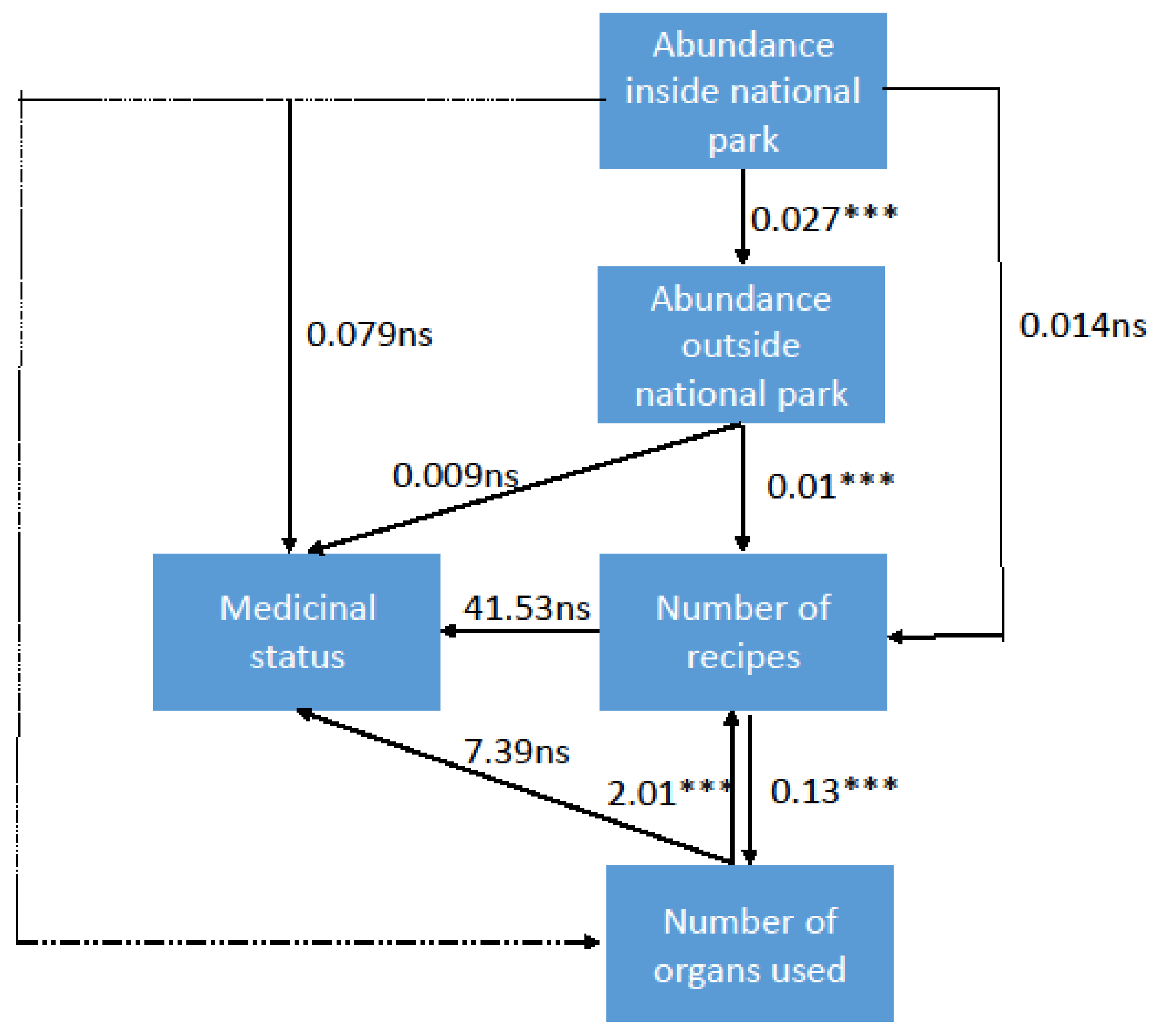
3. Results and Discussion
4. Way Forward: Ex-Situ Conservation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs Over the 30 Years from 1981 to 2012. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schippmann, U.; Leaman, J.D.; Cunningham, A.B. Impact of Cultivation and Gathering of Medicinal Plants on Biodiversity: Global Trends and Issues; Inter-Departmental Working Group on Biological Diversity for Food and Agriculture: Rome, Italy, 2002. [Google Scholar]
- Voeks, R.A. Disturbance pharmacopoeias: Medicine and myth from the humid Tropics. Ann. Assoc. Am. Geogr. 2004, 94, 868–888. [Google Scholar]
- Albuquerque, U.P. Re-examining hypotheses concerning the use and knowledge of medicinal plants: A study in the Caatinga vegetation of NE Brazil. J. Ethnobiol. Ethnomed. 2006, 2, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saslis-Lagoudakis, C.H.; Hawkins, J.A.; Greenhill, S.J.; Pendry, C.A.; Watson, M.F.; Tuladhar-Douglas, W.; Baral, S.R.; Savolainen, V. The evolution of traditional knowledge: Environment shapes medicinal plant use in Nepal. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yessoufou, K.; Ambani, A.E.; Elansary, H.O.; El-Sabrout, A.M.; Shokralla, S. Time, Mediated through Plant Versatility, Is a Better Predictor of Medicinal Status of Alien Plants. Diversity 2022, 14, 286. [Google Scholar] [CrossRef]
- de Oliveira Trindade, M.R.; Jardim, J.G.; Casas, A.; Guerra, N.M.; de Lucena, R.F.P. Availability and use of woody plant resources in two areas of Caatinga in Northeastern Brazil. Ethnobot. Res. Appl. 2015, 14, 313–957. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, P.H.S.; Albuquerque, U.P.; Medeiros, P.M. The most commonly available woody plant species are the most useful for human populations: A meta-analysis. Ecol. Appl. 2016, 26, 2238–2253. [Google Scholar] [CrossRef] [PubMed]
- DeFries, R.; Hansen, A.; Turner, B.L.; Reid, R.; Liu, J.G. Land use change around protected areas: Management to balance human needs and ecological function. Ecol. Appl. 2007, 17, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Dudley, N.; Stolton, S.; Belokurov, A.; Krueger, L.; Lopoukhine, N.; MacKinnon, K.; Sandwith, T.; Sekhran, N. Natural Solutions: Protected Areas Helping People Cope with Climate Change; IUCNWCPA: Gland, Switzerland; TNC: Washington, DC, USA; UNDP: New York, NY, USA; WCS: New York, NY, USA; The World Bank: Washington, DC, USA; WWF: Gland, Switzerland, 2010. [Google Scholar]
- Heywood, V.H. Conserving plants within and beyond protected areas–still problematic and future uncertain. Plant Divers. 2018, 41, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gülez, S. A method of evaluating areas for national park status. Environ. Manag. 1992, 16, 811–818. [Google Scholar] [CrossRef]
- Schmidt, E.; Lotter, M.; McCleland, W. Trees and Shrubs of Mpumalanga and Kruger National Park; Jacana Press: Johannesburg, South Africa, 2007; 702p. [Google Scholar]
- Lahaye, R.; van der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yessoufou, K.; Davies, T.J.; Maurin, O.; Kuzmina, M.; Schaefer, H.; van der Bank, M.; Savolainen, V. Large herbivores favour species diversity but have mixed impacts on phylogenetic community structure in an African savanna ecosystem. J. Ecol. 2013, 101, 614–625. [Google Scholar] [CrossRef]
- Perera, S.J.; Ratnayake-Perera, D.; Procheş, Ş. Vertebrate distributions indicate a Greater Maputaland-Pondoland-Albany region of endemism. S. Afr. J. Sci. 2011, 107, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Coates-Palgrave, M. Keith Coates-Palgrave. In Trees of Southern Africa, 3rd ed.; Struik Nature: Cape Town, South Africa, 2002. [Google Scholar]
- van Wyk, B.; van Wyk, P. Field Guide to Trees of Southern Africa, 2nd ed.; Struik Publisher: Cape Town, South Africa, 2013. [Google Scholar]
- Glen, H.; Van Wyk, B. Guide to Trees Introduced into Southern Africa; Penguin Random House: Cape Town, South Africa, 2016. [Google Scholar]
- Royal Museum for Central Africa. Prelude Database for Medicinal Plants in Africa. 2018. Available online: http://www.africamuseum.be/en/research/collections_libraries/biology/prelude (accessed on 25 January 2018).
- Germishuizen, G.; Meyer, N.L. Plants of Southern Africa: An Annotated Checklist; National Botanical Institute: Pretoria, South Africa, 2003. [Google Scholar]
- Boon, D. Pooley’s Trees of Eastern South Africa–A Complete Guide; Flora and Fauna Publication Trust: Durban, South Africa, 2010. [Google Scholar]
- Van Wyk, B.; van der Berg, E.; Coates-Palgrave, M.; Jordaan, M. Dictionary of Names for Southern African Trees, 1st ed.; Briza: Pretoria, South Africa, 2011. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Lefcheck, J.S. PIECEWISESEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Venter, F.J. A Classification of Land for Management Planning in the Kruger National Park. PhD Thesis, University of South Africa, Pretoria, South Africa, 2012. [Google Scholar]
- Ford, R.I. The Nature and Status of Ethnobotany; University of Michigan: Ann Arbor, MI, USA, 1978. [Google Scholar]
- Phillips, O.; Gentry, A.H. The useful plants of Tambopata, Peru: I. Statistical hypotheses tests with a new quantitative technique. Econ. Bot. 1993, 47, 15–32. [Google Scholar] [CrossRef]
- Bennett, B.C. Ethnobotany education, opportunities, and needs in the US. Ethnobot. Res. Appl. 2005, 3, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.J. Ethnobotany: A Methods Manual; Earthscan: London, UK, 2007. [Google Scholar]
- Albuquerque, U.P.; Hanazaki, N. Commentary: Five Problems in Current Ethnobotanical Research—And Some Suggestions for Strengthening Them. Human Ecol. 2009, 37, 653–661. [Google Scholar] [CrossRef]
- Gaoue, O.G.; Coe, M.A.; Bond, M.; Hart, G.; Seyler, B.C.; McMillen, H. Theories and Major Hypotheses in Ethnobotany. Econ. Bot. 2017, 71, 269–287. [Google Scholar] [CrossRef]
- Robles Ariasa, D.M.; Cevallos, D.; Gaoue, O.; Fadimana, M.G.; Hindlea, T. Non-random medicinal plants selection in the Kichwa community of the Ecuadorian Amazon. J. Ethnopharmacol. 2020, 246, 112220. [Google Scholar] [CrossRef] [PubMed]
- CBD. Global Biodiversity Outlook 2; Secretariat of the Convention on Biological Diversity: Montreal, QC, Canada, 2006; p. 81. [Google Scholar]
- IUCN. IUCN Technical Guidelines on the Management of Ex Situ Populations for Conservation. In Proceedings of the 14th Meeting of the Programme Committee of Council, Gland, Switzerland, 10 December 2002. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yessoufou, K.; Muleba, I.; Rampedi, I.T. The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species. Diversity 2022, 14, 693. https://doi.org/10.3390/d14080693
Yessoufou K, Muleba I, Rampedi IT. The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species. Diversity. 2022; 14(8):693. https://doi.org/10.3390/d14080693
Chicago/Turabian StyleYessoufou, Kowiyou, Isidore Muleba, and Isaac T. Rampedi. 2022. "The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species" Diversity 14, no. 8: 693. https://doi.org/10.3390/d14080693